
Epiphytic Microbial Community and Post-Harvest Characteristics of Strawberry Fruits as Affected by Plant Nutritional Regime with Silicon

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Shelf-Life Assessment
2.3. Microbiological Analyses
2.4. Statistical Analyses
3. Results
3.1. Plants Growth and Yield Parameters
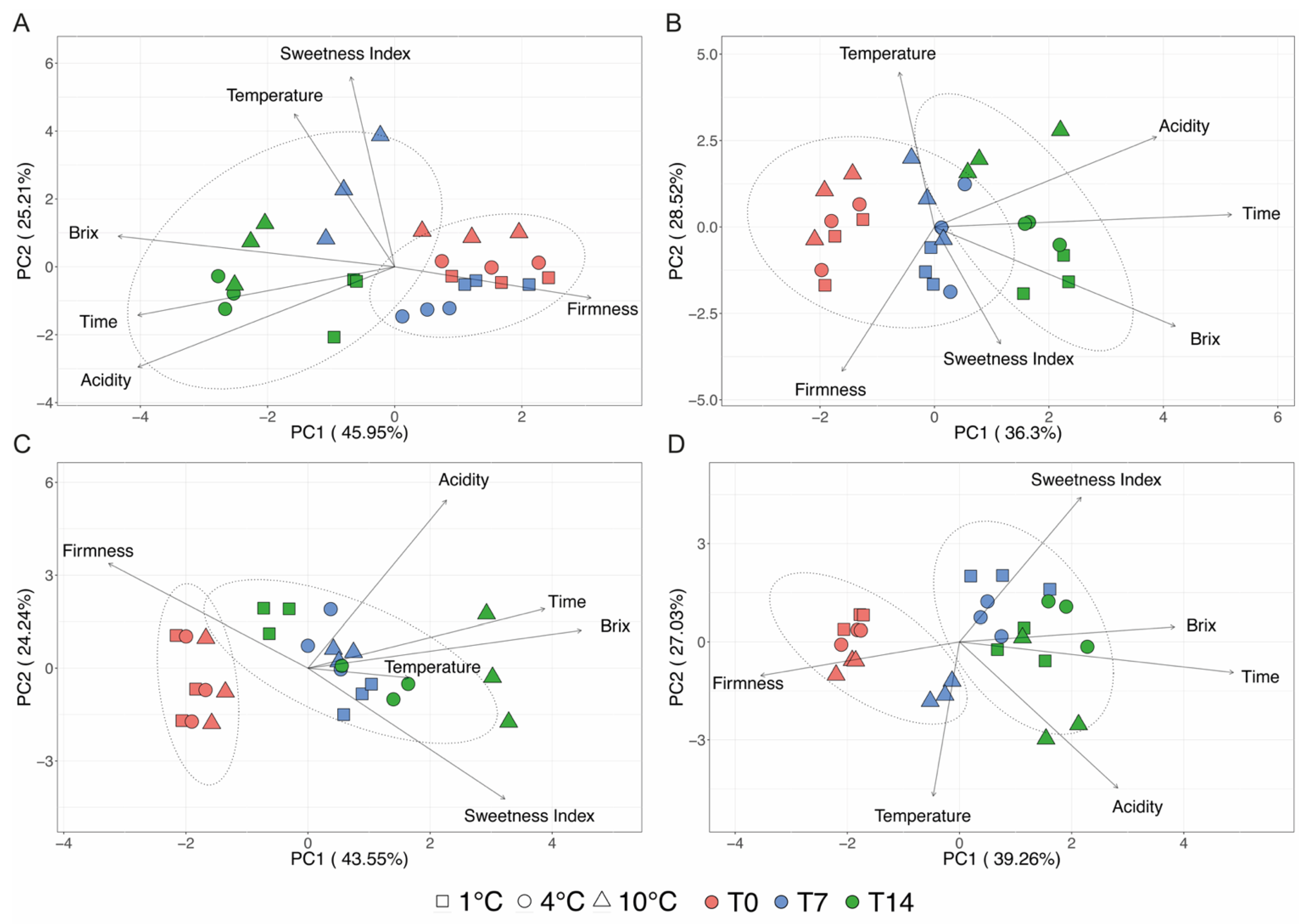
3.2. Post-Harvest
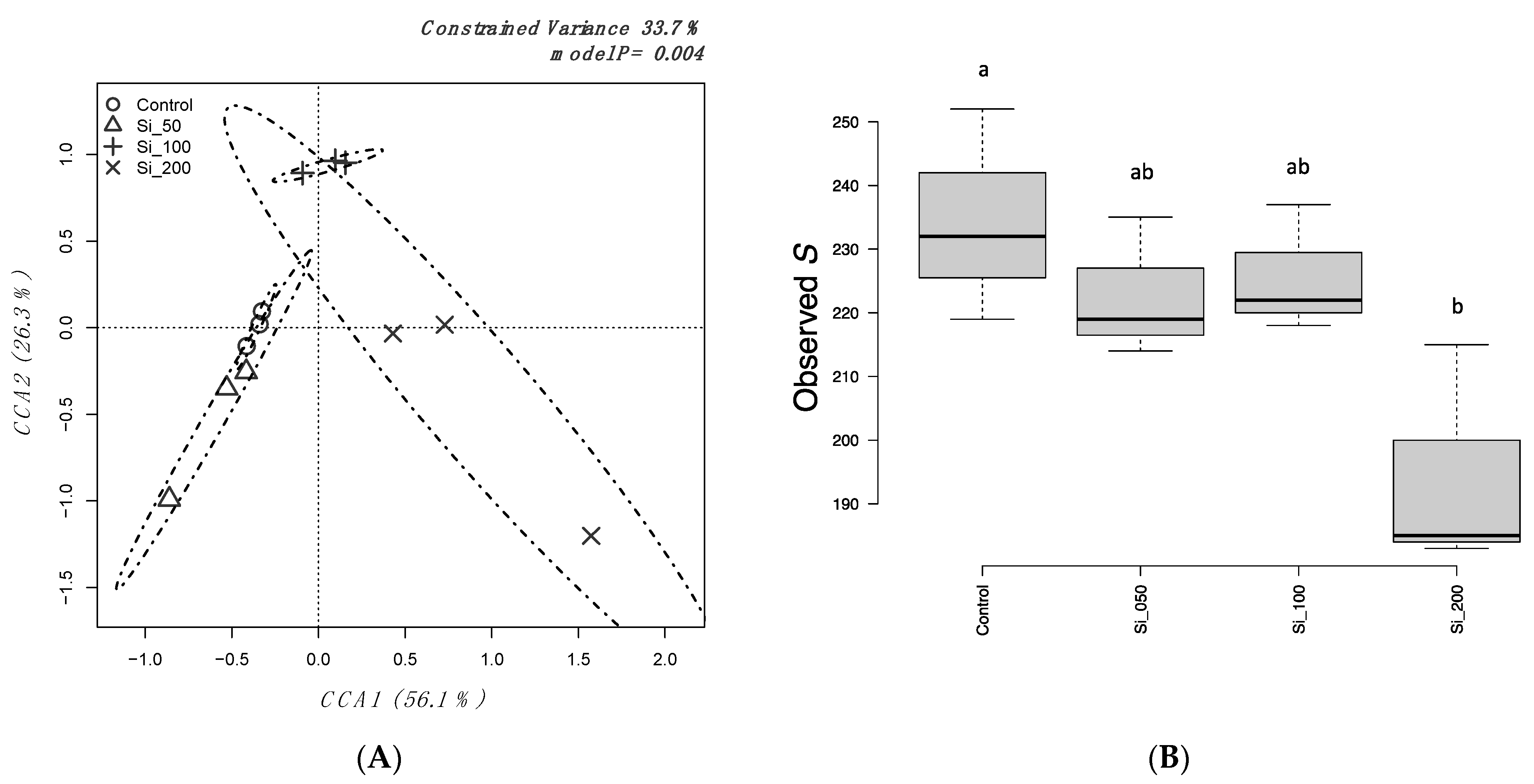
3.3. Bacterial Diversity of Harvested Strawberries
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bailey, R.L.; West, K.P.; Black, R.E. The epidemiology of global micronutrient deficiencies. Ann. Nutr. Metab. 2015, 66, 22–33. [Google Scholar] [CrossRef]
- Hefferon, K.L. Nutritionally enhanced food crops; progress and perspectives. Int. J. Mol. Sci. 2015, 16, 3895–3914. [Google Scholar] [CrossRef]
- O’Hare, T.J. Biofortification of vegetables for the developed world. Acta Hortic. 2015, 1106, 1–8. [Google Scholar] [CrossRef]
- Tardy, A.-L.; Ballesta, A.A.; Yilmaz, G.C.; DAN, M.; Ramirez, D.M.; Lam, H.Y.; Azais-Braesco, V.; Pouteau, E. Adult’s dietary intakes of selected vitamins & minerals essential for energy metabolism and cognition: A comparison across countries & genders (FS10-04-19). Curr. Dev. Nutr. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Alina, V.R.; Carmen, M.C.; Sevastita, M.; Andruţa, M.; Vlad, M.; Ramona, S.; Georgiana, P.; Mihaela, M. Food Fortification through Innovative Technologies. In Food Engineering; IntechOpen: London, UK, 2019; pp. 1–25. ISBN 978-1-83881-976-7. [Google Scholar]
- Mimmo, T.; Tiziani, R.; Valentinuzzi, F.; Lucini, L.; Nicoletto, C.; Sambo, P.; Scampicchio, M.; Pii, Y.; Cesco, S. Selenium biofortification in Fragaria x ananassa: Implications on strawberry fruits quality, content of bioactive health beneficial compounds and metabolomic profile. Front. Plant Sci. 2017, 8, 1887. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Sambo, P.; Nicoletto, C.; Giro, A.; Pii, Y.; Valentinuzzi, F.; Mimmo, T.; Lugli, P.; Orzes, G.; Mazzetto, F.; Astolfi, S.; et al. Hydroponic solutions for soilless production systems: Issues and opportunities in a smart agriculture perspective. Front. Plant Sci. 2019, 10, 923. [Google Scholar] [CrossRef]
- Price, C.T.; Koval, K.J.; Langford, J.R. Silicon: A review of its potential role in the prevention and treatment of postmenopausal osteoporosis. Int. J. Endocrinol. 2013, 2013, 316783. [Google Scholar] [CrossRef] [Green Version]
- D’Imperio, M.; Renna, M.; Cardinali, A.; Buttaro, D.; Santamaria, P.; Serio, F. Silicon biofortification of leafy vegetables and its bioaccessibility in the edible parts. J. Sci. Food Agric. 2016, 96, 751–756. [Google Scholar] [CrossRef]
- Tubaῆa, B.S.; Heckman, J.R. Silicon in soils and plants. In Silicon and Plant Diseases; Springer: Cham, Switzerland, 2015; pp. 7–51. ISBN 9783319229300. [Google Scholar]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- López-Pérez, M.C.; Pérez-Labrada, F.; Ramírez-Pérez, L.J.; Juárez-Maldonado, A.; Morales-Díaz, A.B.; González-Morales, S.; García-Dávila, L.R.; García-Mata, J.; Benavides-Mendoza, A. Dynamic modeling of silicon bioavailability, uptake, transport, and accumulation: Applicability in improving the nutritional quality of tomato. Front. Plant Sci. 2018, 9, 647. [Google Scholar] [CrossRef] [Green Version]
- Gattullo, C.E.; Allegretta, I.; Medici, L.; Fijan, R.; Pii, Y.; Cesco, S.; Mimmo, T.; Terzano, R. Silicon dynamics in the rhizosphere: Connections with iron mobilization. J. Plant Nutr. Soil Sci. 2016, 179, 409–417. [Google Scholar] [CrossRef]
- Raven, J.A. Cycling silicon—The role of accumulation in plants. New Phytol. 2003, 158, 419–421. [Google Scholar] [CrossRef]
- Kaushik, P.; Saini, D.K. Silicon as a vegetable crops modulator—A review. Plants 2019, 8, 148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laane, H.M. The effects of foliar sprays with different silicon compounds. Plants 2018, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buturi, C.V.; Mauro, R.P.; Fogliano, V.; Leonardi, C.; Giuffrida, F. Mineral biofortification of vegetables as a tool to improve human diet. Foods 2021, 10, 223. [Google Scholar] [CrossRef]
- D’Imperio, M.; Brunetti, G.; Gigante, I.; Serio, F.; Santamaria, P.; Cardinali, A.; Colucci, S.; Minervini, F. Integrated in vitro approaches to assess the bioaccessibility and bioavailability of silicon-biofortified leafy vegetables and preliminary effects on bone. In Vitro Cell. Dev. Biol.-Anim. 2017, 53, 217–224. [Google Scholar] [CrossRef]
- Gottardi, S.; Iacuzzo, F.; Tomasi, N.; Cortella, G.; Manzocco, L.; Pinton, R.; Römheld, V.; Mimmo, T.; Scampicchio, M.; Dalla Costa, L.; et al. Beneficial effects of silicon on hydroponically grown corn salad (Valerianella locusta (L.) Laterr) plants. Plant Physiol. Biochem. 2012, 56, 14–23. [Google Scholar] [CrossRef]
- Montesano, F.F.; D’Imperio, M.; Parente, A.; Cardinali, A.; Renna, M.; Serio, F. Green bean biofortification for Si through soilless cultivation: Plant response and Si bioaccessibility in pods. Sci. Rep. 2016, 6, 31662. [Google Scholar] [CrossRef] [Green Version]
- Valentinuzzi, F.; Cologna, K.; Pii, Y.; Mimmo, T.; Cesco, S. Assessment of silicon biofortification and its effect on the content of bioactive compounds in strawberry (Fragaria x ananassa ‘Elsanta’) fruits. Acta Hortic. 2018, 1217, 307–312. [Google Scholar] [CrossRef]
- Gerasopoulos, D.; Chouliaras, V.; Lionakis, S. Effects of preharvest calcium chloride sprays on maturity and storability of Hayward kiwifruit. Postharvest Biol. Technol. 1996, 7, 65–72. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Saladié, M.; Matas, A.J.; Isaacson, T.; Jenks, M.A.; Goodwin, S.M.; Niklas, K.J.; Xiaolin, R.; Labavitch, J.M.; Shackel, K.A.; Fernie, A.R.; et al. A reevaluation of the key factors that influence tomato fruit softening and integrity. Plant Physiol. 2007, 144, 1012–1028. [Google Scholar] [CrossRef] [Green Version]
- D’Amato, R.; De Feudis, M.; Hasuoka, P.E.; Regni, L.; Pacheco, P.H.; Onofri, A.; Businelli, D.; Proietti, P. The selenium supplementation influences olive tree production and oil stability against oxidation and can alleviate the water deficiency effects. Front. Plant Sci. 2018, 9, 1191. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Dubé, C.; Fang, C.; Roussel, D.; Charles, M.T.; Desjardins, Y.; Khanizadeh, S. Effect of production systems on phenolic composition and oxygen radical absorbance capacity of “Orléans” strawberry. LWT-Food Sci. Technol. 2012, 45, 241–245. [Google Scholar] [CrossRef]
- Alexandre, E.M.C.; Brandão, T.R.S.; Silva, C.L.M. Efficacy of non-thermal technologies and sanitizer solutions on microbial load reduction and quality retention of strawberries. J. Food Eng. 2012, 108, 417–426. [Google Scholar] [CrossRef]
- Baka, M.; Mercier, J.; Corcuff, R.; Castaigne, F.; Arul, J. Photochemical treatment to improve storability of fresh strawberries. J. Food Sci. 1999, 64, 1068–1072. [Google Scholar] [CrossRef]
- Bialka, K.L.; Demirci, A. Efficacy of aqueous ozone for the decontamination of Escherichia coli O157:H7 and Salmonella on raspberries and strawberries. J. Food Prot. 2007, 70, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Molina, F.; Gómez, P.L.; Castro, M.A.; Alzamora, S.M. Storage quality of strawberry fruit treated by pulsed light: Fungal decay, water loss and mechanical properties. Innov. Food Sci. Emerg. Technol. 2016, 34, 267–274. [Google Scholar] [CrossRef]
- Contigiani, E.V.; Jaramillo-Sánchez, G.; Castro, M.A.; Gómez, P.L.; Alzamora, S.M. Postharvest quality of strawberry fruit (Fragaria x ananassa Duch cv. Albion) as affected by ozone washing: Fungal spoilage, mechanical properties, and structure. Food Bioprocess Technol. 2018, 11, 1639–1650. [Google Scholar] [CrossRef]
- Jensen, B.; Knudsen, I.M.B.; Andersen, B.; Nielsen, K.F.; Thrane, U.; Jensen, D.F.; Larsen, J. Characterization of microbial communities and fungal metabolites on field grown strawberries from organic and conventional production. Int. J. Food Microbiol. 2013, 160, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Zhimo, V.Y.; Kumar, A.; Biasi, A.; Salim, S.; Feygenberg, O.; Toamy, M.A.; Abdelfattaah, A.; Medina, S.; Freilich, S.; Wisniewski, M.; et al. Compositional shifts in the strawberry fruit microbiome in response to near-harvest application of Metschnikowia fructicola, a yeast biocontrol agent. Postharvest Biol. Technol. 2021, 175, 111469. [Google Scholar] [CrossRef]
- Pelissari, E.M.R.; Covre, K.V.; do Rosario, D.K.A.; de São José, J.F.B. Application of chemometrics to assess the influence of ultrasound and chemical sanitizers on vegetables: Impact on natural microbiota, Salmonella enteritidis and physicochemical nutritional quality. LWT 2021, 148, 111711. [Google Scholar] [CrossRef]
- Darlison, J.; Mogren, L.; Rosberg, A.K.; Grudén, M.; Minet, A.; Liné, C.; Mieli, M.; Bengtsson, T.; Håkansson, Å.; Uhlig, E.; et al. Leaf mineral content govern microbial community structure in the phyllosphere of spinach (Spinacia oleracea) and rocket (Diplotaxis tenuifolia). Sci. Total Environ. 2019, 675, 501–512. [Google Scholar] [CrossRef]
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Vasileiadis, S.; Puglisi, E.; Trevisan, M.; Scheckel, K.G.; Langdon, K.A.; McLaughlin, M.J.; Lombi, E.; Donner, E. Changes in soil bacterial communities and diversity in response to long-term silver exposure. FEMS Microbiol. Ecol. 2015, 91, fiv114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Keller, K.; Brodie, E.L.; Larsen, N.; Piceno, Y.M.; Phan, R.; Andersen, G.L. NAST: A multiple sequence alignment server for comparative analysis of 16S rRNA genes. Nucleic Acids Res. 2006, 34, W394–W399. [Google Scholar] [CrossRef]
- Schloss, P.D. The effects of alignment quality, distance calculation method, sequence filtering, and region on the analysis of 16S rRNA gene-based studies. PLoS Comput. Biol. 2010, 6, e1000844. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; Desantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016; ISBN 3319242776. [Google Scholar]
- De Mendiburu, F. Package ‘agricolae’. In R Package Version; 2019; pp. 1–2. Available online: https://cran.r-project.org/web/packages/agricolae/index.html (accessed on 7 September 2021).
- Tang, Y.; Horikoshi, M.; Li, W. Ggfortify: Unified interface to visualize statistical results of popular r packages. R J. 2016, 8, 478–489. [Google Scholar] [CrossRef] [Green Version]
- Paulson, J.N.; Pop, M.; Bravo, H.C. Metastats: An improved statistical method for analysis of metagenomic data. Genome Biol. 2011, 12, P17. [Google Scholar] [CrossRef] [Green Version]
- Pistón, F.; Pérez, A.G.; Sanz, C.; Refoyo, A. Strawberry postharvest shelf life is related to total acid content and fruit firmness. In IX International Strawberry Symposium; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2021; pp. 869–872. [Google Scholar]
- Chiabrando, V.; Garavaglia, L.; Giacalone, G. The postharvest quality of fresh sweet cherries and strawberries with an active packaging system. Foods 2019, 8, 335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emongor, V.E.; Ramagonono, G. Storage temperature influences postharvest quality of wild plum (Ximmenia Americana L.) Fruit. Ghana J. Sci. 2019, 60, 1–10. [Google Scholar] [CrossRef]
- Da Silva, E.P.; Cardoso, A.F.L.; Fante, C.; Rosell, C.M.; de Boas, E.V.B.V. Effect of postharvest temperature on the shelf life of gabiroba fruit (Campomanesia pubescens). Food Sci. Technol. 2013, 33, 632–637. [Google Scholar] [CrossRef] [Green Version]
- Wescombe, P.A.; Hale, J.D.; Heng, N.C.; Tagg, J.R. Developing oral probiotics from Streptococcus salivarius. Future Microbiol. 2012, 7, 1355–1371. [Google Scholar] [CrossRef]
- Song, J.; Lim, H.X.; Lee, A.; Kim, S.; Lee, J.H.; Kim, T.S. Staphylococcus succinus 14BME20 prevents allergic airway inflammation by induction of regulatory T Cells via interleukin-10. Front. Immunol. 2019, 10, 1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syuhadah, A.S.N.; Ali, U.K.; Norazizah, M. A rare and fatal case of Terrisporobacter glycolicus bacteremia: Case report and review of literature. Int. J. Infect. Dis. 2020, 101, 151. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | Si 50 | Si 100 | Si 200 | |
|---|---|---|---|---|
| FW shoot (g plant−1) ** | 20.59 ± 2.35 a | 13.42 ± 0.88 b | 14.88 ± 1.43 b | 10.08 ± 1.22 b |
| FW root (g plant−1) ** | 19.08 ± 1.20 a | 14.70 ± 1.28 b | 13.29 ± 0.49 b | 13.61 ± 0.93 b |
| Leaf area (cm2) ns | 38.28 ± 2.72 a | 34.85 ± 2.42 a | 36.41 ± 2.16 a | 30.23 ± 1.95 a |
| Average n. of flowers ns | 21.89 ± 2.45 ns | 23.78 ± 1.36 ns | 21.78 ± 1.86 ns | 22.38 ± 2.27 ns |
| Average yield plant−1 (g) * | 60.22 ± 5.69 a | 49.70 ± 5.46 ab | 41.85 ± 2.46 b | 37.10 ± 4.52 b |
| Average n. of berries plant−1 * | 16.00 ± 0.93 a | 14.67 ± 1.64 a | 13.44 ± 0.84 ab | 12.25 ± 1.35 b |
| Average berry weight (g) *** | 3.71 ± 0.18 ab | 3.42 ± 0.17 b | 4.13 ± 0.22 a | 2.79 ± 0.19 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valentinuzzi, F.; Pii, Y.; Borruso, L.; Mimmo, T.; Puglisi, E.; Trevisan, M.; Cesco, S. Epiphytic Microbial Community and Post-Harvest Characteristics of Strawberry Fruits as Affected by Plant Nutritional Regime with Silicon. Agronomy 2021, 11, 2407. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122407
Valentinuzzi F, Pii Y, Borruso L, Mimmo T, Puglisi E, Trevisan M, Cesco S. Epiphytic Microbial Community and Post-Harvest Characteristics of Strawberry Fruits as Affected by Plant Nutritional Regime with Silicon. Agronomy. 2021; 11(12):2407. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122407
Chicago/Turabian StyleValentinuzzi, Fabio, Youry Pii, Luigimaria Borruso, Tanja Mimmo, Edoardo Puglisi, Marco Trevisan, and Stefano Cesco. 2021. "Epiphytic Microbial Community and Post-Harvest Characteristics of Strawberry Fruits as Affected by Plant Nutritional Regime with Silicon" Agronomy 11, no. 12: 2407. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122407