
Interruption of Seed Dormancy and In Vitro Germination of the Halophile Emerging Crop Suaeda edulis (Chenopodiaceae)

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Experimental Setup
2.2. Origin of Suaeda edulis Seeds
2.3. Preliminary Experiment to Determine the Germination Potential of S. edulis
2.4. Preparation of Treatments with the Regulator Biozyme® TS
2.5. Description of the Experiment
2.6. Germination Response Variables
2.7. Experimental Design and Statistical Analysis
3. Results
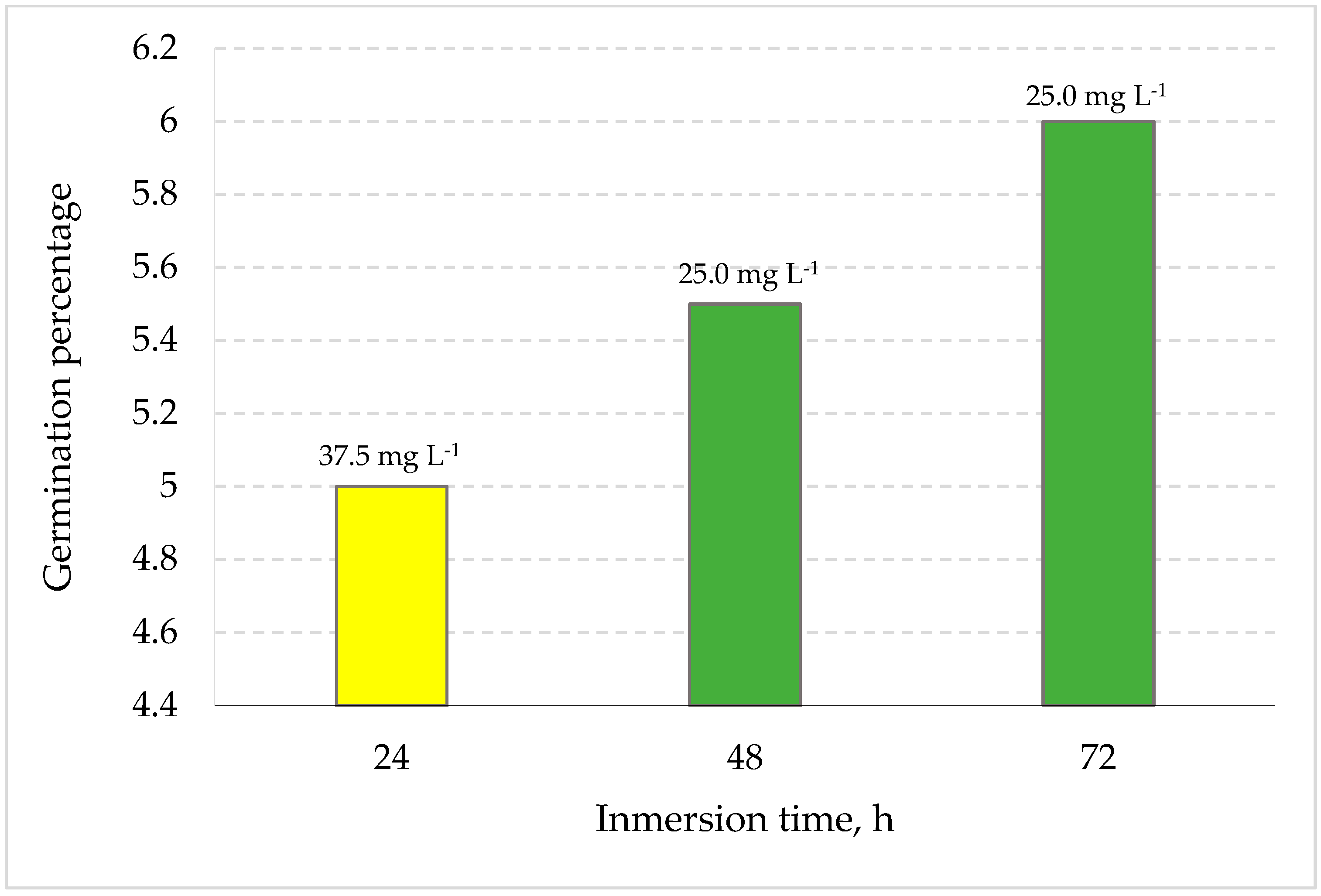
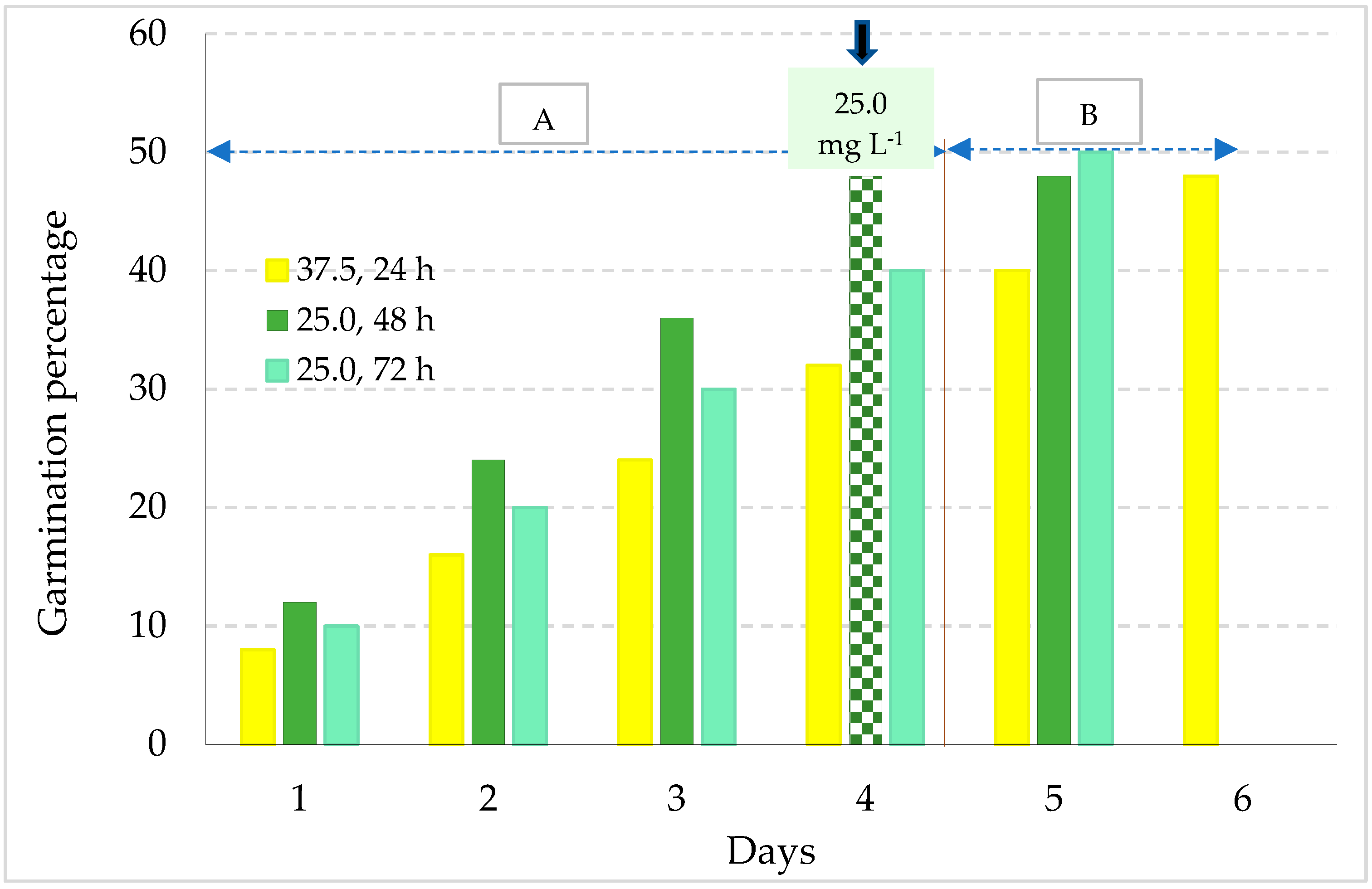
3.1. Germination Potential of S. edulis under Saline Conditions
3.2. Kolmogorov Smirnov Normality and Levene’s Homoscedasticity Tests
3.3. Effects of Treatments with the Regulator Biozyme® TS
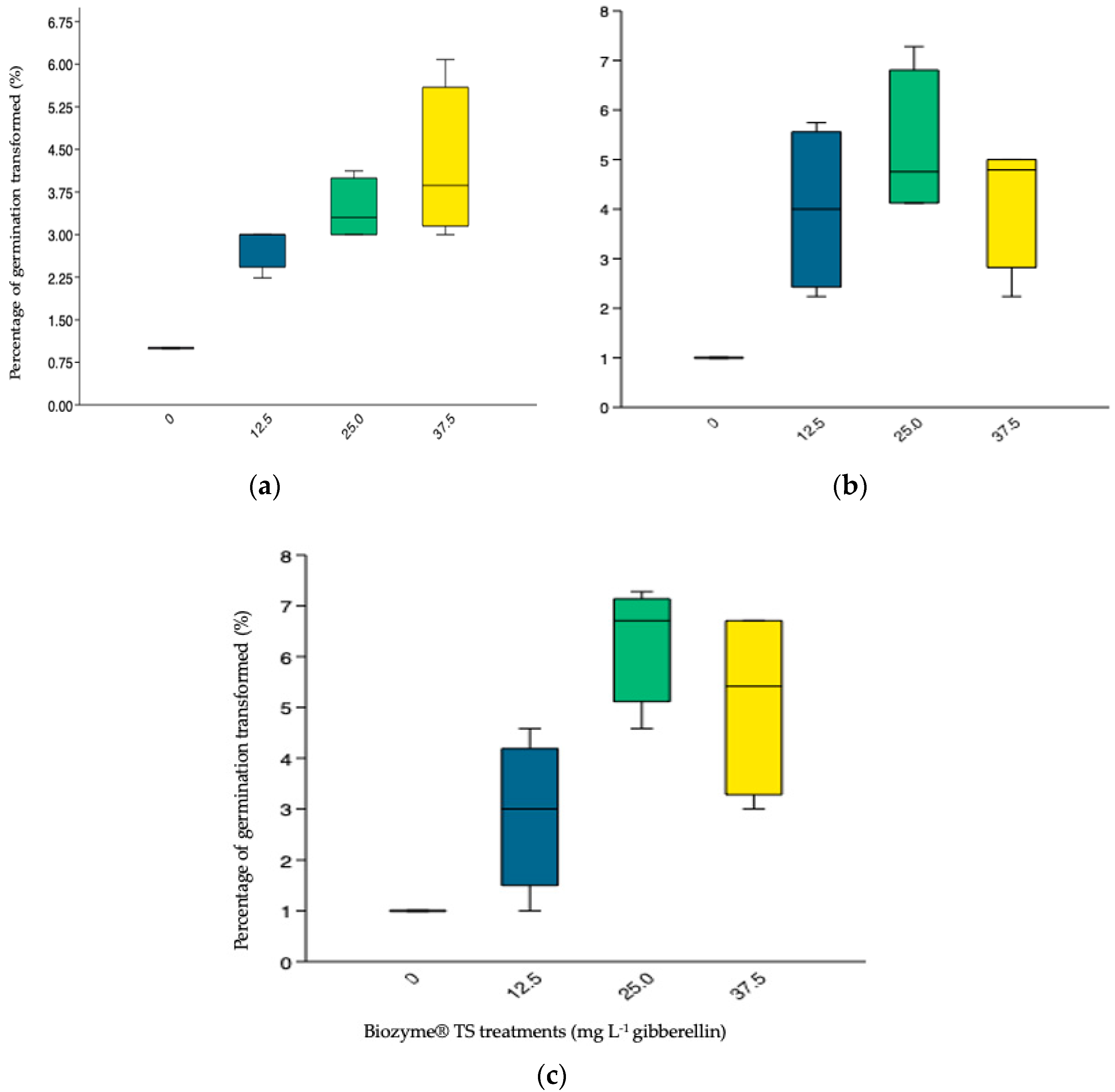
3.3.1. Germination Percentage (GP) (Transformed Values)
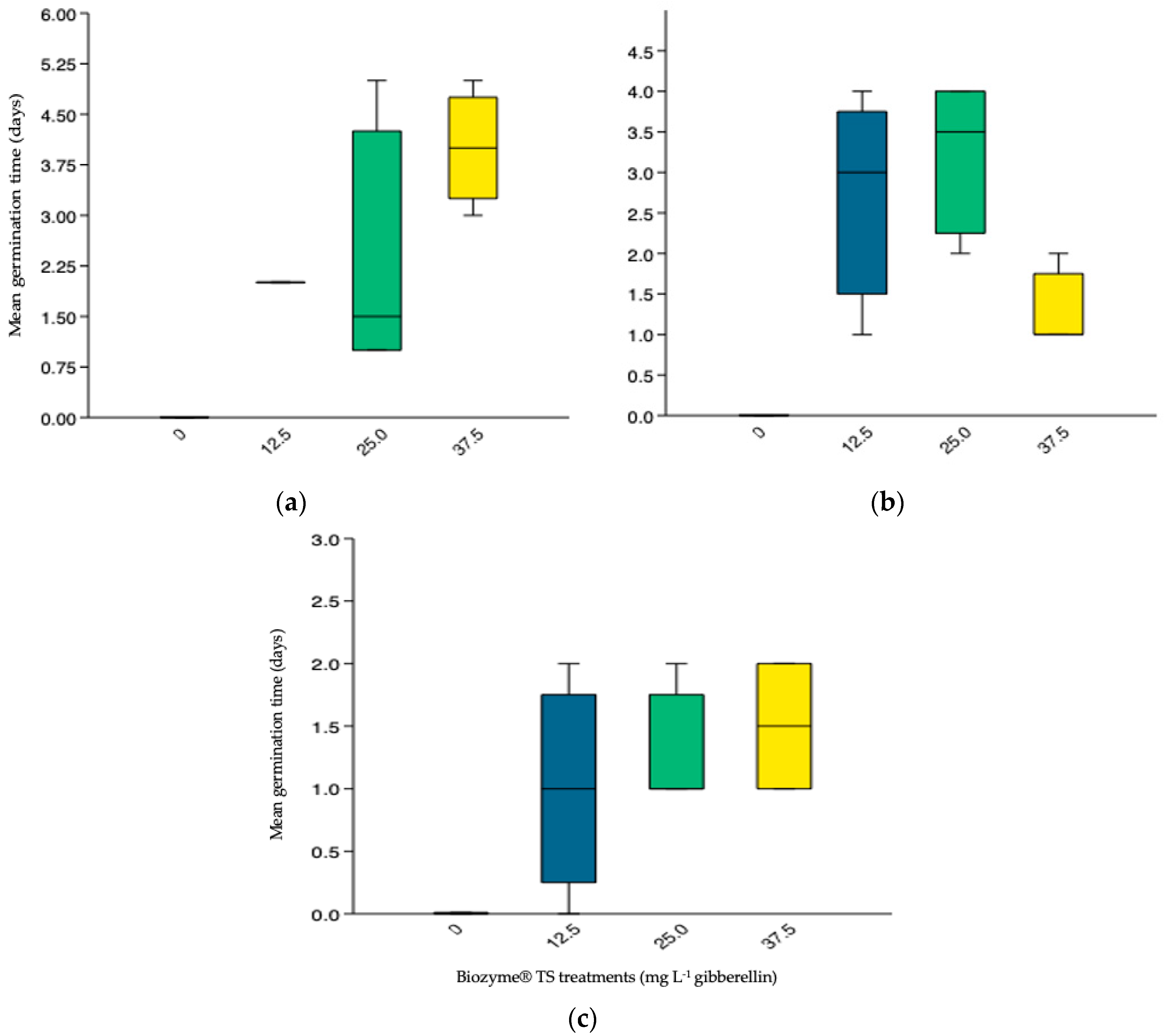
3.3.2. Mean Germination Time (MGT)
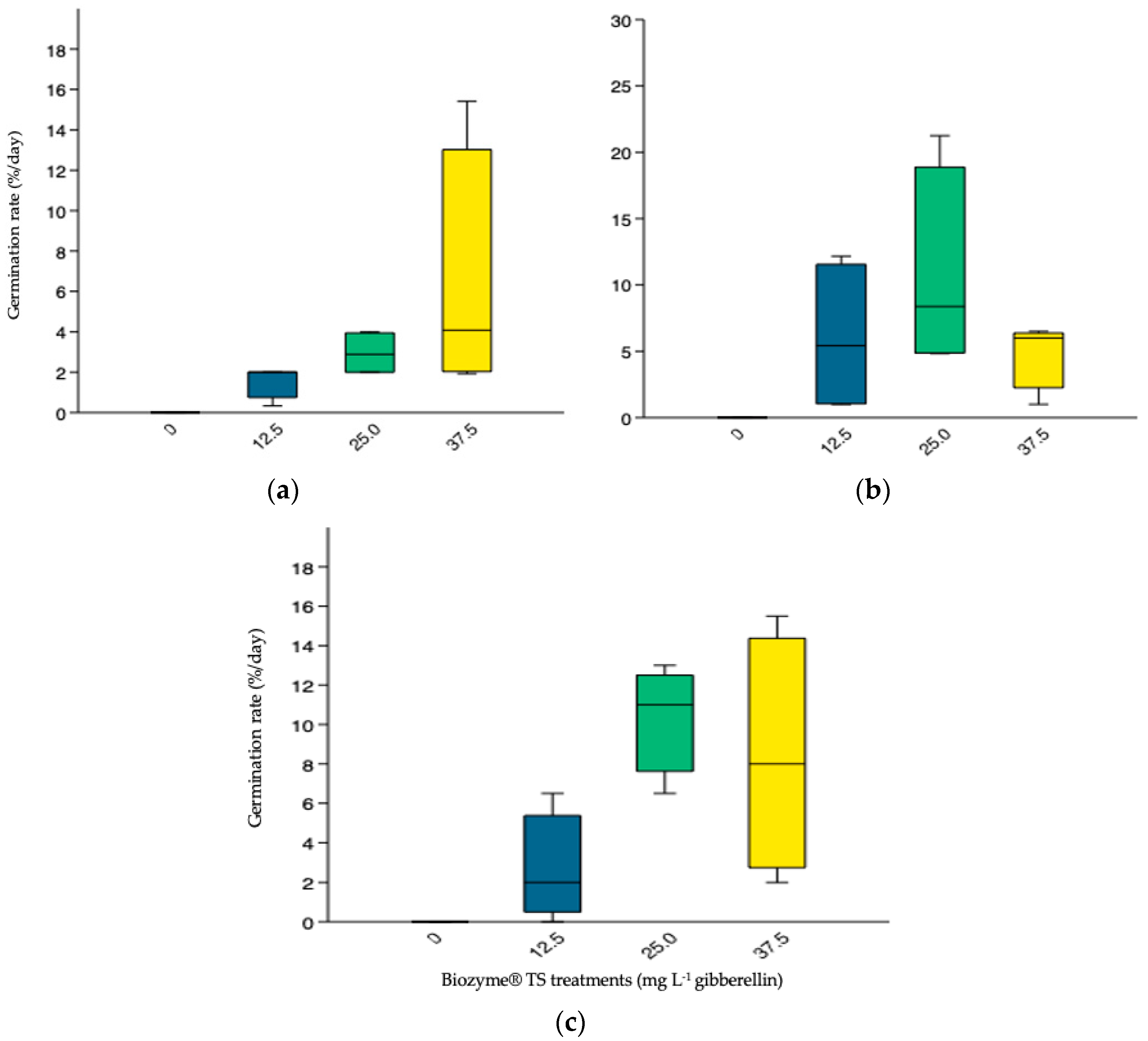
3.3.3. Germination Rate (GR)
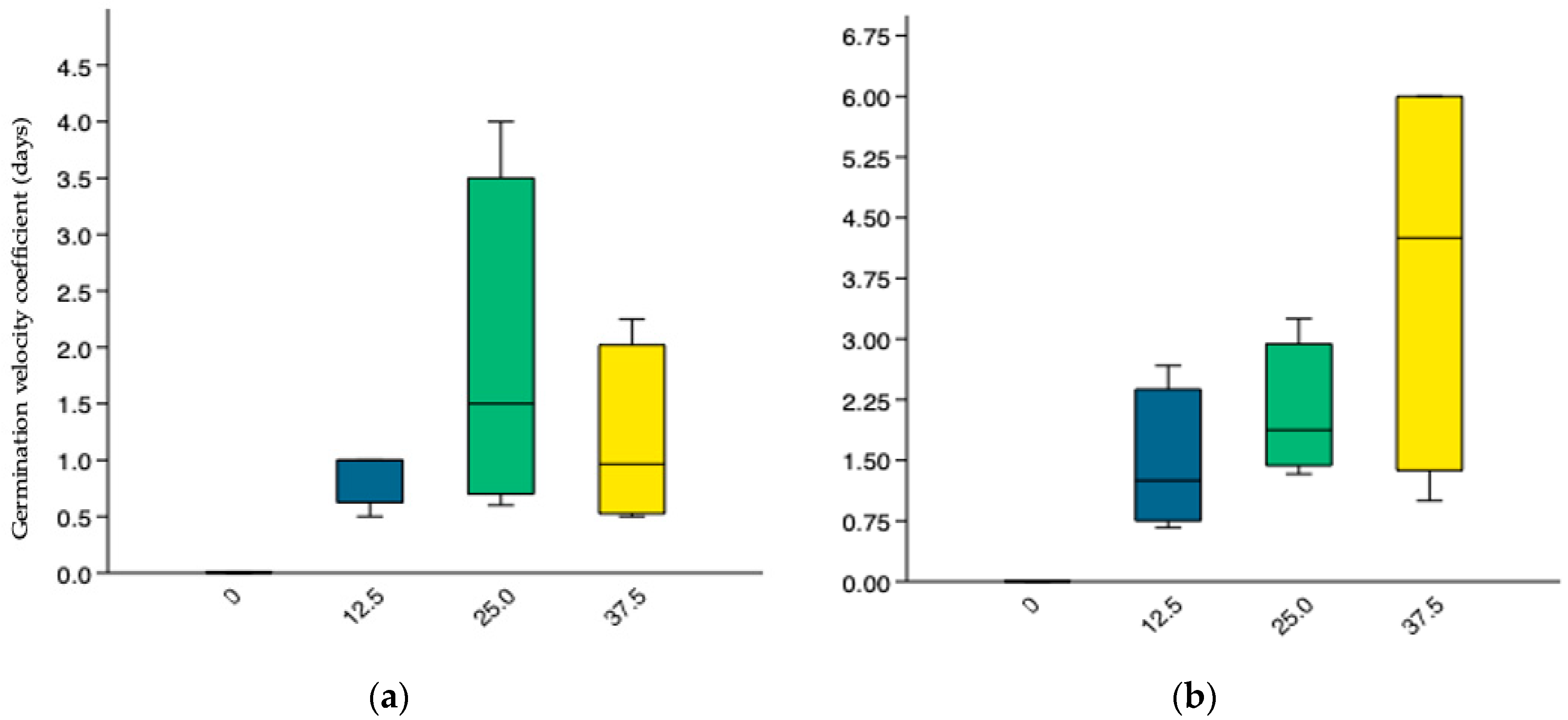
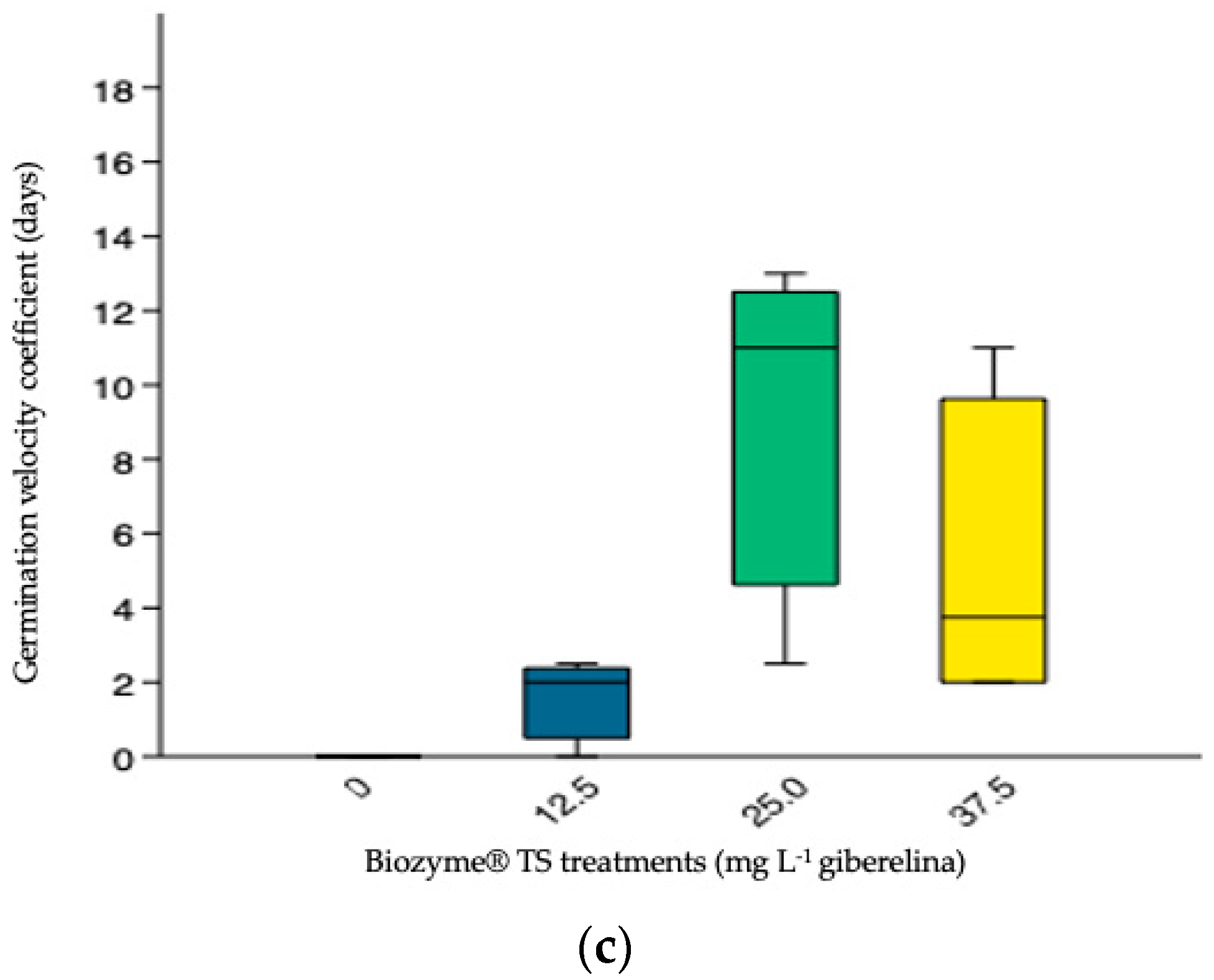
3.3.4. Germination Velocity Coefficient (GVC)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Re-Comiendo México. Available online: https://recomiendomexico.wordpress.com/2016/10/07/el-origen-de-los-romeritos/ (accessed on 3 September 2021).
- Protocolo Foreign Affairs & Lifestyle. Lista la Producción de Romeritos en San Andrés Mixquic. Available online: http://www.protocolo.com.mx/comercio-y-negocios/lista-la-produccion-de-romeritos-en-san-andres-mixquic/ (accessed on 3 September 2021).
- SIAP (Servicio de Información Agroalimentaria y Pesquera). Los Romeritos. Available online: https://www.gob.mx/siap/articulos/los-romeritos (accessed on 6 September 2021).
- Bye, R.; Linares, E. Los quelites, plantas comestibles de México: Una reflexión sobre intercambio cultural. Biodiversitas 2000, 31, 11–14. [Google Scholar]
- El Universal. ¿Sabías Que los Romeritos son un Quelite? Available online: https://www.eluniversal.com.mx/menu/sabias-que-los-romeritos-son-un-quelite (accessed on 6 September 2021).
- El poder del Consumidor. El poder de… Los Romeritos. Available online: https://elpoderdelconsumidor.org/2015/12/el-poder-de-los-romeritos/ (accessed on 7 September 2021).
- Costa-Becheleni, F.R.; Troyo-Diéguez, E.; Nieto-Garibay, A.; Bustamante-Salazar, L.A.; García-Galindo, H.S.; Murillo-Amador, B. Hydro-Environmental Criteria for Introducing an Edible Halophyte from a Rainy Region to an Arid Zone: A Study Case of Suaeda spp. as a New Crop in NW México. Plants 2021, 10, 1996. [Google Scholar] [CrossRef]
- Noguez-Hernández, R.; Carballo-Carballo, A.; Flores-Olvera, H. Suaeda edulis (Chenopodiaceae), Una nueva especies de lagos salinos del centro de México. Bot. Sci. 2013, 91, 19–25. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-42982013000100003&Ing=es&tlng= (accessed on 7 September 2021). [CrossRef] [Green Version]
- Diario Oficial de la Federación (DOF). Lista de Especies en Riesgo de la Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Available online: https://www.dof.gob.mx/nota_detalle.php?codigo=5578808&fecha=14/11/2019 (accessed on 8 September 2021).
- Carballo-Carballo, A.; Noguez-Hernández, R.; Zárate-Castrejón, J.L.; Espinosa-Cruz, R. El romerito: Una hortaliza cultivada en el Distrito Federal. Technical Report, Colegio de Postgraduados. México. 2011. Available online: https://www.worldcat.org/title/romerito-una-hortaliza-cultivada-en-el-distrito-federal/oclc/892920676 (accessed on 8 September 2021).
- Gobierno de México (Gob.mex). Romerito. Monografías. Available online: https://www.gob.mx/cms/uploads/attachment/file/96324/Romerito_monografias.pdf (accessed on 8 September 2021).
- Gold, K.; León-Lobos, P.; Way, M. Manual de Recolección de Semillas de Plantas Silvestres. 2004. Available online: Bosques.ciren.cl/handle/123456789/32083 (accessed on 9 September 2021).
- Núñez Cuerda, E. Respuestas a la Salinidad en Varias Especies Halófitas Adaptadas a Diferentes Hábitats. Grado en Biología, Universidad de Jaén. Repository of Academic Reports, University of Jaén. 2020. Available online: Tauja.ujaen.es/handle/10953.1/12300 (accessed on 9 September 2021).
- Mendoza, A.B. Ecofisiología y bioquímica del estrés en plantas. Departamento de Horticultura, Universidad Autonoma Agraria Antonio Narro, Buenavista, Saltillo, Coah. México. 2002. Available online: https://www.researchgate.net/profile/Adalberto-Benavides-Mendoza/publication/305346608_Ecofisiologia_y_bioquimica_del_estres_en_plantas/links/57893ca808ae59aa6675e204/Ecofisiologia-y-bioquimica-del-estres-en-plantas.pdf (accessed on 10 September 2021).
- Jurado, E.; Flores, J. Is seed dormancy under environmental control or bound to plant traits? J. Veg. Sci. 2005, 16, 559–564. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Physiology and Biochemistry of Seeds in Relation to Germination: Volume 2: Viability, Dormancy, and Environmental Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Available online: https://books.google.es/books?hl=es&lr=&id=91nsCAAAQBAJ&oi=fnd&pg=PA1&dq=Physiology+and+biochemistry+of+seeds+in+relation+to+germination&ots=sy3VshRCLs&sig=1nACcMkiGfliDeN_eqZkIKwFxkI#v=onepage&q=Physiology%20and%20biochemistry%20of%20seeds%20in%20relation%20to%20germination&f=false (accessed on 10 September 2021).
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular aspects of seed dormancy. Annu. Rev. Plant. Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Beevers, H. Metabolic Sinks. Agronomy & Horticulture-Faculty Publications 182. 1969. Available online: https://digitalcommons.unl.edu/cgi/viewcontent.cgi?article=1189&context=agronomyfacpub (accessed on 15 September 2021).
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytologist. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Linkies, A.; Leubner-Metzger, G. Beyond gibberellins and abscisic acid: How ethylene and jasmonates control seed germination. Plant Cell Rep. 2012, 31, 253–270. [Google Scholar] [CrossRef]
- Sánchez-Tizapantzi, G.; Ruiz-Font, A. Efecto del NaCl y de los termoperiodos sobre la germinación de semillas de Suaeda mexicana (Standl.) Standl. (Chenopodiaceae). Rev. Tecnol. Marcha. 2012, 25. [Google Scholar] [CrossRef]
- Al-Mudaris, M.A. Notes on various parameters recording the speed of seed germination. Der Trop. J. Agri. Trop. Subtrop. 1998, 99, 147–154. Available online: https://www.jarts.info/index.php/tropenlandwirt/article/view/1495 (accessed on 10 September 2021).
- Orchard, T.J. Estimating the parameters of plant seedling emergence. Seed Sci. Technol. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201302496360 (accessed on 10 September 2021).
- Kolmogorov-Smirnov Test of Normality (Social Science Statistics). Available online: https://www.socscistatistics.com/tests/kolmogorov/default.aspx (accessed on 10 September 2021).
- Levene’s Homocedasticity Test of Similarity of Variances (Statistics Kingdom). Available online: https://www.statskingdom.com/230var_levenes.html (accessed on 10 September 2021).
- PAST (The Past of the Future) Version 4.04 (mac). Available online: https://www.nhm.uio.no/english/research/infrastructure/past/ (accessed on 12 September 2021).
- Binet, P.; Boucaud, J. Dormance, levée de dormance et aptitude à germer en milieu salé dans le genre Suaeda forsk. Bull. Soc. Fr. Physiol. Vég. 1968, 14, 125–132. [Google Scholar]
- Capilupo, F.; Ungar, I.A. An ecological life history study of Suaeda depressa (Pursh) Wats. Adv. Front. Plant. Sci. 1969, 23, 137–158. Available online: https://ci.nii.ac.jp/naid/10016578686/ (accessed on 10 September 2021).
- Boucaud, J.; Ungar, I.A. The role of hormones in controlling the mechanically induced dormancy of Suaeda spp. Physiol. Plant 1973, 29, 97–102. [Google Scholar] [CrossRef]
- Wareing, P.F.; Saunders, P.F. Hormones, and dormancy. Annu. Rev. Plant. Biol. 1971, 22, 261–288. [Google Scholar] [CrossRef]
- Khan, A.A. Cytokinins: Permissive role in seed germination. Science 1971, 171, 853–859. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/1731340 (accessed on 15 October 2021). [CrossRef]
- Wei, Y.; Dong, M.; Huang, Z.Y.; Tan, D.Y. Factors influencing seed germination of Salsola affinis (Chenopodiaceae), a dominant annual halophyte inhabiting the deserts of Xinjiang, China. Flora-Morphol. Distrib. Funct. Ecol. Plants 2008, 203, 134–140. [Google Scholar] [CrossRef]
- Pujol, J.A.; Calvo, J.F.; Ramirez-Diaz, L. Recovery of germination from different osmotic conditions by four halophytes from southeastern Spain. Ann. Bot. 2000, 85, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Nikolaeva, M.G. Physiology of Deep Dormancy in Seeds. Izdatel’stvo “Nauka,” Leningrad. Translated from Russian by Z. Shapiro, National Science Foundation, Washington, D.C. 1969. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300461276 (accessed on 15 October 2021).
- Nikoaleva, M.G. Factors controlling the seed dormancy pattern. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A.A., Ed.; North-Holland: Amsterdam, The Netherlands; New York, NY, USA, 1977; pp. 51–74. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201302404007 (accessed on 15 October 2021).
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Elsevier: Amsterdam, The Netherlands, 1998; Available online: https://books.google.es/books?hl=es&lr=&id=vXfNCgAAQBAJ&oi=fnd&pg=PP1&dq=Seeds:+ecology,+biogeography,+and+evolution+of+dormancy+and+germination&ots=-rhk-hGRve&sig=CbjKNJwKk6JbaHvowEPGGCEtMjo#v=onepage&q=Seeds%3A%20ecology%2C%20biogeography%2C%20and%20evolution%20of%20dormancy%20and%20germination&f=false (accessed on 15 October 2021).
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Rosbakh, S.; Baskin, C.C.; Baskin, J.M. Nikolaeva et al.’s reference book on seed dormancy and germination. Ecology 2020, 101, e03049. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Murmu, J.; Razeq, F.M.; Santos, P.; Bourgault, R.; Molina, I.; Rowland, O. At MYB 41 activates ectopic suberin synthesis and assembly in multiple plant species and cell types. Plant J. 2014, 80, 216–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amen, R.D. A model of seed dormancy. Bot. Rev. 1968, 34, 1–31. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Rezaei, M.; Mousavi, A. A general overview on seed dormancy and methods of breaking it. Adv. Environ. Biol. 2011, 5, 3333–3337. Available online: https://link.gale.com/apps/doc/A275851029/AONE?u=anon~3817724b&sid=googleScholar&xid=530acd5e (accessed on 15 October 2021).
- Otroshy, M.; Zamani, A.; Khodambashi, M.; Ebrahimi, M.; Struik, P.C. Effect of exogenous hormones and chilling on dormancy breaking of seeds of asafoetida (Ferula assafoetida L.). J. Seed Sci. 2009, 2, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Erker, B. Seed Dormancy Mechanisms; Colorado Seed Laboratory, Department of Soil and Crop Sciences: Fort Collins, CO, USA, 2010. [Google Scholar]
- Leon, R.G.; Bassham, D.C.; Owen, M.D. Germination and proteome analyses reveal intraspecific variation in seed dormancy regulation in common waterhemp (Amaranthus tuberculatus). Weed Sci. 2006, 54, 305–315. [Google Scholar] [CrossRef]
- Geneve, R.L. Seed dormancy in commercial vegetable and flower species. Seed Technol. 1998, 236–250. Available online: https://0-www-jstor-org.brum.beds.ac.uk/stable/23433025 (accessed on 16 October 2021).
- Hameed, A.; Ahmed, M.Z.; Gulzar, S.; Gul, B.; Alam, J.; Hegazy, A.K.; Alatar, A.R.A.; Khan, M.A. Seed germination and recovery responses of Suaeda heterophylla to abiotic stresses. Pak. J. Bot. 2013, 45, 1649–1656. Available online: https://www.researchgate.net/publication/256706040_Seed_germination_and_recovery_responses_of_Suaeda_heterophylla_to_abiotic_stresses (accessed on 27 December 2021). [CrossRef]
- Tiwari, A.K.; Tiwari, T.N.; Prasad, S.R. Seed dormancy in ornamental plants: A review. Indian J. Agric. Sci. 2016, 86, 580–592. Available online: https://www.cabdirect.org/cabdirect/abstract/20163195467 (accessed on 15 October 2021).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| I. T., h. | Statistic D | p-Value | Significance |
|---|---|---|---|
| Germination Percentage (GP) (transformed values) | |||
| 24 | 0.22857 | 0.32239 ns | Does not differ from the normal distribution |
| 48 | 0.1677 | 0.69854 ns | “ |
| 72 | 0.20154 | 0.47375 ns | “ |
| Mean Germination Time (MGT) | |||
| 24 | 0.20237 | 0.46857 ns | Does not differ from the normal distribution |
| 48 | 0.21015 | 0.42179 ns | “ |
| 72 | 0.22082 | 0.36213 ns | “ |
| Germination Rate (GR) | |||
| 24 | 0.29324 | 0.10301 ns | Does not differ from the normal distribution |
| 48 | 0.20585 | 0.44733 ns | “ |
| 72 | 0.23889 | 0.27428 ns | “ |
| Germination Velocity Coefficient (GVC) | |||
| 24 | 0.2446 | 0.24996 ns | Does not differ from the normal distribution |
| 48 | 0.1574 | 0.76720 ns | “ |
| 72 | 0.31936 | 0.05965 ns | “ |
| I. T., h. | p-Value | F | Significance |
|---|---|---|---|
| Germination Percentage (GP) (transformed values) | |||
| 24 | 0.13524 | 2.2473 ns | Similarity of variances is accepted |
| 48 | 0.13077 | 2.2864 ns | “ |
| 72 | 0.0880 | 2.7612 ns | “ |
| Mean Germination Time (MGT) | |||
| 24 | 0.14799 | 2.1433 ns | Similarity of variances is accepted |
| 48 | 0.2475 | 1.5718 ns | “ |
| 72 | 0.2542 | 1.5430 ns | “ |
| Germination Rate (GR) | |||
| 24 | 0.1009 | 2.5943 ns | Similarity of variances is accepted |
| 48 | 0.0313 | 4.1436 ns | Similarity of variances is accepted |
| 72 | 0.0214 | 4.7075 * | Similarity of variance is not accepted |
| Germination Velocity Coefficient (GVC) | |||
| 24 | 0.0515 | 3.4506 ns | Similarity of variances is accepted |
| 48 | 0.0010 | 10.8974 ** | Similarity of variances is not accepted |
| 72 | 0.2685 | 1.4843 ns | Similarity of variances is accepted |
| I. T., h. | Treatm. | 0 | 12.5 | 25.0 | 37.5 |
|---|---|---|---|---|---|
| 24 | 0 | --- | 0.02226 * | 0.002878 ** | 0.0002785 *** |
| 12.5 | 4.855 | --- | 0.6484 ns | 0.08674 ns | |
| 25.0 | 6.528 | 1.673 | --- | 0.4876 ns | |
| 37.5 | 8.597 | 3.741 | 2.069 | --- | |
| 48 | 0 | --- | 0.02955 * | 0.00287 ** | 0.01978 * |
| 12.5 | 4.626 | --- | 0.5534 ns | 0.9955 ns | |
| 25.0 | 6.53 | 1.904 | --- | 0.6865 ns | |
| 37.5 | 4.95 | 0.324 | 1.58 | --- | |
| 72 | 0 | --- | 0.2357 ns | 0.0005322 *** | 0.004179 *** |
| 12.5 | 2.85 | --- | 0.01548 * | 0.1342 ns | |
| 25.0 | 7.998 | 5.148 | --- | 0.6034 ns | |
| 37.5 | 6.217 | 3.366 | 1.782 | --- |
| I. T., h. | Treatm. | 0 | 12.5 | 25.0 | 37.5 |
|---|---|---|---|---|---|
| 24 | 0 | --- | 0.07434 ns | 0.04124 * | 0.0007001 *** |
| 12.5 | 3.871 | --- | 0.9854 ns | 0.07349 ns | |
| 25.0 | 4.356 | 0.4851 | --- | 0.1298 ns | |
| 37.5 | 7.751 | 3.881 | 3.395 | --- | |
| 48 | 0 | --- | 0.002576 ** | 0.0006433 *** | 0.2006 ns |
| 12.5 | 6.621 | --- | 0.8284 ns | 0.1003 ns | |
| 25.0 | 7.827 | 1.206 | --- | 0.02313 * | |
| 37.5 | 3.003 | 3.618 | 4.824 | --- | |
| 72 | 0 | --- | 0.1074 ns | 0.03655 * | 0.01208 * |
| 12.5 | 3.56 | --- | 0.9196 ns | 0.6005 ns | |
| 25.0 | 4.454 | 0.8944 | --- | 0.9196 ns | |
| 37.5 | 5.349 | 1.789 | 0.8944 | --- |
| I. T., h. | Treat. | 0 | 12.5 | 25.0 | 37.5 |
|---|---|---|---|---|---|
| 24 | 0 | --- | 0.8973 ns | 0.5842 ns | 0.06628 ns |
| 12.5 | 0.9819 | --- | 0.9306 ns | 0.2046 ns | |
| 25.0 | 1.828 | 0.8465 | --- | 0.4608 ns | |
| 37.5 | 3.966 | 2.985 | 2.138 | --- | |
| 48 | 0 | --- | 0.3662 ns | 0.04498 * | 0.5347 ns |
| 12.5 | 2.401 | --- | 0.5615 ns | 0.9882 ns | |
| 25.0 | 4.285 | 1.884 | --- | 0.3888 ns | |
| 37.5 | 1.95 | 0.4514 | 2.335 | --- | |
| 72 | 0 | --- | 0.7354 ns | 0.007195 ** | 0.02845 * |
| 12.5 | 1.458 | --- | 0.04353 * | 0.1619 ns | |
| 25.0 | 5.77 | 4.312 | --- | 0.859 ns | |
| 37.5 | 4.657 | 3.199 | 1.113 | --- |
| I. T., h. | Treatm. | 0 | 12.5 | 25.0 | 37.5 |
|---|---|---|---|---|---|
| 24 | 0 | --- | 0.5142 ns | 0.04111 * | 0.2805 ns |
| 12.5 | 2.001 | --- | 0.3811 ns | 0.9621 ns | |
| 25.0 | 4.359 | 2.358 | --- | 0.6458 ns | |
| 37.5 | 2.68 | 0.6785 | 1.679 | --- | |
| 48 | 0 | --- | 0.4839 ns | 0.2085 ns | 0.009695 ** |
| 12.5 | 2.078 | --- | 0.9209 ns | 0.1221 ns | |
| 25.0 | 2.967 | 0.889 | --- | 0.3155 ns | |
| 37.5 | 5.527 | 3.449 | 2.56 | --- | |
| 72 | 0 | --- | 0.8897 ns | 0.006601 ** | 0.1634 ns |
| 12.5 | 1.01 | --- | 0.02295 * | 0.4444 ns | |
| 25.0 | 5.84 | 4.83 | --- | 0.2892 ns | |
| 37.5 | 3.191 | 2.181 | 2.649 | --- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Becheleni, F.R.; Troyo-Diéguez, E.; Nieto-Garibay, A.; Bustamante-Salazar, L.A.; García-Galindo, H.S.; Murillo-Amador, B. Interruption of Seed Dormancy and In Vitro Germination of the Halophile Emerging Crop Suaeda edulis (Chenopodiaceae). Agronomy 2022, 12, 103. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010103
Costa-Becheleni FR, Troyo-Diéguez E, Nieto-Garibay A, Bustamante-Salazar LA, García-Galindo HS, Murillo-Amador B. Interruption of Seed Dormancy and In Vitro Germination of the Halophile Emerging Crop Suaeda edulis (Chenopodiaceae). Agronomy. 2022; 12(1):103. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010103
Chicago/Turabian StyleCosta-Becheleni, Francyelli Regina, Enrique Troyo-Diéguez, Alejandra Nieto-Garibay, Luis Alejandro Bustamante-Salazar, Hugo Sergio García-Galindo, and Bernardo Murillo-Amador. 2022. "Interruption of Seed Dormancy and In Vitro Germination of the Halophile Emerging Crop Suaeda edulis (Chenopodiaceae)" Agronomy 12, no. 1: 103. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010103