
Screening of Wheat (Triticum aestivum L.) Genotypes for Drought Tolerance through Agronomic and Physiological Response

 , , ,
, , ,  ,
,  ,
,  ,
,  ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Analysis
2.2. Irrigation Water Analysis
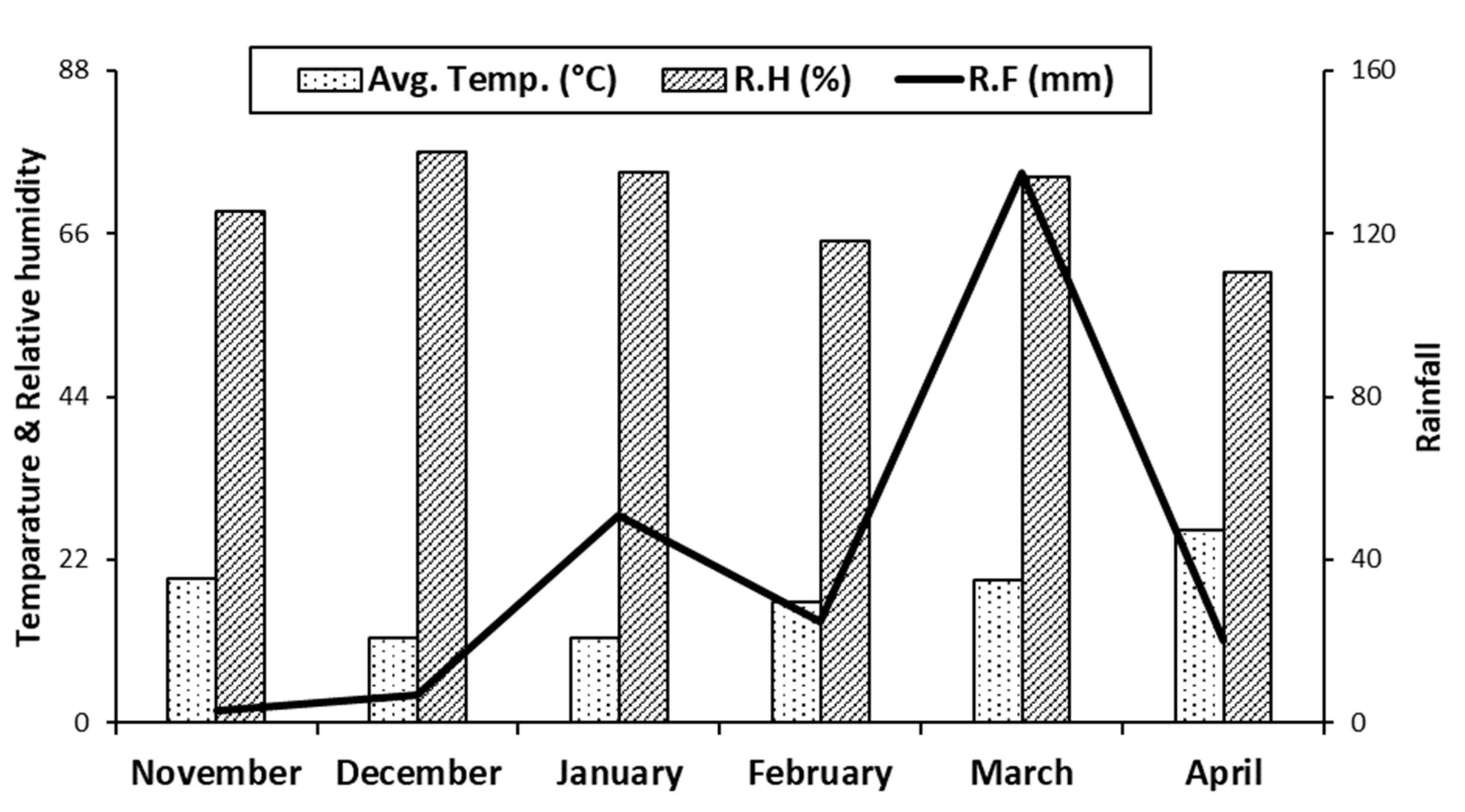
2.3. Meteorological Data
2.4. Yield and Yield-Related Parameters
2.5. Leaf Nitrogen Content
2.6. Leaf Phosphorus Content
2.7. Leaf Potassium Content
2.8. Chlorophyll Contents
2.9. Leaf Chlorophyll Contents (SPAD Value)
2.10. Relative Water Contents (RWC)
2.11. Membrane Stability Index
2.12. Statistical Analysis
3. Results
3.1. Number of Tillers per Plant and Number of Spikelets per Spike
3.2. Number of Grains per Spike
3.3. 100-Grain Weight
3.4. Seed and Biological Yield
3.5. Leaf Nitrogen Content
3.6. Leaf Phosphorus Content
3.7. Leaf Potassium Content
3.8. Chlorophyll a
3.9. Chlorophyll b
3.10. SPAD Values
3.11. Leaf Relative Water Content
3.12. Membrane Stability Index
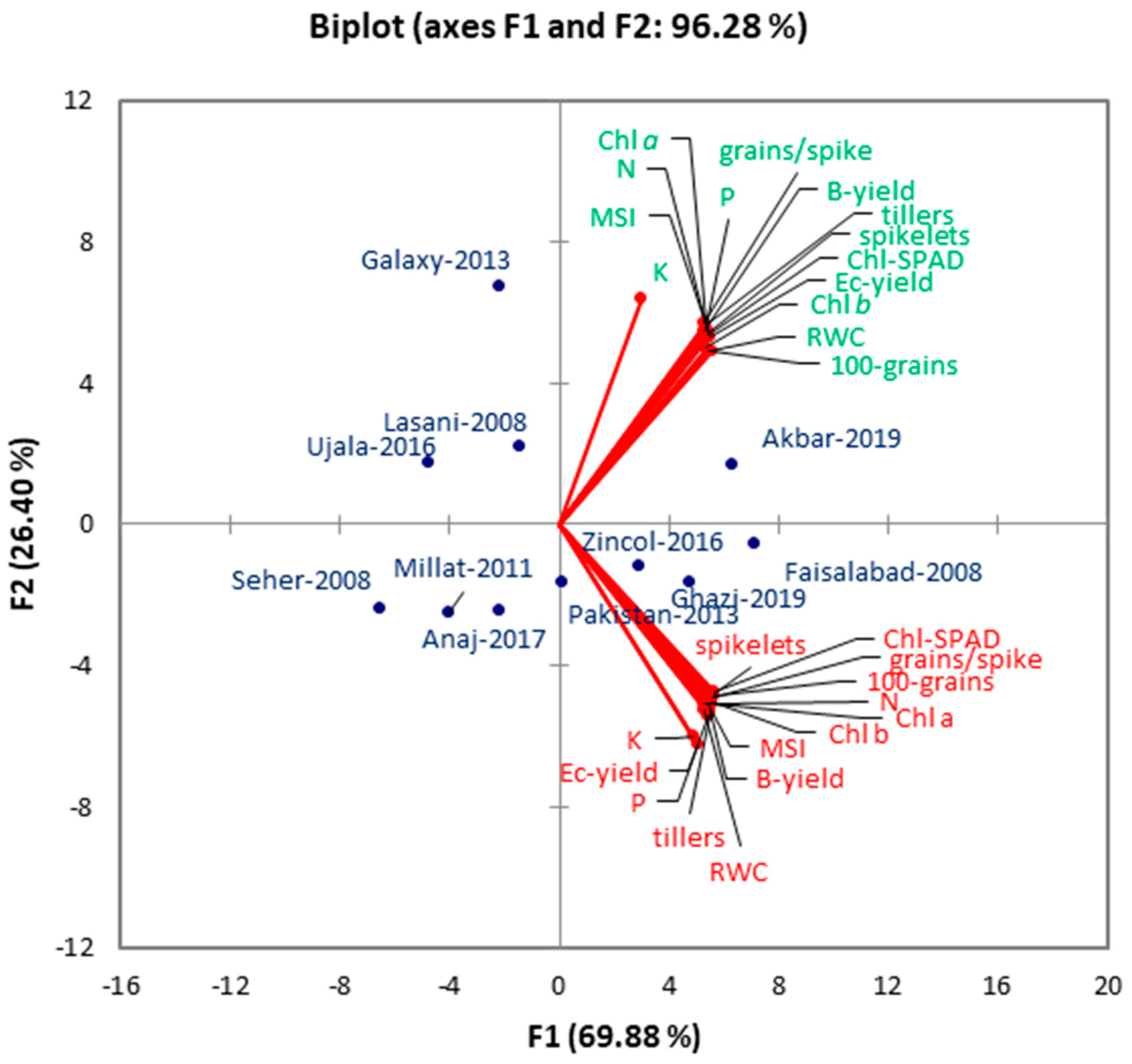
3.13. Principal Component Analysis
4. Discussion
5. Conclusions
6. Suggestions and Recommendations
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahman, M.; Soomro, U.; Haq, M.Z.-U.; Gul, S. Effects of NaCl salinity on wheat (Triticum aestivum L.) cultivars. World J. Agric. Sci. 2008, 4, 398–403. [Google Scholar]
- Abdelsalam, N.R.; Kandil, E.E.; Al-Msari, M.A.; Al-Jaddadi, M.A.; Ali, H.M.; Salem, M.Z.; Elshikh, M.S. Effect of foliar application of NPK nanoparticle fertilization on yield and genotoxicity in wheat (Triticum aestivum L.). Sci. Total Environ. 2019, 653, 1128–1139. [Google Scholar] [CrossRef]
- Datta, J.; Nag, S.; Banerjee, A.; Mondai, N. Impact of salt stress on five varieties of wheat (Triticum aestivum L.) cultivars under laboratory condition. J. Appl. Sci. Environ. Manag. 2009, 13, 93–97. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Mehmood, S.S.; Tabassum, J.; Batool, R. Targeting plant hormones to develop abiotic stress resistance in wheat. In Wheat Production in Changing Environments; Springer: Berlin/Heidelberg, Germany, 2019; pp. 557–577. [Google Scholar]
- Ken, P. Forms of micronutrient fertilizer. Sask agriculture and food, Rigas Karamanos (Westco). Agric. Agri-Food Can. 2004, 1, 1–10. [Google Scholar]
- Saddiq, M.S.; Afzal, I.; Iqbal, S.; Hafeez, M.B.; Raza, A. Low leaf sodium content improves the grain yield and physiological performance of wheat genotypes in saline-sodic soil. Pesqui. Agropecu. Trop. 2021, 51. [Google Scholar] [CrossRef]
- Wasaya, A.; Manzoor, S.; Yasir, T.A.; Sarwar, N.; Mubeen, K.; Ismail, I.A.; Raza, A.; Rehman, A.; Hossain, A.; EL Sabagh, A. Evaluation of Fourteen Bread Wheat (Triticum aestivum L.) Genotypes by Observing Gas Exchange Parameters, Relative Water and Chlorophyll Content, and Yield Attributes under Drought Stress. Sustainability 2021, 13, 4799. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Naz, M.; Hussain, S.; Javed, T.; Aslam, S.; Raza, A.; Ali, H.M.; Siddiqui, M.H.; Salem, M.Z. Exogenous salicylic acid-induced drought stress tolerance in wheat (Triticum aestivum L.) grown under hydroponic culture. PLoS ONE 2021, 16, e0260556. [Google Scholar] [CrossRef] [PubMed]
- Sammar Raza, M.A.; Saleem, M.F.; Jamil, M.; Khan, I.H. Impact of Foliar Applied Glycinebetaine on Growth and Physiology of Wheat (Triticum aestivum L.) Under Drought Conditions. Pak. J. Agric. Sci. 2014, 51, 327–334. [Google Scholar]
- Abdelsalam, N.R. Marker assisted-selection of major traits in egyptian bread wheat (Triticum aestivum L.) and wild wheat (Aegilops ventricosa tausch). Plant Cell Biotechnol. Mol. Biol. 2014, 15, 67–74. [Google Scholar]
- Salim, B.B.M.; Abou El-Yazied, A.; Salama, Y.A.M.; Raza, A.; Osman, H.S. Impact of silicon foliar application in enhancing antioxidants, growth, flowering and yield of squash plants under deficit irrigation condition. Ann. Agric. Sci. 2021, 66, 176–183. [Google Scholar] [CrossRef]
- Raza, S.; Saleem, M.; Khan, I.; Jamil, M.; Ijaz, M.; Khan, M. Evaluating the drought stress tolerance efficiency of wheat (Triticum aestivum L.) cultivars. Russ. J. Agric. Socio-Econ. Sci. 2012, 12. [Google Scholar]
- Hussain, S.; Hussain, S.; Qadir, T.; Khaliq, A.; Ashraf, U.; Parveen, A.; Saqib, M.; Rafiq, M. Drought stress in plants: An overview on implications, tolerance mechanisms and agronomic mitigation strategies. Plant Sci. Today 2019, 6, 389–402. [Google Scholar] [CrossRef]
- Mubarik, M.S.; Khan, S.H.; Sajjad, M.; Raza, A.; Hafeez, M.B.; Yasmeen, T.; Rizwan, M.; Ali, S.; Arif, M.S. A manipulative interplay between positive and negative regulators of phytohormones: A way forward for improving drought tolerance in plants. Physiol. Plant. 2021, 172, 1269–1290. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Naseer, M.A.; Hussain, S.; Nengyan, Z.; Ejaz, I.; Ahmad, S.; Farooq, M.; Xiaolong, R. Shading under drought stress during grain filling attenuates photosynthesis, grain yield and quality of winter wheat in the Loess Plateau of China. J. Agron. Crop. Sci. 2021. [Google Scholar] [CrossRef]
- Li, M.; Yang, Y.; Raza, A.; Yin, S.; Wang, H.; Zhang, Y.; Dong, J.; Wang, G.; Zhong, C.; Zhang, H. Heterologous expression of Arabidopsis thaliana rty gene in strawberry (Fragaria x ananassa Duch.) improves drought tolerance. BMC Plant Biol. 2021, 21, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.F.; Sammar Raza, M.A.; Ahmad, S.; Khan, I.H.; Shahid, A.M. Understanding and mitigating the impacts of drought stress in cotton—A review. Pak. J. Agric. Sci. 2016, 53. [Google Scholar]
- Abdelsalam, N.R.; Abdel-Megeed, A.; Ghareeb, R.Y.; Ali, H.M.; Salem, M.Z.; Akrami, M.; Al-Hayalif, M.F.; Desoky, E.S.M. Genotoxicity assessment of amino zinc nanoparticles in wheat (Triticum aestivum L.) as cytogenetical perspective. Saudi J. Biol. Sci. 2022, in press. [Google Scholar] [CrossRef]
- Chowdhury, M.K.; Hasan, M.A.; Bahadur, M.M.; Islam, M.; Hakim, M.; Iqbal, M.A.; Javed, T.; Raza, A.; Shabbir, R.; Sorour, S. Evaluation of drought tolerance of some wheat (Triticum aestivum L.) genotypes through phenology, growth, and physiological indices. Agronomy 2021, 11, 1792. [Google Scholar] [CrossRef]
- Bahar, A.A.; Faried, H.N.; Razzaq, K.; Ullah, S.; Akhtar, G.; Amin, M.; Bashir, M.; Ahmed, N.; Masoud, F.; Ahmar, S.; et al. Potassium-Induced Drought Tolerance of Potato by Improving Morpho-Physiological and Biochemical Attributes. Agronomy 2021, 11, 2573. [Google Scholar] [CrossRef]
- Islam, M.R.; Sarker, B.C.; Alam, M.A.; Javed, T.; Alam, M.J.; Zaman, M.S.U.; Azam, M.G.; Shabbir, R.; Raza, A.; Habib-ur-Rahman, M.; et al. Yield Stability and Genotype Environment Interaction of Water Deficit Stress Tolerant Mung Bean (Vigna radiata L. Wilczak) Genotypes of Bangladesh. Agronomy 2021, 11, 2136. [Google Scholar]
- Mahmood, T.; Wang, X.; Ahmar, S.; Abdullah, M.; Iqbal, M.S.; Rana, R.M.; Yasir, M.; Khalid, S.; Javed, T.; Mora-Poblete, F.; et al. Genetic Potential and Inheritance Pattern of Phenological Growth and Drought Tolerance in Cotton (Gossypium Hirsutum L.). Front. Plant Sci. 2021, 12, 705392. [Google Scholar] [PubMed]
- Zafar-ul-Hye, M.; Zahir, Z.; Shahzad, S.; Irshad, U.; Arshad, M. Isolation and screening of rhizobia for improving growth and nodulation of lentil (Lens culinaris Medic) seedlings under axenic conditions. Soil Environ. 2007, 26, 81–91. [Google Scholar]
- Keyvan, S. The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Xiuling, J.; Jiali, J.; Ruikun, M.; Jianli, L. A study on water use efficiency and its components in high-yielding winter wheat. Zuo Wu Xue Bao 1999, 25, 309–314. [Google Scholar]
- Abid, N.; Maqbool, A.; Malik, K.A. Screening commercial wheat (Triticum aestivum L.) varieties for Agrobacterium mediated transformation ability. Pak. J. Agric. Sci. 2014, 51, 83–89. [Google Scholar]
- Jones, H.D. Wheat transformation: Current technology and applications to grain development and composition. J. Cereal Sci. 2005, 41, 137–147. [Google Scholar] [CrossRef]
- Vendruscolo, E.C.G.; Schuster, I.; Negra, E.S.; Scapim, C.A. Callus induction and plant regeneration by Brazilian new elite wheat genotypes. Crop. Breed. Appl. Biotechnol. 2008, 8, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water relations in winter wheat as drought resistance indicators. Crop. Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Premachandra, G.S.; Saneoka, H.; Ogata, S. Cell membrane stability, an indicator of drought tolerance, as affected by applied nitrogen in soyabean. J. Agric. Sci. 1990, 115, 63–66. [Google Scholar] [CrossRef]
- Sairam, R. Effect of moisture-stress on physiological activities of two contrasting wheat genotypes. Indian J. Exp. Biol. 1994, 32, 594. [Google Scholar]
- Steel, R.; Torrie, J. Principle and Procedure of Statistics: A Biometrical Approach; McGraw Hill Publication Co.: New York, NY, USA, 1997; p. 633. [Google Scholar]
- Ashraf, M.; Bokhari, H.; Cristiti, S.N. Variation in osmotic adjustment of lentil (Lens culinaris Medik) in response to drought. Acta Bot. Neerl. 1992, 41, 51–62. [Google Scholar] [CrossRef]
- Tuberosa, R.; Salvi, S. Genomics-based approaches to improve drought tolerance of crops. Trends Plant Sci. 2006, 11, 405–412. [Google Scholar] [CrossRef]
- Dutta, P.; Bera, A.K. Screening of mung bean genotypes for drought tolerance. Legume Res. 2008, 31, 145–148. [Google Scholar]
- Kaur, R.; Kaur, J.; Bain, T.S. Screening of Mung bean genotypes for drought tolerance using different water potential levels. J. Adv. Agril. Technol. 2017, 4, 159–164. [Google Scholar]
- Tian, Y.; Guan, B.; Zhou, D.; Yu, J.; Li, G.; Lou, Y. Responses of seed germination, seedling growth, and seed yield traits to seed pretreatment in maize (Zea mays L.). Sci. World J. 2014, 2014, 1–8. [Google Scholar]
- Patade, V.Y.; Maya, K.; Zakwan, A. Seed priming mediated germination improvement and tolerance to subsequent exposure to cold and salt stressin capsicum. Res. J. Seed Sci. 2011, 4, 125–136. [Google Scholar]
- Mickky, B.M.; Aldesuquy, H.S. Impact of osmotic stress on seedling growth observations, membrane characteristics and antioxidant defense system of different wheat genotypes. Egypt. J. Basic Appl. Sci. 2017, 4, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A.; Lewandrowski, W.; Stevens, J.; Muñoz-Rojas, M. The limitations of seedling growth and drought tolerance to novel soil substrates in arid systems: Implications for restoration success. Geophys. Res. Abst. 2016, 18, 1–2. [Google Scholar]
- Din, A.; Ahmad, M.; Watto, F.M.; Ahmed, S.; Ali, I.; Shah, M.K.N. Drought tolerance screening in thirty common wheat (Triticum aestivum L.) genotypes. Sarhad J. Agric. 2020, 36, 168–177. [Google Scholar] [CrossRef]
- Chorfi, A.; Taıbi, K. Biochemical screening for osmotic adjustment of wheat genotypes under drought stress. Tropicult 2011, 29, 82–87. [Google Scholar]
- Razzaq, A.; Ali, Q.; Qayyum, A.; Mahmood, I.; Ahmad, M.; Rasheed, M. Physiological responses and drought resistance index of nine wheat (Triticum aestivum L.) cultivars under different moisture conditions. Pak. J. Bot. 2013, 45, 151–155. [Google Scholar]
- Maqbool, M.M.; Ali, A.; Haq, T.; Majeed, M.; Lee, D. Response of spring wheat (Triticum aestivum L.) to induced water stress at critical growth stages. Sarhad J. Agric. 2015, 31, 53–58. [Google Scholar]
- Naeem, M.; Ahmad, M.; Shah, M.; Kamran, M.; Iqbal, M. Character association and path analysis of osmotic adjustment, growth and physiological traits in wheat. J. Anim. Plant Sci. 2016, 26, 680–685. [Google Scholar]
- Sayar, R.; Khemira, H.; Kameli, A.; Mosbahi, M. Physiological tests as predictive appreciation for drought tolerance in durum wheat (Triticum durum Desf.). Agron. Res. 2008, 6, 79–90. [Google Scholar]
- Sultan, M.A.R.F.; Hui, L.; Yang, L.J.; Xian, Z.H. Assessment of drought tolerance of some Triticum L. species through physiological indices. Czech J. Genet. Plant Breed. 2012, 48, 178–184. [Google Scholar] [CrossRef]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013, 2013, 610721. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Abbas, A.; Awan, S.; Jabran, K.; Gardezi, S. Correlated response of various morpho-physiological characters with grain yield in sorghum landraces at different growth phases. J. Anim. Plant Sci. 2011, 21, 671–679. [Google Scholar]
- Ahmad, A.; Ilyas, M.Z.; Aslam, Z.; Roman, M.; Ali, A.; Naeem, S.; Nazar, M.; ur Rehman, S. Physiological screening of cotton (Gossypium hirsutum L.) genotypes against drought tolerance. Pure Appl. Biol. (PAB) 2020, 9, 140–147. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Bellitürk, K.; Ibrahim, M.U.; Nadeem, M.; Yilmaz, K.; Ali, U.; Akbar, G. Morphological study of cotton (Gossypium hirsutum L.) varieties against drought tolerance under greenhouse conditions. Pure Appl. Biol. (PAB) 2020, 9, 696–703. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Ahmadizadeh, M. Physiological and agro-morphological response to drought stress. Middle-East J. Sci. Res. 2013, 13, 998–1009. [Google Scholar]
- Brown, S.B.; Houghton, J.D.; Hendry, G.A.F. Chlorophyll breakdown. In Chlorophyllus; Scheer, H., Ed.; CRC Press: Boca Raton, FL, USA, 1991; p. 465. [Google Scholar]
- Lv, S.; Feng, K.; Peng, S.; Wang, J.; Zhang, Y.; Bian, J.; Nie, X. Comparative analysis of the transcriptional response of tolerant and sensitive wheat genotypes to drought stress in field conditions. Agronomy 2018, 8, 247. [Google Scholar] [CrossRef] [Green Version]
- Farshadfar, E.; Ghasemi, M.; Rafii, F. Evaluation of physiological parameters as a screening technique for drought tolerance in bread wheat. J. Biodiv. Environ. Sci. 2014, 4, 175–186. [Google Scholar]
- Sikuku, P.A.; Netondo, G.W.; Onyango, J.C.; Musyimi, D.M. Chlorophyll fluorescence, protein and chlorophyll content of three nerica rainfed rice varieties under varying irrigation regimes. J. Agric. Biol. Sci. 2010, 5, 19–25. [Google Scholar]
- Rana, M.S.; Hasan, M.A.; Bahadur, M.M.; Islam, M.R. Physiological evaluation of wheat genotypes for tolerance to water deficit stress. Bangladesh Agron. J. 2017, 20, 37–52. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulias, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progres-sive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.; De Longa, J.; Ladab, R.; Prangea, R. The relationship between water status and chlorophyll a fluorescence in grapes (Vitis spp.). Postharvest Biol. Technol. 2009, 51, 193–199. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Energy dissipation in C3 plants under drought. Funct. Plant Biol. 2002, 29, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Herzog, H. Source and Sink during Reproductive Period of Wheat; Scientific Publishers: Berlin/Hamburg, Germany, 1986; pp. 147–148. [Google Scholar]
- Hosseinzadeh, S.R.; Amiri, H.; Ismaili, A. Evaluation of photosynthesis, physiological, and biochemical responses of chick-pea (Cicer arietinum L. cv. Pirouz) under water deficit stress and use of vermicompost fertilizer. J. Integr. Agric. 2018, 17, 2426–2437. [Google Scholar] [CrossRef] [Green Version]
- Tang, A.C.; Kawamitsa, Y.; Kanechi, M.; Boyer, J.S. Photosynthesis at low water potentials in leaf discs lacking epidermis. Ann. Bot. 2002, 89, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serragoa, R.A.; Alzuetaa, I.; Savinb, R.; Slafer, G.A. Understanding grain yield responses to source-sink ratios during grain filling in wheat and barley under contrasting environments. Field Crop. Res. 2013, 150, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Maralian, F.; Ebadi, A.; Didar, T.R.; Haji-Eghrari, B. Influence of water stress on wheat grain yield and proline accumulation rate. Afr. J. Agril. Res. 2010, 5, 286–289. [Google Scholar] [CrossRef]
- Li, Y.P.; Ye, W.; Wang, M.; Yan, X.D. Climate change and drought: A risk assessment of crop yield impacts. Clim. Res. 2009, 39, 31–46. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Mona, F.A.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youldash, K.M.; Barutcular, C.; EL Sabagh, A.; Toptas, I.; Kayaalp, G.T.; Hossain, A.; Alharby, H.; Bamagoos, A.; Saneoka, H.; Farooq, M. Evaluation of grain yield in fifty-eight spring bread wheat genotypes grown under heat stress. Pak. J. Bot. 2020, 52, 33–42. [Google Scholar] [CrossRef]
- Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.J. Transcription factors in plant stress responses: Challenges and potential for sugarcane improvement. Plants 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, R.; Javed, T.; Afzal, I.; Sabagh, A.E.; Ali, A.; Vicente, O.; Chen, P. Modern Biotechnologies: Innovative and Sustainable Approaches for the Improvement of Sugarcane Tolerance to Environmental Stresses. Agronomy 2021, 11, 1042. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameters | Values |
|---|---|
| pH | 8.2 |
| EC | 1.40 (dSm−1) |
| Organic Matter | 1.08 (%) |
| Nitrogen | 0.042 (%) |
| Phosphorus | 4 (ppm) |
| Potassium | 320 (ppm) |
| Textural class | Loam |
| Parameters | Values (Units) |
|---|---|
| Extra Sodium Bicarbonate (RSC) | Not found |
| Sodium Adsorption Ratio (SAR) | 0.64 (mmolc L−1) |
| Sulphate (SO4)−2 | Not found |
| Chloride (Cl)−1 | 0.81 (mmolc L−1) |
| Bicarbonate (HCo)−2 | 7.01 (mmolc L−1) |
| Carbonate (Co)−2 | Not found |
| Sodium (Na)+1 | 1.24 (mmolc L−1) |
| Calcium+Magnesium (Ca+Mg)+2 | 7.13 (mmolc L−1) |
| Electrical conductivity (ECx10) | 843 (mmolc L−1) |
| Wheat Varieties | Number of Tillers per Plant | Number of Spikelets per Spike | Number of Grains per Spike | 100-Grain Weight (g) | Seed Yield (g plant−1) | Biological Yield (g plant−1) | Leaf Nitrogen Contents (mg g−1 DW) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | |
| Akbar-2019 | 6.11 a | 4.25 fgh | 26.00 a | 14.33 ghi | 50.66 a | 33.66 kl | 5.83 a | 2.88 gh | 6.21 a | 3.18 ijk | 17.83 a | 9.01 ijk | 21.16 a | 12.20 hij |
| Ghazi-2019 | 5.91 abc | 4.31 fg | 24.58 abc | 14.83 gh | 46.33 d | 34.33 k | 5.200 c | 2.96 g | 5.84 bc | 3.26 ij | 16.70 cd | 9.23 ij | 19.33 bcd | 12.91 hi |
| Anaj-2017 | 5.66 cde | 4.05 fghij | 22.05 ef | 13.16 hij | 41.00 h | 31.00 no | 4.38 de | 2.53 ijk | 5.16 fg | 2.92 klm | 15.83 fg | 8.53 klm | 17.63 efg | 11.70 ijk |
| Ujala-2016 | 5.69 bcde | 3.79 jk | 22.50 def | 11.88 jk | 43.00 g | 27.16 qr | 4.500 de | 2.03 lm | 5.28 ef | 2.28 o | 16.08 ef | 7.8 no | 18.05 defg | 11.00 jk |
| Zincol-2016 | 5.87 abc | 4.20 fghi | 24.16 abcd | 14.00 ghi | 45.16 de | 32.500 lm | 5.03 c | 2.78 ghi | 5.76 cd | 3.09 jkl | 16.56 de | 8.95 jk | 18.97 bcde | 12.98 hi |
| Galaxy-2013 | 6.00 ab | 3.68 k | 25.08 ab | 11.16 k | 48.00 c | 26.00 r | 5.33 bc | 1.93 m | 5.98 abc | 2.11 o | 17.13 bc | 7.43 o | 19.93 abc | 10.33 k |
| Pakistan-2013 | 5.71 bcd | 4.13 fghi | 22.91 cdef | 13.55 ghij | 43.83 fg | 31.83 mn | 4.60 d | 2.61 hij | 5.50 de | 3.02 jkl | 16.21 def | 8.67 kl | 18.41 cdefg | 11.86 jk |
| Millat-2011 | 5.52 de | 3.98 ghijk | 21.66 ef | 12.91 hijk | 40.00 h | 30.16 o | 4.200 ef | 2.40 jk | 5.00 gh | 2.84l mn | 15.45 g | 8.30l mn | 17.18 fg | 11.46 ijk |
| Lasani-2008 | 5.82 abcd | 3.95 hijk | 23.50 bcde | 12.66 jk | 44.50 ef | 28.66 p | 4.68 d | 2.33 jkl | 5.55 d | 2.70 mn | 16.36 def | 8.01 mn | 18.73 cdef | 11.26 jk |
| Faisalabad-2008 | 6.06 a | 4.35 f | 25.50 a | 15.33 g | 49.33 b | 35.66 j | 5.61 ab | 3.06 g | 4.88 h | 3.39 i | 17.46 ab | 9.50 i | 20.60 ab | 13.50 h |
| Seher-2008 | 5.36 e | 3.90 ijk | 21.00 f | 12.00 jk | 38.66 i | 27.83 pq | 4.01 f | 2.2000 klm | 2.61 n | 2.61 n | 14.66 h | 7.91 no | 16.82 gs | 11.150 jk |
| Drought (WS) | 49.93 ** | 1744.82 ** | 3063.96 ** | 89.71 ** | 121.50 ** | 1029.92 ** | 797.47 ** | |||||||
| Variety (V) | 0.21 ** | 9.90 ** | 51.07 ** | 1.10 ** | 0.73 ** | 2.68 ** | 5.96 ** | |||||||
| WS × V | 0.086 * | 3.40 * | 19.55 ** | 0.37 ** | 0.35 ** | 1.06 ** | 2.53 * | |||||||
| LSD ≤ 0.05 | 4.12 | 6.58 | 2.10 | 5.61 | 3.80 | 2.62 | 6.43 | |||||||
| Wheat Genotypes | Leaf Phosphorus Content (mg g−1 DW) | Leaf Potassium Content (mg g−1 DW) | Chlorophyll a (mg g−1 FW) | Chlorophyll b (mg g−1 FW) | Chlorophyll (Spade Value) | Leaf Relative Water Content (%) | Membrane Stability Index (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | WW | WS | |
| Akbar-2019 | 6.26 a | 3.73 jk | 6.52 ab | 3.69 d | 2.81 a | 1.34 hi | 0.96 ab | 0.36 ijk | 56.00 a | 34.00 kl | 91.33 a | 73.00 hi | 85.66 a | 70.00 ij |
| Ghazi-2019 | 5.98 bc | 3.91 ij | 5.95 bc | 3.74 d | 2.65 bc | 1.38 hi | 0.97 a | 0.38 ij | 52.66 cd | 35.66 jk | 87.66 bc | 74.00 h | 81.33 bc | 71.00 hi |
| Anaj-2017 | 5.28 fg | 3.41l m | 5.57 bc | 3.41 d | 2.26 f | 1.26 ij | 0.77 f | 0.32 klm | 45.00 h | 30.33 no | 81.33 efg | 69.33 jkl | 76.33 ef | 66.00 kl |
| Ujala-2016 | 5.43 ef | 2.81 pq | 5.68 bc | 3.05 d | 2.35 ef | 1.06 kl | 0.80 ef | 0.27 no | 47.00 g | 25.66 rs | 82.66 def | 62.66 n | 77.33 def | 61.66 no |
| Zincol-2016 | 5.78 cd | 3.66 k | 5.87 bc | 3.61 d | 2.58 c | 1.32 hi | 0.88 cd | 0.35 jk | 51.33 de | 33.00 lm | 90.0 ab | 71.66 hij | 81.00 bc | 72.33 ghi |
| Galaxy-2013 | 6.10 ab | 2.63 q | 6.36 abc | 2.96 d | 2.71 ab | 1.02 l | 0.92 bc | 0.24 o | 53.33 bc | 24.33 s | 89.00 ab | 62.66 n | 83.33 ab | 59.33 o |
| Pakistan-2013 | 5.53 e | 3.56 kl | 5.75 bc | 3.50 d | 2.41 de | 1.28 ij | 0.83 de | 0.34 jkl | 49.00 f | 31.66 mn | 83.66 de | 70.33 ijk | 78.66 cde | 68.00 jk |
| Millat-2011 | 5.15 gh | 3.25 mn | 5.5 bc | 3.28 d | 2.22 fg | 1.18 jk | 0.72 g | 0.30 lmn | 43.33 h | 29.00 op | 79.66 fg | 67.66 klm | 74.66 fg | 65.00 lm |
| Lasani-2008 | 5.63 de | 3.10 no | 7.14 a | 3.20 d | 2.52 cd | 1.12 kl | 0.89 c | 0.29 mn | 50.00 ef | 27.66 pq | 85.33 cd | 66.0 lmn | 79.33 cd | 63.33 lmn |
| Faisalabad-2008 | 6.18 ab | 4.01 i | 6.45 ab | 3.78 d | 2.76 ab | 1.42 h | 0.94 ab | 0.39 i | 54.66 ab | 36.66 j | 90.33 ab | 71.66 hij | 84.66 a | 72.00 ghi |
| Seher-2008 | 5.06 h | 2.98 op | 5.39 c | 3.46 d | 2.10 g | 1.09 kl | 0.67 h | 0.28 no | 41.33 i | 26.66 qr | 78.33 g | 65.33 mn | 73.33 gh | 62.33 mn |
| Drought (WS) | 87.37 ** | 110.91 ** | 26.42 ** | 4.64 ** | 5956.50 ** | 4667.05 ** | 2853.88 ** | |||||||
| Variety (V) | 0.82 ** | 0.544 ns | 0.15 ** | 0.02 ** | 90.91 ** | 80.75 ** | 79.96 ** | |||||||
| WS × V | 0.32 ** | 0.553 ns | 0.06 ** | 0.01 ** | 28.97 ** | 29.71 ** | 29.65 ** | |||||||
| LSD ≤ 0.05 | 2.75 | 13.43 | 4.21 | 4.62 | 2.70 | 2.81 | 2.32 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, A.; Aslam, Z.; Javed, T.; Hussain, S.; Raza, A.; Shabbir, R.; Mora-Poblete, F.; Saeed, T.; Zulfiqar, F.; Ali, M.M.; et al. Screening of Wheat (Triticum aestivum L.) Genotypes for Drought Tolerance through Agronomic and Physiological Response. Agronomy 2022, 12, 287. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020287
Ahmad A, Aslam Z, Javed T, Hussain S, Raza A, Shabbir R, Mora-Poblete F, Saeed T, Zulfiqar F, Ali MM, et al. Screening of Wheat (Triticum aestivum L.) Genotypes for Drought Tolerance through Agronomic and Physiological Response. Agronomy. 2022; 12(2):287. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020287
Chicago/Turabian StyleAhmad, Ali, Zubair Aslam, Talha Javed, Sadam Hussain, Ali Raza, Rubab Shabbir, Freddy Mora-Poblete, Tasbiha Saeed, Faisal Zulfiqar, Muhammad Moaaz Ali, and et al. 2022. "Screening of Wheat (Triticum aestivum L.) Genotypes for Drought Tolerance through Agronomic and Physiological Response" Agronomy 12, no. 2: 287. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020287