
Evaluation of Crossability between Nicotiana benthamiana and Nicotiana excelsior


1
Graduate School of Life and Environmental Sciences, Osaka Prefecture University, Sakai 599-8531, Osaka, Japan
2
Education and Research Field, College of Life, Environment and Advanced Sciences, Osaka Prefecture University, Sakai 599-8531, Osaka, Japan
*
Author to whom correspondence should be addressed.
Agronomy 2021, 11(12), 2583; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122583
Submission received: 17 November 2021
/
Revised: 14 December 2021
/
Accepted: 17 December 2021
/
Published: 19 December 2021
(This article belongs to the Section Crop Breeding and Genetics)
Abstract
:Wild tobacco species in the Nicotiana section Suaveolentes are promising genetic resources to introduce their disease resistance to cultivated tobacco, Nicotiana tabacum. However, hybrid lethality is observed in hybrid seedlings from crosses between most Suaveolentes species and N. tabacum. In particular, N. benthamiana belonging to the section Suaveolentes produces only viable hybrids after crossing with N. tabacum. In the present study, crossability between N. benthamiana and N. excelsior (section Suaveolentes) was investigated to test the possible usefulness of N. benthamiana as the bridge parent to transfer desirable genes of N. excelsior to N. tabacum via bridge crossing. After reciprocal crosses using three accessions of N. benthamiana and N. excelsior each, several crossing barriers such as cross-incompatibility, seed abortion, and male and female hybrid sterility were observed. Although reciprocal hybrids between N. benthamiana and N. excelsior showed a high degree of chromosome pairing in meiosis, univalents and multivalents, as well as chromosome bridges and lagging chromosomes, were observed. These meiotic abnormalities were thought to cause hybrid sterility. The possible usefulness of reciprocal hybrids between N. benthamiana and N. excelsior is discussed.
1. Introduction
An interspecific cross, regardless of whether it is naturally occurring or artificially performed, is an important evolutionary mechanism. Interspecific crosses may produce new species in nature, and several plant species are amphidiploids possessing two or more genomes that naturally arise from interspecific crosses and polyploidization [1,2,3]. In artificial crosses, interspecific crosses are widely used for breeding purposes or genetic analysis [4,5,6]. Crossability between species depends on their genotypes, ploidy levels, and environmental factors.
Nicotiana benthamiana is a model plant species, especially in plant virology, because genetic transformation techniques and tissue culture methods have been well established [7], and a draft genome sequence has been reported [8]. This species belongs to the Nicotiana section Suaveolentes, which consists of wild tobacco species restricted to Australasia and one species in Africa [9]. Species in the section Suaveolentes are allotetraploids with 30 to 48 chromosomes, excluding species that experienced a possible additional chromosome doubling event [10,11,12,13,14]. This section is predicted to have arisen ca. 6 Mya, and most Suaveolentes species have appeared within the last 2 Myr [13,15]. Goodspeed [10] presumed that crosses between early Suaveolentes species and subsequent backcrosses or crosses of derivatives, as well as chromosomal reorganization, were involved in the formation of the current diverse species in the section. Based on the distribution, morphological specificity, and the somewhat low chromosome pairing in hybrids with other Suaveolentes species, N. benthamiana apparently has undergone a long period of independent differentiation [10].
N. benthamiana is one of the two species possessing 38 chromosomes in the section Suaveolentes; the other is N. excelsior. While N. benthamiana is susceptible to a wide range of pathogens and viruses [7], most other species in the section Suaveolentes possess disease resistance. For example, N. excelsior shows resistance to blue mold (Peronospora hyoscyami f.sp. tabacina), powdery mildew, and a specific strain of potato virus Y [16,17,18]. Therefore, Suaveolentes species are promising genetic resources to introduce their disease resistance to cultivated tobacco, N. tabacum (section Nicotiana). However, the transfer of desirable genes of Suaveolentes species into N. tabacum has been disrupted by reproductive isolation [19]. Hybrid lethality, a type of postzygotic reproductive isolation, is commonly observed in crosses between N. tabacum and most Suaveolentes species, including N. excelsior [14,19,20,21,22]. Hybrid lethality is a phenomenon that causes the death of F1 hybrid seedlings. In particular, N. benthamiana is one of the two species yielding only viable hybrid seedlings after crossing with N. tabacum [23]. If N. benthamiana can be used as a bridge species to introduce desirable genes from Suaveolentes species to N. tabacum, tobacco breeding will be facilitated. However, while chromosome pairing in intrasectional F1 hybrids of the section Suaveolentes has been well reported, as mentioned in the discussion, crossability between Suaveolentes species has been scarcely reported.
In the present study, we investigated the crossability between N. benthamiana and N. excelsior to test whether N. benthamiana can be used as a bridge species to transfer desirable genes of N. excelsior into N. tabacum. Using three accessions of N. benthamiana and N. excelsior each, reciprocal crosses were performed. Hybrid seedlings obtained were investigated for pollen fertility, reciprocally backcrossed to both parents, and evaluated for chromosome behavior during meiosis. We discuss the possible usefulness of N. benthamiana as a bridge species in breeding programs.
2. Materials and Methods
2.1. Plant Materials
N. benthamiana (2n = 38) accessions JT, PI 555478, and PI 555684, and N. excelsior (2n = 38) accessions JT, PI 224063, and PI 555685 were used for interspecific crosses. The accessions obtained from the Leaf Tobacco Research Center (Japan Tobacco Inc., Oyama, Japan) were named JT to distinguish them from the United States Department of Agriculture (USDA) Plant Introduction (PI) accessions. All plants used as parents were cultivated in a greenhouse under natural light conditions.
2.2. Interspecific Crosses
N. benthamiana and N. excelsior were reciprocally crossed. Flowers of plants used as female parents were emasculated one day before anthesis and pollinated with pollen from plants used as male parents. F1 seeds were sterilized with 5% sodium hypochlorite for 15 min. The sterilized seeds were sown in Petri dishes (90 mm diameter, 17 mm deep) containing 25 mL of 1/2 MS medium [24] supplemented with 1% sucrose and 0.2% Gelrite (pH 5.8) and then cultured at 25 °C (16 h light/8 h dark; approximately 80 µmol m−2 s−1). Hybrid seedlings germinated in Petri dishes were potted and cultivated in a greenhouse. The hybrid plants were reciprocally backcrossed with both parents. The number of capsules and hybrids obtained were counted for each cross.
2.3. Analysis of Pollen Viability
Flower buds were collected one day before anthesis, and the anthers were placed on glass slides. A drop of acetocarmine was added, and the anthers were crushed with tweezers to release pollen grains. At least 100 pollen grains were observed per anther using a light microscope (BX50; Olympus, Tokyo, Japan), and three anthers were observed per hybrid plant. Stained pollen grains were considered viable, whereas unstained pollen grains were considered inviable.
2.4. Cytological Analysis of Chromosomes in Meiosis
Chromosomes in pollen mother cells during meiosis were also investigated. Anthers collected from young flower buds were fixed in chloroform/ethanol/acetic acid (6:3:1). The anthers were then placed on a glass slide and squashed in acetocarmine. Chromosome pairing during meiotic metaphase I was observed in 25 pollen mother cells per cross combination using a light microscope (Optiphot-2; Nikon, Tokyo, Japan). Chromosomes were also observed at meiotic anaphase I.
3. Results
3.1. Reciprocal Crosses between N. benthamiana and N. excelsior
Reciprocal crosses were carried out between three N. benthamiana accessions and three N. excelsior accessions (Table 1). The percentage of capsules obtained after pollination was low for all reciprocal crosses. Although seeds were obtained from 14 of the 18 crosses, several crosses yielded a small number of seeds. In this case, all the seeds obtained were aseptically sown. No seeds germinated in crosses between N. benthamiana (♀) and N. excelsior (♂), excluding two crosses with seed germination rates of 1% and 33%. Conversely, seed germination rates in crosses between N. excelsior (♀) and N. benthamiana (♂) were high (85–96%), although the rates of the two crosses were low (18%) and moderate (55%). All hybrid seedlings obtained from the nine crosses reached maturity. Leaf and flower shapes were intermediate in appearance to those of the parents (Figure 1).
3.2. Fertility of Hybrids between N. benthamiana and N. excelsior
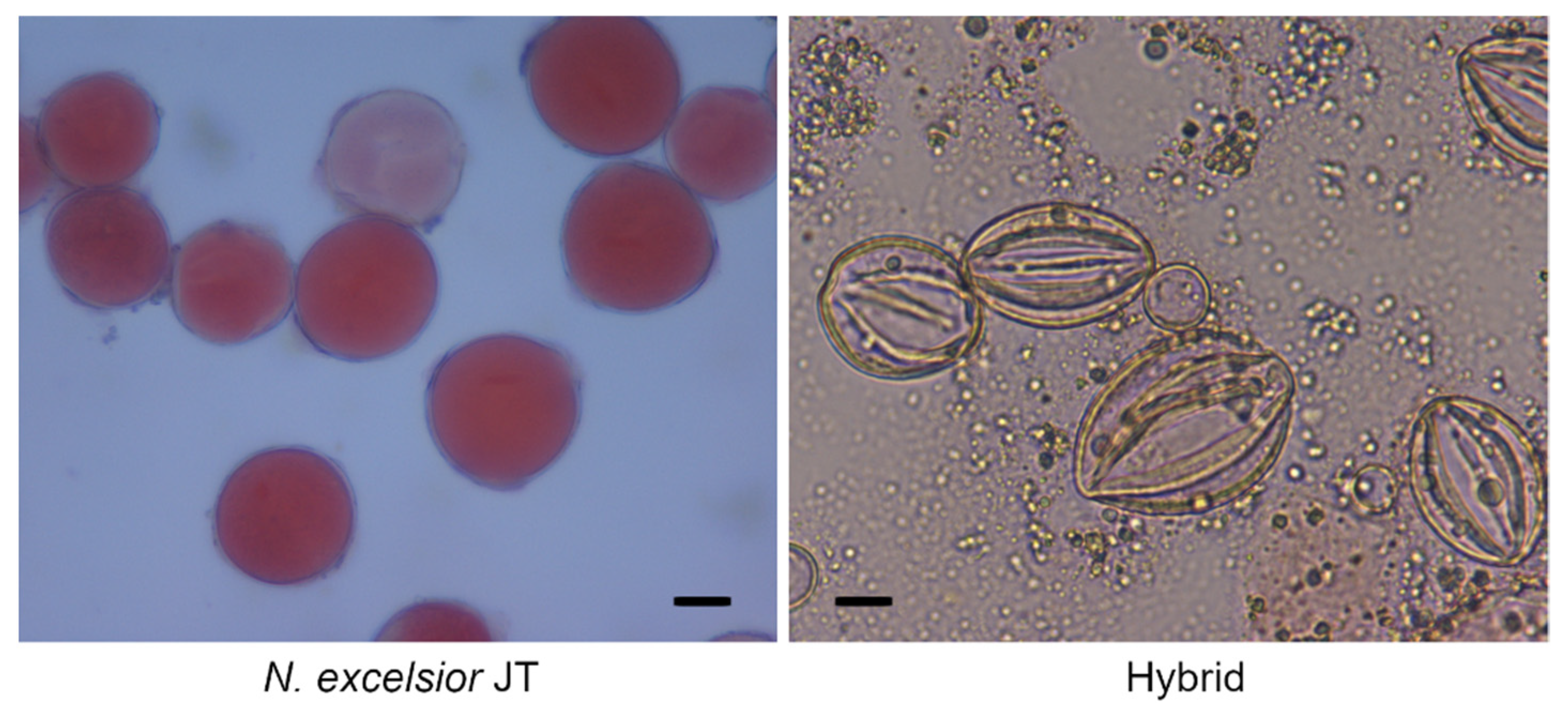
The pollen viability of the hybrid plants was investigated by staining with acetocarmine. More than 90% of the pollen grains were stained and considered viable in both parents (Figure 2). However, pollen grains with irregular shapes were observed, and none of the pollen grains were stained in all hybrid plants from the nine crosses, suggesting pollen inviability (Figure 2 and Table 2).
Hybrid plants from all nine crosses were backcrossed to both parents as male parents. However, seeds could not be obtained from all crosses (Table 3). Seeds were also not obtained in backcrosses of hybrid plants from four crosses to both parents as female parents, as predicted by pollen staining (Table 4).
3.3. Chromosomal Instability in Reciprocal Hybrids between N. benthamiana and N. excelsior
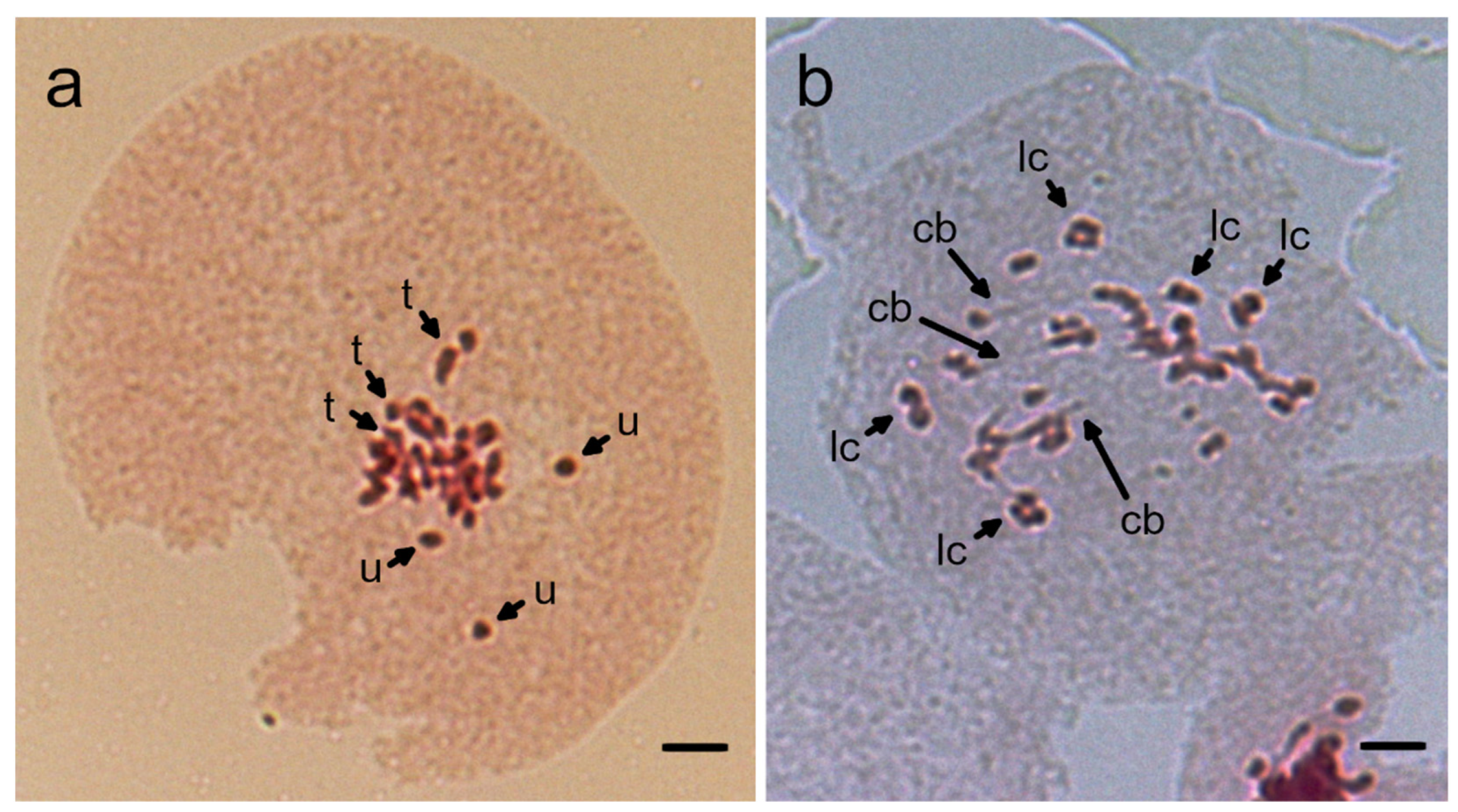
Chromosomes of reciprocal hybrids between N. benthamiana JT and N. excelsior JT were examined during meiosis. At meiotic metaphase I, chromosome pairing with an average of 3.5 univalents, 12.8 bivalents, 2.4 trivalents, and 0.1 tetravalents per pollen mother cell was observed in hybrid plants from the cross N. benthamiana JT × N. excelsior JT (Figure 3a and Table 5). In hybrid plants with the opposite direction of hybridization, chromosome pairing with an average of 2.8 univalents, 14.0 bivalents, and 2.4 trivalents was observed per pollen mother cell (Table 5). When chromosomes at meiotic anaphase I were observed in hybrids from the cross N. benthamiana JT × N. excelsior JT, chromosome bridges and lagging chromosomes, which are aberration phenomena, were observed (Figure 3b).
4. Discussion
Crossing results in the present study indicate that several isolation barriers exist between N. benthamiana and N. excelsior. In reciprocal crosses, the percentage of capsules obtained was low, suggesting the presence of prezygotic cross-incompatibility (Table 1). Excluding reciprocal crosses between N. benthamiana PI 555684 and N. excelsior JT, seed germination percentages were generally lower in crosses using N. benthamiana as maternal parents than in the opposite crosses (Table 1). These differences between reciprocal crosses are caused by seed abortion, which is observed unilaterally. Although all obtained hybrid seedlings matured, they were both male and female sterile (Figure 2 and Table 2, Table 3 and Table 4).
In the Nicotiana section Suaveolentes, a high degree of chromosome pairing in meiosis has been reported in several interspecific hybrids [10,22,25,26,27,28,29,30,31,32]. Reciprocal hybrids between N. benthamiana JT and N. excelsior JT also showed a high degree of chromosome pairing, suggesting high homology of the genomes. However, in addition to bivalents, univalents, trivalents, and tetravalents, as well as chromosome bridges and lagging chromosomes, were observed (Figure 3 and Table 5). Hybrid sterility can be caused by meiotic irregularities, chromosomal aberrations, or interactions between nuclear genes or between the cytoplasm and nuclear genes [33,34,35,36,37]. Although formation of the multivalents suggested some homologies between corresponding chromosomes as inferred from the monophyly of Suaveolentes species and a presumption that the species are allotetraploids derived from common ancestors [10,13,15,38], the univalents and multivalents, as well as chromosome bridges and lagging chromosomes, would lead to imbalanced chromosome segregation, resulting in hybrid sterility in crosses between N. benthamiana and N. excelsior. It was also reported that because amphiploids produced by chromosome doubling of sterile F1 hybrids between Suaveolentes species showed full fertility, sterility in the F1 hybrids can be attributed to chromosomal causes [39].
In crosses between N. excelsior and N. tabacum, the following several crossing barriers were observed: the prezygotic barrier preventing fertilization and the postzygotic barrier during seed development in the cross using N. excelsior as the male parent, and hybrid lethality in reciprocal crosses [19]. Meanwhile, N. benthamiana produced viable hybrid plants when crossed using N. tabacum as the male parent, although the reciprocal cross was unsuccessful because of the prezygotic barrier [23]. These facts promise to transfer desirable genes of N. excelsior into N. tabacum using N. benthamiana as a bridge species. However, both male and female sterility in reciprocal hybrids between N. benthamiana and N. excelsior would disturb the bridge cross. Several reports have provided useful information to address this problem. Hybrid sterility can often be overcome by chromosome doubling methods using colchicine treatment or tissue culture [40,41,42]. As mentioned above, amphiploids derived from several sterile intrasectional hybrids in the section Suaveolentes also showed regular chromosome pairing in a majority of the pollen mother cells and full fertility [39]. Each chromosome is considered to have exact copy and chromosomal homology by chromosome doubling, leading to fertility restoration [42]. Therefore, fertility can be restored through the production of amphiploids from reciprocal hybrids between N. benthamiana and N. excelsior. If this is accomplished, hybrids obtained in the present study would be useful for transferring the desirable genes of N. excelsior to N. tabacum.
5. Conclusions
Several crossing barriers such as cross-incompatibility, seed abortion, and male and female hybrid sterility were observed in reciprocal crosses using three accessions of N. benthamiana and N. excelsior each. Although reciprocal hybrids between N. benthamiana and N. excelsior showed a high degree of chromosome pairing in meiosis, univalents and multivalents, as well as chromosome bridges and lagging chromosomes, were observed. These meiotic abnormalities were thought to cause hybrid sterility.
Author Contributions
Conceptualization, T.T. and N.K.; methodology, T.T., N.K. and T.M.; validation, T.T., N.K., M.Y. and T.M.; formal analysis, T.T., N.K. and T.M.; investigation, N.K.; writing—original draft preparation, T.T.; writing—review and editing, T.T.; visualization, T.T. and N.K.; supervision, T.T., M.Y. and T.M.; project administration, T.T.; funding acquisition, T.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partly funded by JSPS KAKENHI Grant Numbers JP20880024, JP25870627, JP20K05988 from the Japan Society for the Promotion of Science.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank the Leaf Tobacco Research Center, Japan Tobacco Inc., Oyama, Japan, and the United States Nicotiana Germplasm Collection for providing seeds of Nicotiana species.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Abbott, R.J. Plant invasions, interspecific hybridization and the evolution of new plant taxa. Trends Ecol. Evol. 1992, 7, 401–405. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Willis, J.H. Plant Speciation. Science 2007, 317, 910–914. [Google Scholar] [CrossRef]
- Rodionov, A.V.; Amosova, A.V.; Belyakov, E.A.; Zhurbenko, P.M.; Mikhailova, Y.V.; Punina, E.O.; Shneyer, V.S.; Loskutov, I.G.; Muravenko, O.V. Genetic Consequences of Interspecific Hybridization, Its Role in Speciation and Phenotypic Diversity of Plants. Russ. J. Genet. 2019, 55, 278–294. [Google Scholar] [CrossRef]
- Newaskar, G.S.; Chimote, V.P.; Mehetre, S.S.; Jadhav, A.S. Interspecific hybridization in Gossypium L.: Characterization of progenies with different ploidy-confirmed multigenomic backgrounds. Plant Breed. 2013, 132, 211–216. [Google Scholar] [CrossRef]
- Plazas, M.; Vilanova, S.; Gramazio, P.; Rodríguez-Burruezo, A.; Fita, A.; Herraiz, F.J.; Ranil, R.; Fonseka, R.; Niran, L.; Fonseka, H.; et al. Interspecific Hybridization between Eggplant and Wild Relatives from Different Genepools. J. Am. Soc. Hortic. Sci. 2016, 141, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Marasek-Ciolakowska, A.; Nishikawa, T.; Shea, D.J.; Okazaki, K. Breeding of lilies and tulips—Interspecific hybridization and genetic background. Breed. Sci. 2018, 68, 35–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodin, M.M.; Zaitlin, D.; Naidu, R.A.; Lommel, S.A. Nicotiana benthamiana: Its History and Future as a Model for Plant–Pathogen Interactions. Mol. Plant-Microbe Interact. 2008, 21, 1015–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bombarely, A.; Rosli, H.G.; Vrebalov, J.; Moffett, P.; Mueller, L.A.; Martin, G.B. A Draft Genome Sequence of Nicotiana benthamiana to Enhance Molecular Plant-Microbe Biology Research. Mol. Plant-Microbe Interact. 2012, 25, 1523–1530. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Chase, M.W.; Clarkson, J.J. Nomenclatural changes and a new sectional classification in Nicotiana (Solanaceae). Taxon 2004, 53, 73–82. [Google Scholar] [CrossRef]
- Goodspeed, T.H. The Genus Nicotiana; Chronica Botanica Company: Waltham, MA, USA, 1954. [Google Scholar]
- Marks, C.E.; Ladiges, P.Y.; Newbigin, E. Karyotypic variation in Nicotiana section Suaveolentes. Genet. Resour. Crop. Evol. 2011, 58, 797–803. [Google Scholar] [CrossRef]
- He, H.; Iizuka, T.; Maekawa, M.; Sadahisa, K.; Morikawa, T.; Yanase, M.; Yokoi, S.; Oda, M.; Tezuka, T. Nicotiana suaveolens accessions with different ploidy levels exhibit different reproductive isolation mechanisms in interspecific crosses with Nicotiana tabacum. J. Plant Res. 2019, 132, 461–471. [Google Scholar] [CrossRef]
- Dodsworth, S.; Christenhusz, M.J.M.; Conran, J.G.; Guignard, M.S.; Knapp, S.; Struebig, M.; Leitch, A.R.; Chase, M.W. Extensive plastid-nuclear discordance in a recent radiation of Nicotiana section Suaveolentes (Solanaceae). Bot. J. Linn. Soc. 2020, 193, 546–559. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Ohya, Y.; Maekawa, M.; Iizuka, T.; Hasegawa, A.; Shiragaki, K.; He, H.; Oda, M.; Morikawa, T.; Yokoi, S.; et al. Two Nicotiana occidentalis accessions enable gene identification for Type II hybrid lethality by the cross to N. sylvestris. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Clarkson, J.J.; Dodsworth, S.; Chase, M.W. Time-calibrated phylogenetic trees establish a lag between polyploidisation and diversification in Nicotiana (Solanaceae). Plant Syst. Evol. 2017, 303, 1001–1012. [Google Scholar] [CrossRef]
- Burk, L.G.; Heggestad, H.E. The genusNicotiana: A source of resistance to diseases of cultivated tobacco. Econ. Bot. 1966, 20, 76–88. [Google Scholar] [CrossRef]
- Gillham, F.; Wark, D.; Harrigan, E. Disease resistant flue-cured tobacco breeding lines for north Queensland I. Resistance to blue mould, Peronospora tabacina. Aust. J. Exp. Agric. 1977, 17, 652–658. [Google Scholar] [CrossRef]
- Dijk, P.; Cuperus, C. Reactions on Nicotiana species to potato viruses A, X and Y and tobacco mosaic virus in relation to their taxonomy and geographical origin. Neth. J. Plant Pathol. 1989, 95, 343–356. [Google Scholar] [CrossRef]
- Tezuka, T.; Kuboyama, T.; Matsuda, T.; Marubashi, W. Seven of eight species in Nicotiana section Suaveolentes have common factors leading to hybrid lethality in crosses with Nicotiana tabacum. Ann. Bot. 2010, 106, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tezuka, T. Hybrid lethality in Nicotiana: A review with special attention to interspecific crosses between species in sect. Suaveolentes and N. tabacum. In Herbaceous Plants: Cultivation Methods, Grazing and Environmental Impacts; Wallner, F., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2013; pp. 69–94. [Google Scholar]
- Shiragaki, K.; Nakamura, R.; Nomura, S.; He, H.; Yamada, T.; Marubashi, W.; Oda, M.; Tezuka, T. Phenylalanine ammonia-lyase and phenolic compounds are related to hybrid lethality in the cross Nicotiana suaveolens × N. tabacum. Plant Biotechnol. 2020, 37, 327–333. [Google Scholar] [CrossRef]
- Tezuka, T.; Kitamura, N.; Imagawa, S.; Hasegawa, A.; Shiragaki, K.; He, H.; Yanase, M.; Ogata, Y.; Morikawa, T.; Yokoi, S. Genetic Mapping of the HLA1 Locus Causing Hybrid Lethality in Nicotiana Interspecific Hybrids. Plants 2021, 10, 2062. [Google Scholar] [CrossRef] [PubMed]
- Iizuka, T.; Kuboyama, T.; Marubashi, W.; Oda, M.; Tezuka, T. Nicotiana debneyi has a single dominant gene causing hybrid lethality in crosses with N. tabacum. Euphytica 2012, 186, 321–328. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Wheeler, H.-M. A Contribution to the Cytology of the Australian-South Pacific Species of Nicotiana. Proc. Natl. Acad. Sci. USA 1945, 31, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, Y. Cytogenetic studies of Nicotiana, V. Reduction divisions in hybrids Between. Bot. Mag. Tokyo 1953, 66, 269–276. [Google Scholar] [CrossRef]
- Goodspeed, T.H.; Thompson, M.C. Cytotaxonomy of Nicotiana. II. Bot. Rev. 1959, 25, 385–415. [Google Scholar] [CrossRef]
- Gopinath, D.M.; Krishnamurthy, K.V.; Krishnamurthy, A.S. Cytological studies on interspecific hybrids in nicotiana involving a new Australian species, Nicotiana amplexicaulis. Can. J. Genet. Cytol. 1965, 7, 328–340. [Google Scholar] [CrossRef]
- Williams, E. Meiotic Chromosome Pairing in Interspecific Hybrids of Nicotiana. N. Z. J. Bot. 1975, 13, 601–609. [Google Scholar] [CrossRef] [Green Version]
- Gerstel, D.U.; Burns, J.A.; Burk, L.G. Interspecific hybridizations with an African tobacco, Nicotiana africana Merxm. J. Hered. 1979, 70, 342–344. [Google Scholar] [CrossRef]
- Gangadevi, T.; Rao, P.N.; Satyanarayana, K.V. Cytogenetic study of an interspecific cross of Nicotiana debneyi X N. umbratica. Theor. Appl. Genet. 1982, 63, 177–181. [Google Scholar] [CrossRef]
- Gangadevi, T.; Rao, P.N.; Satyanarayana, K.V. Morphological and cytological studies of interspecific hybrids in Nicotiana involving N. umbratica Burbidge. Cytologia 1987, 52, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Kubo, T.; Kumashiro, T.; Saito, Y. Cytoplasmic male sterile lines of a tobacco variety, Tsukuba 1, developed by asymmetric protoplast fusion. Jpn. J. Breed. 1988, 38, 158–164. [Google Scholar] [CrossRef]
- Rieseberg, L.H.; Blackman, B.K. Speciation genes in plants. Ann. Bot. 2010, 106, 439–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stathos, A.; Fishman, L. Chromosomal rearrangements directly cause underdominant F1 pollen sterility in Mimulus lewisii–Mimulus cardinalis hybrids. Evolution 2014, 68, 3109–3119. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.E.Z.; Lin, H.-X. Evolution and Molecular Control of Hybrid Incompatibility in Plants. Front. Plant Sci. 2016, 7, 1208. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, J.; Zhang, Y.; Yang, Y.; Pu, Q.; Tao, D. New Insights Into the Nature of Interspecific Hybrid Sterility in Rice. Front. Plant Sci. 2020, 11, 555572. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.J.; Leitch, A.R.; Clarkson, J.J.; Knapp, S.; Chase, M.W. Reconstructing the complex evolutionary origin of wild allopolyploid tobaccos (Nicotiana section Suaveolentes). Evolution 2013, 67, 80–94. [Google Scholar] [CrossRef] [PubMed]
- Gangadevi, T.; Rao, P.N.; Satyanarayana, K.V. Cytogenetic Studies of Some Synthetic Amphiploids of Nicotiana. J. Hered. 1988, 79, 119–122. [Google Scholar] [CrossRef]
- Mariam, A.L.; Zakri, A.H.; Mahani, M.C.; Normah, M.N. Interspecific hybridization of cultivated rice, Oryza sativa L. with the wild rice, O. minuta Presl. Theor. Appl. Genet. 1996, 93, 664–671. [Google Scholar] [CrossRef]
- Chetelat, R.T. Overcoming sterility and unilateral incompatibility of Solanum lycopersicum × S. sitiens hybrids. Euphytica 2016, 207, 319–330. [Google Scholar] [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Leaves and flowers of reciprocal hybrids between N. benthamiana PI 555684 and N. excelsior JT. Scale bars = 1 cm.
Figure 1.
Leaves and flowers of reciprocal hybrids between N. benthamiana PI 555684 and N. excelsior JT. Scale bars = 1 cm.

Figure 2.
Pollen grains stained with acetocarmine in N. excelsior JT and a hybrid from the cross N. excelsior JT × N. benthamiana JT. Scale bars = 10 µm.
Figure 2.
Pollen grains stained with acetocarmine in N. excelsior JT and a hybrid from the cross N. excelsior JT × N. benthamiana JT. Scale bars = 10 µm.

Figure 3.
Chromosome instability in hybrids between N. benthamiana JT and N. excelsior JT. (a) Chromosome pairing in meiotic metaphase I. u, univalent; t, trivalent. (b) Chromosomes at meiotic anaphase I. cb, chromosome bridge; lc, lagging chromosome. Scale bars = 10 µm.
Figure 3.
Chromosome instability in hybrids between N. benthamiana JT and N. excelsior JT. (a) Chromosome pairing in meiotic metaphase I. u, univalent; t, trivalent. (b) Chromosomes at meiotic anaphase I. cb, chromosome bridge; lc, lagging chromosome. Scale bars = 10 µm.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Reciprocal crosses between N. benthamiana and N. excelsior.
| Cross Combination | No. of Flowers Pollinated | No. of Capsules Obtained | No. of Seeds Sown | No. of Hybrids Obtained |
|---|---|---|---|---|
| N. benthamiana JT × N. excelsior JT | 40 | 10 (25% 1) | 178 | 2 (1% 2) |
| N. benthamiana JT × N. excelsior PI 224063 | 11 | 2 (18%) | 48 | 0 (0%) |
| N. benthamiana JT × N. excelsior PI 555685 | 5 | 1 (20%) | 2 | 0 (0%) |
| N. benthamiana PI 555478 × N. excelsior JT | 20 | 0 (0%) | – | – |
| N. benthamiana PI 555478 × N. excelsior PI 224063 | 13 | 1 (8%) | 22 | 0 (0%) |
| N. benthamiana PI 555478 × N. excelsior PI 555685 | 20 | 1 (5%) | 143 | 0 (0%) |
| N. benthamiana PI 555684 × N. excelsior JT | 20 | 2 (%) | 27 | 9 (33%) |
| N. benthamiana PI 555684 × N. excelsior PI 224063 | 5 | 1 (20%) | 48 | 0 (0%) |
| N. benthamiana PI 555684 × N. excelsior PI 555685 | 20 | 0 (0%) | – | – |
| N. excelsior JT × N. benthamiana JT | 18 | 1 (6%) | 32 | 28 (88%) |
| N. excelsior JT × N. benthamiana PI 555478 | 20 | 0 (0%) | – | – |
| N. excelsior JT × N. benthamiana PI 555684 | 9 | 1 (11%) | 17 | 3 (18%) |
| N. excelsior PI 224063 × N. benthamiana JT | 17 | 2 (12%) | 23 | 22 (96%) |
| N. excelsior PI 224063 × N. benthamiana PI 555478 | 17 | 2 (12%) | 20 | 11 (55%) |
| N. excelsior PI 224063 × N. benthamiana PI 555684 | 6 | 2 (33%) | 20 | 17 (85%) |
| N. excelsior PI 555685 × N. benthamiana JT | 3 | 1 (33%) | 31 | 29 (94%) |
| N. excelsior PI 555685 × N. benthamiana PI 555478 | 22 | 0 (0%) | – | – |
| N. excelsior PI 555685 × N. benthamiana PI 555684 | 2 | 1 (50%) | 34 | 32 (94%) |
1 Percentage of capsules obtained. 2 Percentage of seed germination.
Table 2.
Pollen viability of hybrids between N. benthamiana and N. excelsior.
| Cross Combination | Total no. of Pollens Observed | Percentage of Pollen Viability |
|---|---|---|
| N. benthamiana JT × N. excelsior JT | 368 | 0 |
| N. benthamiana PI 555684 × N. excelsior JT | 326 | 0 |
| N. excelsior JT × N. benthamiana JT | 348 | 0 |
| N. excelsior JT × N. benthamiana PI 555684 | 335 | 0 |
| N. excelsior PI 224063 × N. benthamiana JT | 331 | 0 |
| N. excelsior PI 224063 × N. benthamiana PI 555478 | 331 | 0 |
| N. excelsior PI 224063 × N. benthamiana PI 555684 | 360 | 0 |
| N. excelsior PI 555685 × N. benthamiana JT | 313 | 0 |
| N. excelsior PI 555685 × N. benthamiana PI 555684 | 318 | 0 |
Table 3.
Backcrosses of hybrids between N. benthamiana and N. excelsior to both parents as male parents.
Table 3.
Backcrosses of hybrids between N. benthamiana and N. excelsior to both parents as male parents.
| Cross Combination | No. of Flowers Pollinated | No. of Capsules Obtained |
|---|---|---|
| (N. benthamiana JT × N. excelsior JT) × N. benthamiana JT | 20 | 0 |
| (N. benthamiana JT × N. excelsior JT) × N. excelsior JT | 20 | 0 |
| (N. benthamiana PI 555684 × N. excelsior JT) × N. benthamiana PI 555684 | 20 | 0 |
| (N. benthamiana PI 555684 × N. excelsior JT) × N. excelsior JT | 20 | 0 |
| (N. excelsior JT × N. benthamiana JT) × N. benthamiana JT | 20 | 0 |
| (N. excelsior JT × N. benthamiana JT) × N. excelsior JT | 20 | 0 |
| (N. excelsior JT × N. benthamiana PI 555684) × N. benthamiana PI 555684 | 20 | 0 |
| (N. excelsior JT × N. benthamiana PI 555684) × N. excelsior JT | 20 | 0 |
| (N. excelsior PI 224063 × N. benthamiana JT) × N. benthamiana JT | 20 | 0 |
| (N. excelsior PI 224063 × N. benthamiana JT) × N. excelsior PI 224063 | 20 | 0 |
| (N. excelsior PI 224063 × N. benthamiana PI 555478) × N. benthamiana PI 555478 | 20 | 0 |
| (N. excelsior PI 224063 × N. benthamiana PI 555478) × N. excelsior PI 224063 | 20 | 0 |
| (N. excelsior PI 224063 × N. benthamiana PI 555684) × N. benthamiana PI 555684 | 23 | 0 |
| (N. excelsior PI 224063 × N. benthamiana PI 555684) × N. excelsior PI 224063 | 20 | 0 |
| (N. excelsior PI 555685 × N. benthamiana JT) × N. benthamiana JT | 20 | 0 |
| (N. excelsior PI 555685 × N. benthamiana JT) × N. excelsior PI 555685 | 20 | 0 |
| (N. excelsior PI 555685 × N. benthamiana PI 555684) × N. benthamiana PI 555684 | 20 | 0 |
| (N. excelsior PI 555685 × N. benthamiana PI 555684) × N. excelsior PI 555685 | 20 | 0 |
Table 4.
Backcrosses of hybrids between N. benthamiana and N. excelsior to both parents as female parents.
Table 4.
Backcrosses of hybrids between N. benthamiana and N. excelsior to both parents as female parents.
| Cross Combination | No. of Flowers Pollinated | No. of Capsules Obtained |
|---|---|---|
| N. benthamiana JT × (N. benthamiana JT × N. excelsior JT) | 20 | 0 |
| N. excelsior JT × (N. benthamiana JT × N. excelsior JT) | 14 | 0 |
| N. benthamiana JT × (N. excelsior JT × N. benthamiana JT) | 20 | 0 |
| N. excelsior JT × (N. excelsior JT × N. benthamiana JT) | 20 | 0 |
| N. benthamiana JT × (N. excelsior PI 555685 × N. benthamiana JT) | 20 | 0 |
| N. excelsior PI 555685 × (N. excelsior PI 555685 × N. benthamiana JT) | 20 | 0 |
| N. benthamiana PI 555684 × (N. excelsior PI 555685 × N. benthamiana PI 555684) | 10 | 0 |
| N. excelsior PI 555685 × (N. excelsior PI 555685 × N. benthamiana PI 555684) | 20 | 0 |
Table 5.
Metaphase I chromosome pairing in reciprocal hybrids between N. benthamiana JT and N. excelsior JT.
Table 5.
Metaphase I chromosome pairing in reciprocal hybrids between N. benthamiana JT and N. excelsior JT.
| Cross Combination | Mean Chromosome Configuration Per Cell | |||
|---|---|---|---|---|
| Univalent | Bivalent | Trivalent | Tetravalent | |
| N. benthamiana JT × N. excelsior JT | 3.50 ± 0.32 | 12.80 ± 0.25 | 2.80 ± 0.18 | 0.12 ± 0.07 |
| N. excelsior JT × N. benthamiana JT | 2.80 ± 0.21 | 14.04 ± 0.24 | 2.44 ± 0.15 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tezuka, T.; Kitamura, N.; Yanase, M.; Morikawa, T. Evaluation of Crossability between Nicotiana benthamiana and Nicotiana excelsior. Agronomy 2021, 11, 2583. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122583
AMA Style
Tezuka T, Kitamura N, Yanase M, Morikawa T. Evaluation of Crossability between Nicotiana benthamiana and Nicotiana excelsior. Agronomy. 2021; 11(12):2583. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122583
Chicago/Turabian StyleTezuka, Takahiro, Naoto Kitamura, Masanori Yanase, and Toshinobu Morikawa. 2021. "Evaluation of Crossability between Nicotiana benthamiana and Nicotiana excelsior" Agronomy 11, no. 12: 2583. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11122583
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.