
Comparison and Characterization of Oxidation Resistance and Carbohydrate Content in Cd-Tolerant and -Sensitive Kentucky Bluegrass under Cd Stress

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seedling Cultivation and Cd Treatment
2.2. Plant Height, Root Length, Fresh Weight, and Cd Determination
2.3. Determination of MDA, H2O2, and O2− Content
2.3.1. MDA Determination
2.3.2. Determination of H2O2 Content
2.3.3. Determination of the Rate of O2− Generation
2.4. Determination of POD, SOD, APX, CAT Activities, and Soluble Protein Content
2.5. Determination of Reducing Sugar, Soluble Sugar, Sucrose, Trehalose, Fructose, and Glucose Content
2.5.1. Determination of Reducing Sugar and Soluble Sugar Content
2.5.2. Determination of Sucrose, Trehalose, Fructose, and Glucose Content
2.6. Determination of Pectin, Lignin, and Cellulose Content
2.7. Statistical Analysis
3. Results
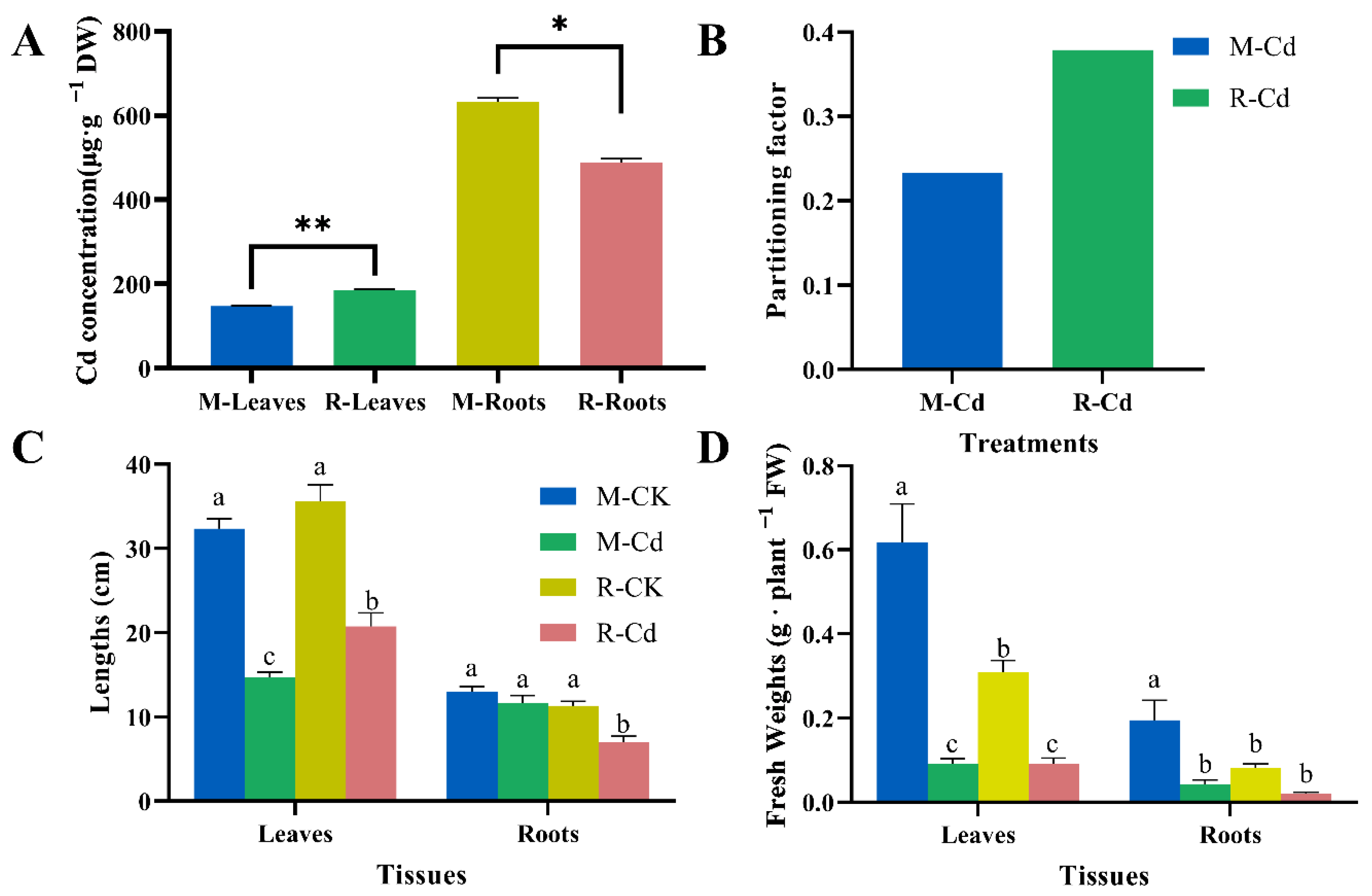
3.1. Cd Accumulation and Distribution and the Effect of Cd Stress on the Growth Status of Kentucky Bluegrass
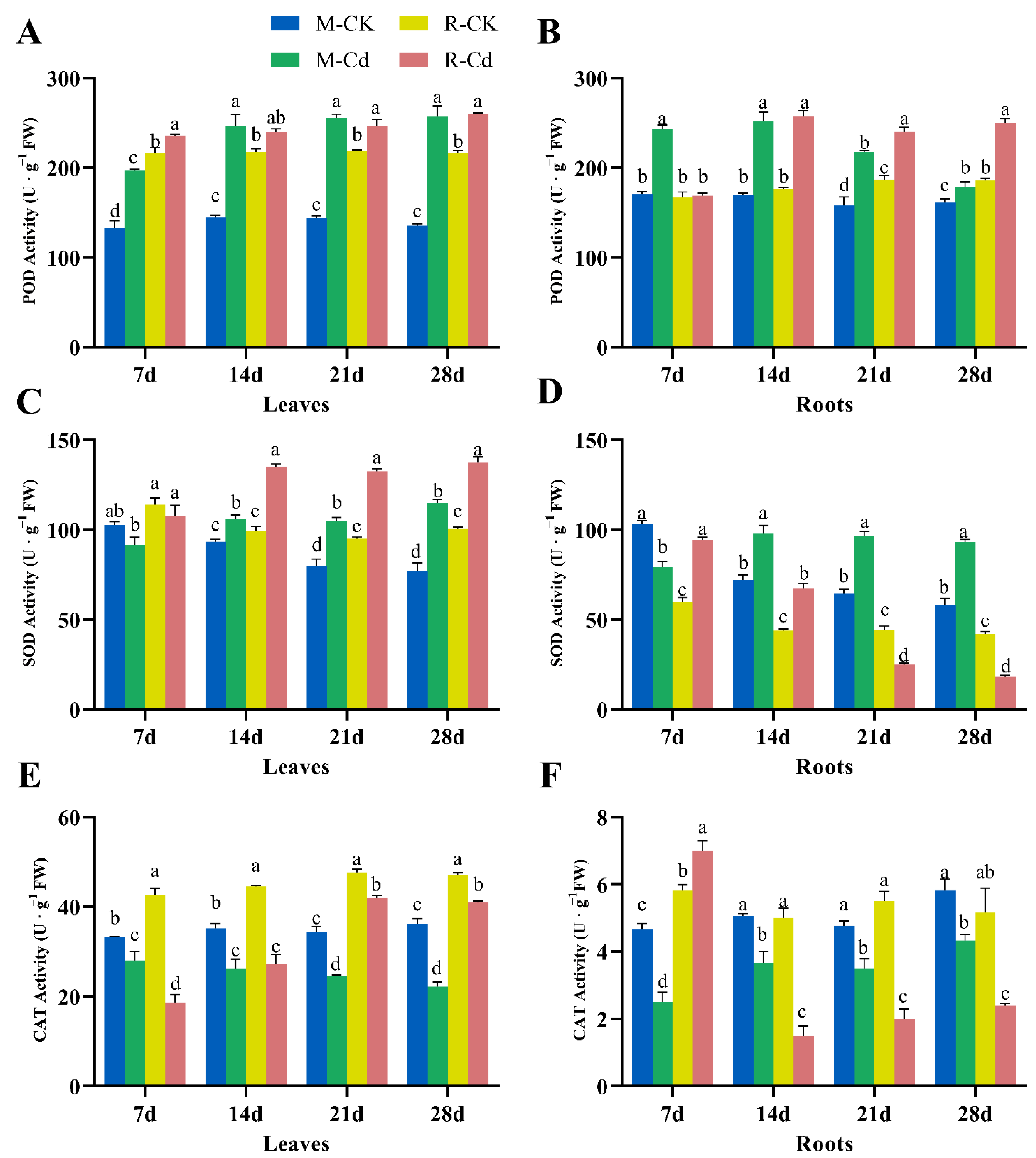
3.2. Effects of Cd on Antioxidant Enzyme Activity
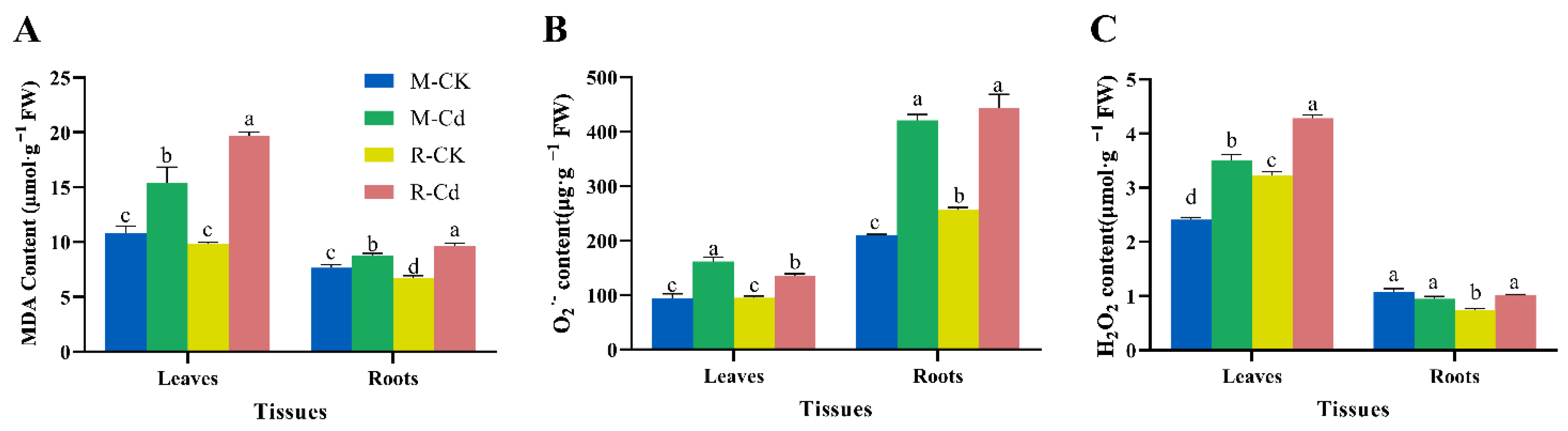
3.3. Effects of Cd on MDA, H2O2, and O2− Content
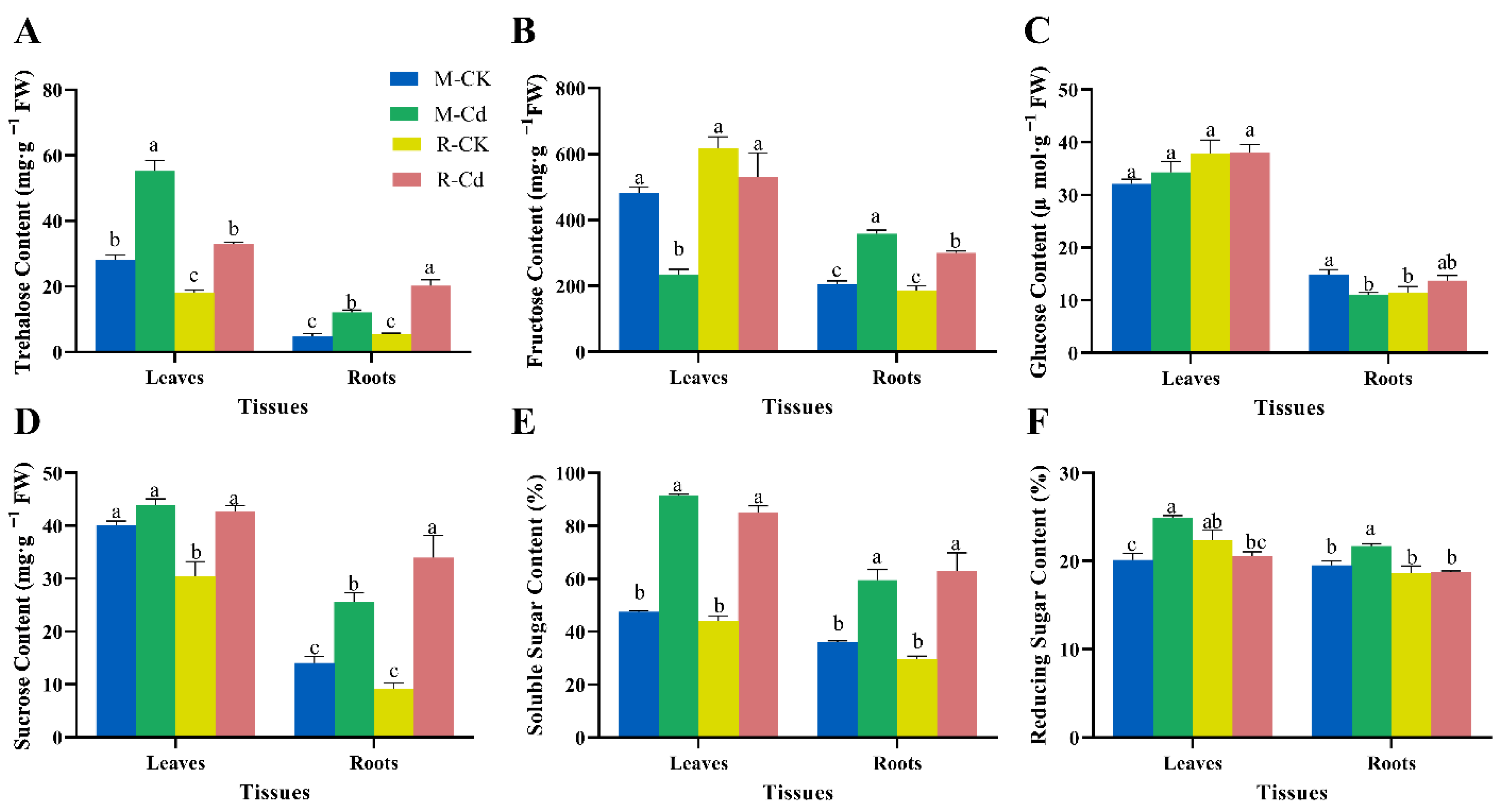
3.4. Effects of Cd on Non-Structural Carbohydrate Contents
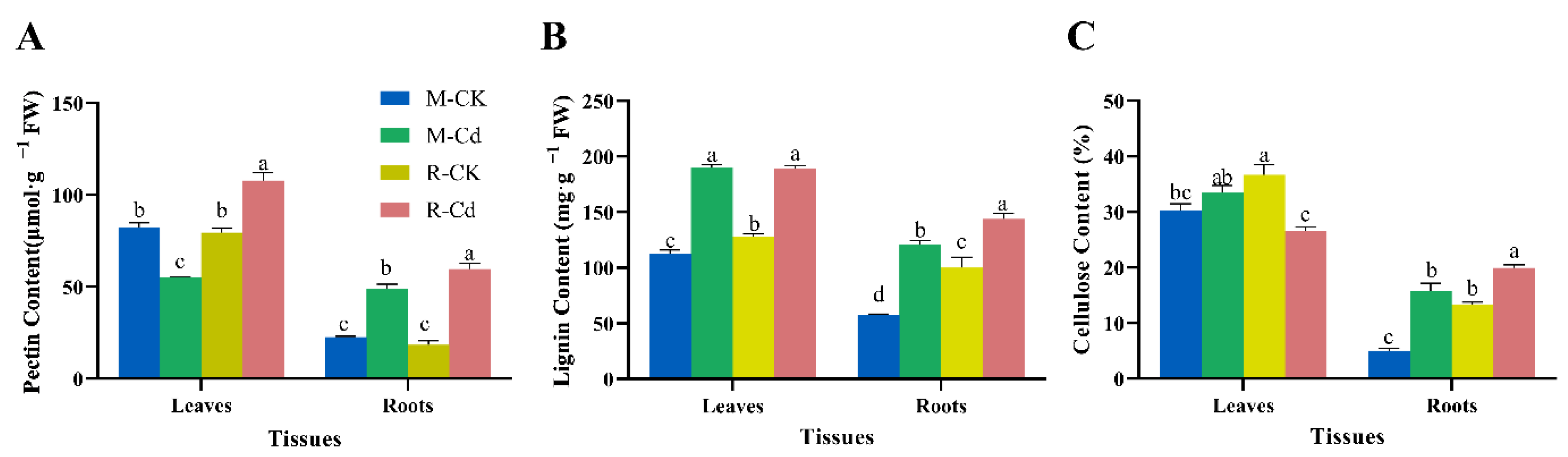
3.5. Effects of Cd on Structural Carbohydrate Contents
4. Discussion
4.1. Cd-Induced Growth Inhibition of the Leaves and Roots and the Defensive Reaction of the Cell Wall
4.2. Cd-Induced Antioxidative Stress Reaction of Kentucky Bluegrass
4.3. Non-Structural Carbohydrates Involved in the Defense against Cd Stress in Kentucky Bluegrass
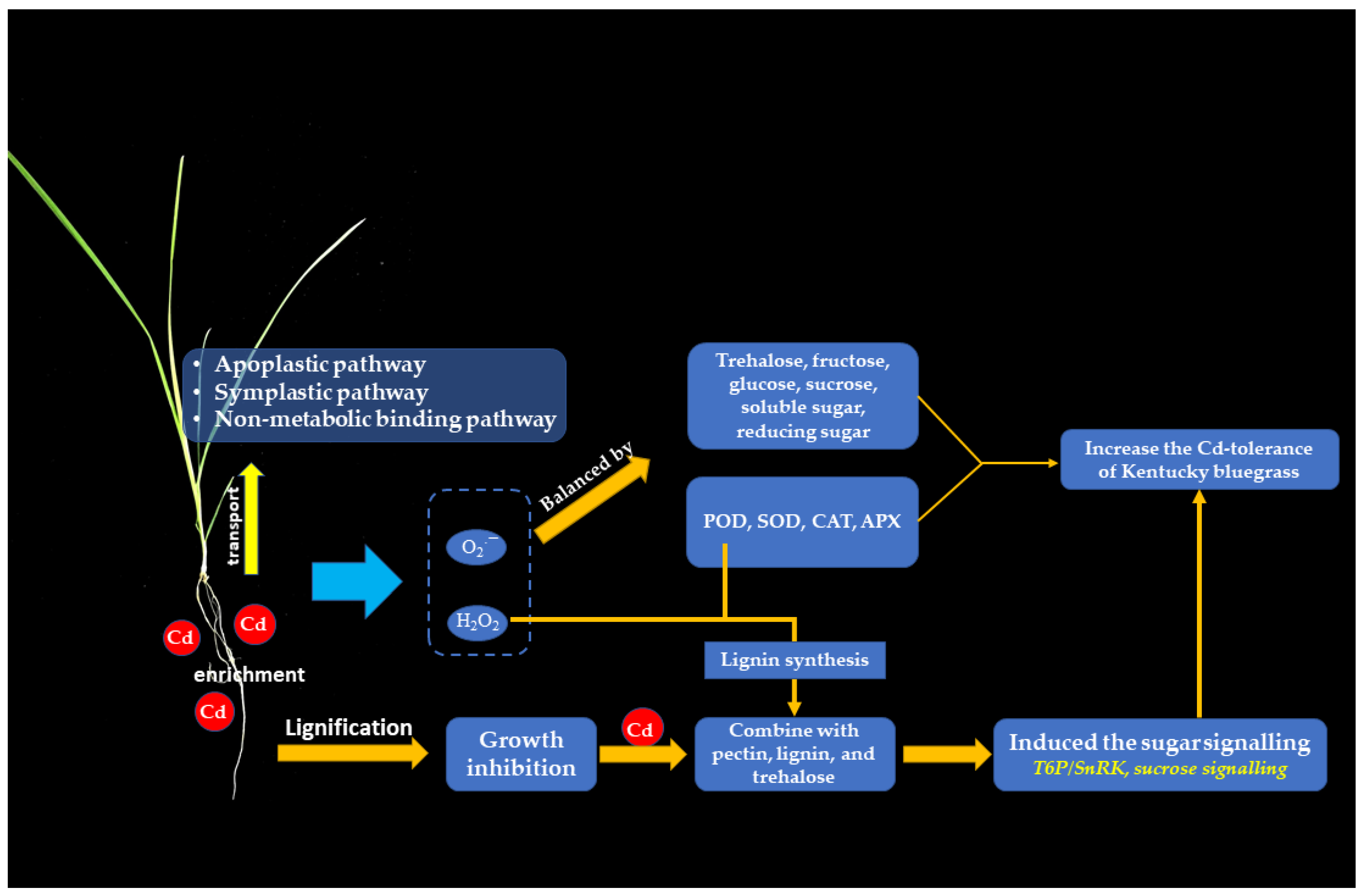
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling Cadmium Toxicity and Tolerance in Plants: Insight into Regulatory Mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Li, Z.; Ma, Z.; van der Kuijp, T.J.; Yuan, Z.; Huang, L. A Review of Soil Heavy Metal Pollution from Mines in China: Pollution and Health Risk Assessment. Sci. Total Environ. 2014, 468–469, 843–853. [Google Scholar] [CrossRef]
- Mielke, H.W.; Adams, J.L.; Chaney, R.L.; Mielke, P.W.; Ravikumar, V.C. The Pattern of Cadmium in the Environment of Five Minnesota Cities. Environ Geochem Health 1991, 13, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Subrahmanyam, G.; Mondal, R.; Cabral-Pinto, M.M.S.; Shabnam, A.A.; Jigyasu, D.K.; Malyan, S.K.; Fagodiya, R.K.; Khan, S.A.; Kumar, A.; et al. Bio-Remediation Approaches for Alleviation of Cadmium Contamination in Natural Resources. Chemosphere 2021, 268, 128855. [Google Scholar] [CrossRef]
- Prozialeck, W.C.; Vaidya, V.S.; Liu, J.; Waalkes, M.P.; Edwards, J.R.; Lamar, P.C.; Bernard, A.M.; Dumont, X.; Bonventre, J.V. Kidney Injury Molecule-1 Is an Early Biomarker of Cadmium Nephrotoxicity. Kidney Int. 2007, 72, 985–993. [Google Scholar] [CrossRef] [Green Version]
- Pulscher, L.A.; Gray, R.; McQuilty, R.; Rose, K.; Welbergen, J.A.; Phalen, D.N. Evidence of Chronic Cadmium Exposure Identified in the Critically Endangered Christmas Island Flying-Fox (Pteropus natalis). Sci. Total Environ. 2021, 766, 144374. [Google Scholar] [CrossRef] [PubMed]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium Toxicity in Plants: Impacts and Remediation Strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef]
- Chakrabarti, M.; Mukherjee, A. Investigating the Underlying Mechanism of Cadmium-Induced Plant Adaptive Response to Genotoxic Stress. Ecotoxicol. Environ. Saf. 2021, 209, 111817. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. REACTIVE OXYGEN SPECIES: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Pommerrenig, B.; Ludewig, F.; Cvetkovic, J.; Trentmann, O.; Klemens, P.A.W.; Neuhaus, H.E. In Concert: Orchestrated Changes in Carbohydrate Homeostasis Are Critical for Plant Abiotic Stress Tolerance. Plant Cell Physiol. 2018, 59, 1290–1299. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl Radical Scavenging Activity of Compatible Solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of Photosynthesis during Abiotic Stress-Induced Photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljung, K.; Nemhauser, J.L.; Perata, P. New Mechanistic Links between Sugar and Hormone Signalling Networks. Curr. Opin. Plant Biol. 2015, 25, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Bolouri-Moghaddam, M.R.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar Signalling and Antioxidant Network Connections in Plant Cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Griffiths, C.A.; Sagar, R.; Geng, Y.; Primavesi, L.F.; Patel, M.K.; Passarelli, M.K.; Gilmore, I.S.; Steven, R.T.; Bunch, J.; Paul, M.J.; et al. Chemical Intervention in Plant Sugar Signalling Increases Yield and Resilience. Nature 2016, 540, 574–578. [Google Scholar] [CrossRef]
- Dong, S.; Beckles, D.M. Dynamic Changes in the Starch-Sugar Interconversion within Plant Source and Sink Tissues Promote a Better Abiotic Stress Response. J. Plant Physiol. 2019, 234, 80–93. [Google Scholar] [CrossRef]
- Loix, C.; Huybrechts, M.; Vangronsveld, J.; Gielen, M.; Keunen, E.; Cuypers, A. Reciprocal Interactions between Cadmium-Induced Cell Wall Responses and Oxidative Stress in Plants. Front. Plant Sci. 2017, 8, 1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Wang, Z. Physiological Mechanism of Hypertolerance of Cadmium in Kentucky Bluegrass and Tall Fescue: Chemical Forms and Tissue Distribution. Environ. Exp. Bot. 2013, 96, 35–42. [Google Scholar] [CrossRef]
- Podazza, G.; Arias, M.; Prado, F.E. Cadmium Accumulation and Strategies to Avoid Its Toxicity in Roots of the Citrus Rootstock Citrumelo. J. Hazard. Mater. 2012, 215–216, 83–89. [Google Scholar] [CrossRef]
- Finger-Teixeira, A.; de Lourdes Lucio Ferrarese, M.; Ricardo Soares, A.; da Silva, D.; Ferrarese-Filho, O. Cadmium-Induced Lignification Restricts Soybean Root Growth. Ecotoxicol. Environ. Saf. 2010, 73, 1959–1964. [Google Scholar] [CrossRef]
- Moura, J.C.M.S.; Bonine, C.A.V.; Viana, J.D.O.F.; Dornelas, M.C.; Mazzafera, P. Abiotic and Biotic Stresses and Changes in the Lignin Content and Composition in Plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef]
- Guo, X.; Luo, J.; Du, Y.; Li, J.; Liu, Y.; Liang, Y.; Li, T. Coordination between Root Cell Wall Thickening and Pectin Modification Is Involved in Cadmium Accumulation in Sedum Alfredii. Environ. Pollut. 2021, 268, 115665. [Google Scholar] [CrossRef]
- Li, H.; Zheng, X.; Tao, L.; Yang, Y.; Gao, L.; Xiong, J. Aeration Increases Cadmium (Cd) Retention by Enhancing Iron Plaque Formation and Regulating Pectin Synthesis in the Roots of Rice (Oryza sativa) Seedlings. Rice 2019, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Muschitz, A.; Riou, C.; Mollet, J.-C.; Gloaguen, V.; Faugeron, C. Modifications of Cell Wall Pectin in Tomato Cell Suspension in Response to Cadmium and Zinc. Acta Physiol. Plant. 2015, 37, 245. [Google Scholar] [CrossRef]
- Cheng, S.; Yu, H.; Hu, M.; Wu, Y.; Cheng, L.; Cai, Q.; Tu, Y.; Xia, T.; Peng, L. Miscanthus Accessions Distinctively Accumulate Cadmium for Largely Enhanced Biomass Enzymatic Saccharification by Increasing Hemicellulose and Pectin and Reducing Cellulose CrI and DP. Bioresour. Technol. 2018, 263, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, Z. A Comparison Study in Cadmium Tolerance and Accumulation in Two Cool-Season Turfgrasses and Solanum nigrum L. Water Air Soil Pollut. 2014, 225, 1938. [Google Scholar] [CrossRef]
- Xian, J.; Wang, Y.; Niu, K.; Ma, H.; Ma, X. Transcriptional Regulation and Expression Network Responding to Cadmium Stress in a Cd-Tolerant Perennial Grass Poa Pratensis. Chemosphere 2020, 250, 126158. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Ma, X.; Liang, G.; Ma, H.; Jia, Z.; Liu, W.; Yu, Q. 5-Aminolevulinic Acid Modulates Antioxidant Defense Systems and Mitigates Drought-Induced Damage in Kentucky Bluegrass Seedlings. Protoplasma 2017, 254, 2083–2094. [Google Scholar] [CrossRef] [PubMed]
- Kelestemur, S.; Özcan, M. Determination of Pb in Glasses by Direct Solid Sampling and High-Resolution Continuum Source Graphite Furnace Atomic Absorption Spectrometry: Method Development and Analyses of Glass Samples. Microchem. J. 2015, 118, 55–61. [Google Scholar] [CrossRef]
- Kumar, G.; Knowles, N.R. Changes in Lipid Peroxidation and Lipolytic and Free-Radical Scavenging Enzyme Activities during Aging and Sprouting of Potato (Solanum tuberosum) Seed-Tubers. Plant Physiol. 1993, 102, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Quintanilla-Guerrero, F.; Duarte-Vázquez, M.A.; García-Almendarez, B.E.; Tinoco, R.; Vazquez-Duhalt, R.; Regalado, C. Polyethylene Glycol Improves Phenol Removal by Immobilized Turnip Peroxidase. Bioresour. Technol. 2008, 99, 8605–8611. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, D.; Lin, X. Effects of Waterlogging on Nitrogen Accumulation and Alleviation of Waterlogging Damage by Application of Nitrogen Fertilizer and Mixtalol in Winter Rape (Brassica napus L.). J. Plant Growth Regul. 1997, 16, 47–53. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and Developmental Characterization of Multiple Forms of Catalase in Tobacco Leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Pena, L.B.; Pasquini, L.A.; Tomaro, M.L.; Gallego, S.M. Proteolytic System in Sunflower (Helianthus annuus L.) Leaves under Cadmium Stress. Plant Sci. 2006, 171, 531–537. [Google Scholar] [CrossRef]
- Miller, G.L. Lorenz. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Sinay, H.; Karuwal, R.L. Proline and Total Soluble Sugar Content at the Vegetative Phase of Six Corn Cultivars from Kisar Island Maluku, Grown under Drought Stress Conditions. Int. J. Adv. Agric. Res. 2014, 2, 77–82. [Google Scholar]
- Cadet, F.; Offmann, B. Direct Spectroscopic Sucrose Determination of Raw Sugar Cane Juices. J. Agric. Food Chem. 1997, 45, 166–171. [Google Scholar] [CrossRef]
- Viles, F.J.; Silverman, L. Determination of Starch and Cellulose with Anthrone. Anal. Chem. 1949, 21, 950–953. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Terrón-Camero, L.C.; Peláez-Vico, M.Á.; Olmedilla, A.; Sandalio, L.M. Reactive Oxygen and Nitrogen Species as Key Indicators of Plant Responses to Cd Stress. Environ. Exp. Bot. 2019, 161, 107–119. [Google Scholar] [CrossRef]
- Hédiji, H.; Djebali, W.; Cabasson, C.; Maucourt, M.; Baldet, P.; Bertrand, A.; Boulila Zoghlami, L.; Deborde, C.; Moing, A.; Brouquisse, R.; et al. Effects of Long-Term Cadmium Exposure on Growth and Metabolomic Profile of Tomato Plants. Ecotoxicol. Environ. Saf. 2010, 73, 1965–1974. [Google Scholar] [CrossRef] [PubMed]
- Arruda, M.a.Z.; Azevedo, R.A. Metallomics and Chemical Speciation: Towards a Better Understanding of Metal-Induced Stress in Plants. Ann. Appl. Biol. 2009, 155, 301–307. [Google Scholar] [CrossRef]
- Cutler, J.M.; Rains, D.W. Characterization of Cadmium Uptake by Plant Tissue. Plant Physiol. 1974, 54, 67. [Google Scholar] [CrossRef] [Green Version]
- Dong, Q.; Xu, P.; Wang, Z. Differential Cadmium Distribution and Translocation in Roots and Shoots Related to Hyper-Tolerance between Tall Fescue and Kentucky Bluegrass. Front. Plant Sci. 2017, 8, 113. [Google Scholar] [CrossRef] [Green Version]
- Gutsch, A.; Sergeant, K.; Keunen, E.; Prinsen, E.; Guerriero, G.; Renaut, J.; Hausman, J.-F.; Cuypers, A. Does Long-Term Cadmium Exposure Influence the Composition of Pectic Polysaccharides in the Cell Wall of Medicago Sativa Stems? BMC Plant Biol. 2019, 19, 271. [Google Scholar] [CrossRef] [Green Version]
- Krzeslowska, M. The Cell Wall in Plant Cell Response to Trace Metals: Polysaccharide Remodeling and Its Role in Defense Strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, R.; Yan, X.; Liang, X.; Sun, Y.; Xu, Y. Pivotal Role for Root Cell Wall Polysaccharides in Cultivar-Dependent Cadmium Accumulation in Brassica chinensis L. Ecotoxicol. Environ. Saf. 2020, 194, 110369. [Google Scholar] [CrossRef] [PubMed]
- van de Mortel, J.E.; Almar Villanueva, L.; Schat, H.; Kwekkeboom, J.; Coughlan, S.; Moerland, P.D.; Ver Loren van Themaat, E.; Koornneef, M.; Aarts, M.G.M. Large Expression Differences in Genes for Iron and Zinc Homeostasis, Stress Response, and Lignin Biosynthesis Distinguish Roots of Arabidopsis thaliana and the Related Metal Hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2006, 142, 1127–1147. [Google Scholar] [CrossRef] [Green Version]
- Song, X.-Q.; Liu, L.-F.; Jiang, Y.-J.; Zhang, B.-C.; Gao, Y.-P.; Liu, X.-L.; Lin, Q.-S.; Ling, H.-Q.; Zhou, Y.-H. Disruption of Secondary Wall Cellulose Biosynthesis Alters Cadmium Translocation and Tolerance in Rice Plants. Mol. Plant 2013, 6, 768–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vatehová, Z.; Malovíková, A.; Kollárová, K.; Kučerová, D.; Lišková, D. Impact of Cadmium Stress on Two Maize Hybrids. Plant Physiol. Biochem. 2016, 108, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.-S.; Wang, Y.-J.; Ding, G.; Ma, H.-L.; Zhang, Y.-J.; Gong, J.-M. A Pivotal Role of Cell Wall in Cadmium Accumulation in the Crassulaceae Hyperaccumulator Sedum Plumbizincicola. Mol. Plant 2017, 10, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.; Sigaud-kutner, T.C.S.; Leitão, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy Metal–Induced Oxidative Stress in Algae1. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Huihui, Z.; Xin, L.; Zisong, X.; Yue, W.; Zhiyuan, T.; Meijun, A.; Yuehui, Z.; Wenxu, Z.; Nan, X.; Guangyu, S. Toxic Effects of Heavy Metals Pb and Cd on Mulberry (Morus alba L.) Seedling Leaves: Photosynthetic Function and Reactive Oxygen Species (ROS) Metabolism Responses. Ecotoxicol. Environ. Saf. 2020, 195, 110469. [Google Scholar] [CrossRef] [PubMed]
- Sandalio, L.M.; Rodríguez-Serrano, M.; Gupta, D.K.; Archilla, A.; Romero-Puertas, M.C.; del Río, L.A. Reactive Oxygen Species and Nitric Oxide in Plants Under Cadmium Stress: From Toxicity to Signaling. In Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 199–215. ISBN 978-1-4614-0815-4. [Google Scholar]
- Keunen, E.; Remans, T.; Bohler, S.; Vangronsveld, J.; Cuypers, A. Metal-Induced Oxidative Stress and Plant Mitochondria. Int. J. Mol. Sci. 2011, 12, 6894–6918. [Google Scholar] [CrossRef] [Green Version]
- Cosio, C.; Dunand, C. Specific Functions of Individual Class III Peroxidase Genes. J. Exp. Bot. 2009, 60, 391–408. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Li, H.; Zhang, X.; Fu, J. Antioxidant Responses and Gene Expression in Perennial Ryegrass (Lolium perenne L.) under Cadmium Stress. Ecotoxicology 2011, 20, 770–778. [Google Scholar] [CrossRef]
- Kosar, F.; Akram, N.A.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A Key Organic Osmolyte Effectively Involved in Plant Abiotic Stress Tolerance. J. Plant Growth Regul. 2019, 38, 606–618. [Google Scholar] [CrossRef]
- Duman, F.; Aksoy, A.; Aydin, Z.; Temizgul, R. Effects of Exogenous Glycinebetaine and Trehalose on Cadmium Accumulation and Biological Responses of an Aquatic Plant (Lemna gibba L.). Water Air Soil Pollut. 2011, 217, 545–556. [Google Scholar] [CrossRef]
- Wingler, A. Transitioning to the Next Phase: The Role of Sugar Signaling throughout the Plant Life Cycle. Plant Physiol. 2018, 176, 1075–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Li, F.; Gao, M.; Huang, Y.; Song, Z. Mechanisms of Trehalose-Mediated Mitigation of Cd Toxicity in Rice Seedlings. J. Clean. Prod. 2020, 267, 121982. [Google Scholar] [CrossRef]
- Meng, H.; Hua, S.; Shamsi, I.H.; Jilani, G.; Li, Y.; Jiang, L. Cadmium-Induced Stress on the Seed Germination and Seedling Growth of Brassica napus L., and Its Alleviation through Exogenous Plant Growth Regulators. Plant Growth Regul. 2009, 58, 47–59. [Google Scholar] [CrossRef]
- Kınay, A. Effects of Cadmium on Nicotine, Reducing Sugars and Phenolic Contents of Basma Tobacco Variety. Fresenius Environ. Bull. 2018, 27, 9195–9202. [Google Scholar]
- Rahoui, S.; Chaoui, A.; El Ferjani, E. Membrane Damage and Solute Leakage from Germinating Pea Seed under Cadmium Stress. J. Hazard. Mater. 2010, 178, 1128–1131. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.S.; Giardina, T. Sucrose and Invertases, a Part of the Plant Defense Response to the Biotic Stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Dubey, R.S. Effect of Cadmium on Soluble Sugars and Enzymes of Their Metabolism in Rice. Biol. Plant. 2001, 44, 117–123. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Cui, T.; Niu, K.; Ma, H. Comparison and Characterization of Oxidation Resistance and Carbohydrate Content in Cd-Tolerant and -Sensitive Kentucky Bluegrass under Cd Stress. Agronomy 2021, 11, 2358. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112358
Wang Y, Cui T, Niu K, Ma H. Comparison and Characterization of Oxidation Resistance and Carbohydrate Content in Cd-Tolerant and -Sensitive Kentucky Bluegrass under Cd Stress. Agronomy. 2021; 11(11):2358. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112358
Chicago/Turabian StyleWang, Yong, Ting Cui, Kuiju Niu, and Huiling Ma. 2021. "Comparison and Characterization of Oxidation Resistance and Carbohydrate Content in Cd-Tolerant and -Sensitive Kentucky Bluegrass under Cd Stress" Agronomy 11, no. 11: 2358. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11112358