
Heat Stress during Meiosis Has Lasting Impacts on Plant Growth and Reproduction in Wheat (Triticum aestivum L.)



Abstract
:1. Introduction
2. Materials and Methods
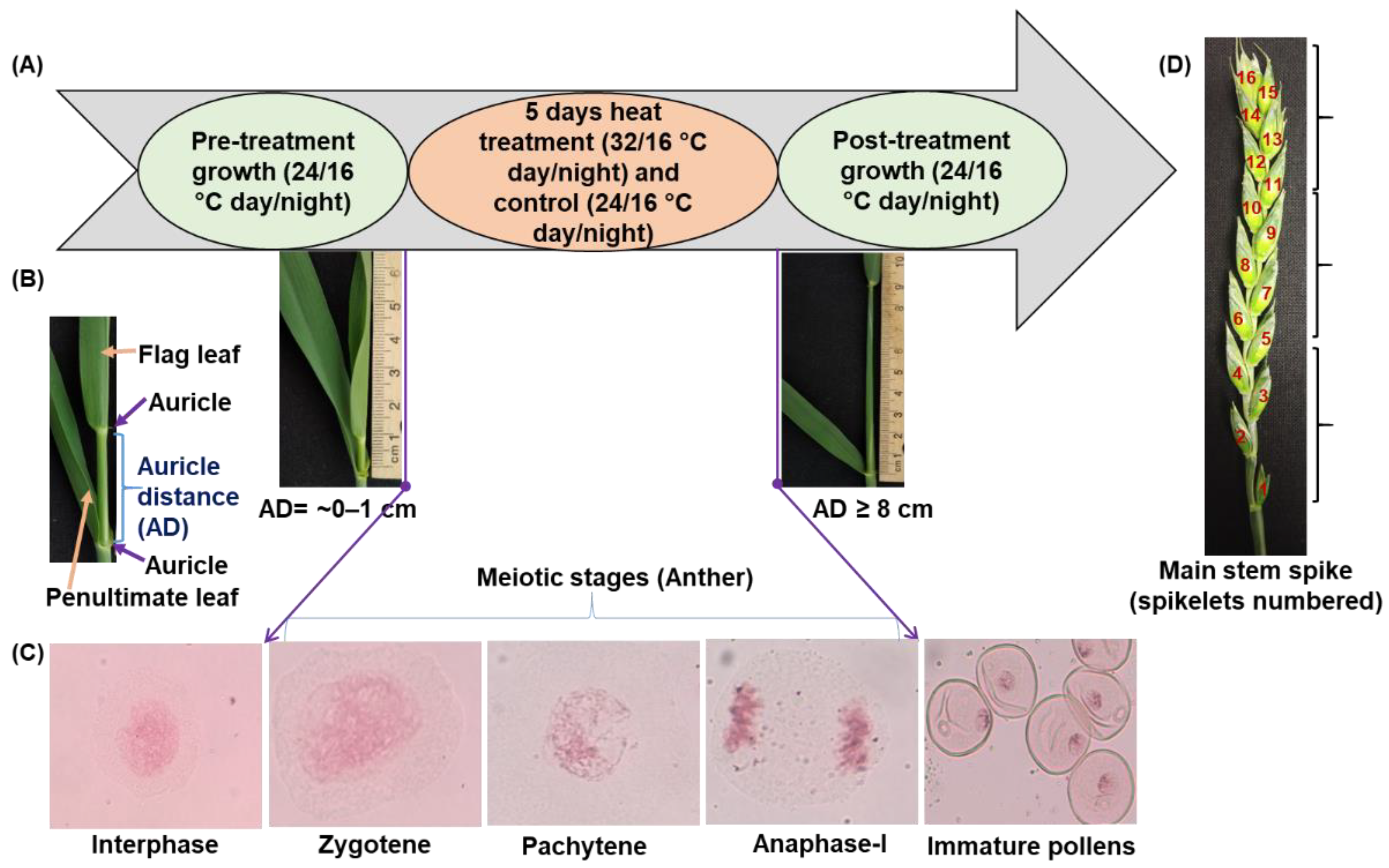
2.1. Experimental Location and Plant Material
2.2. Growth Conditions before Temperature Treatment
2.3. Auricle Distance at the Beginning of Meiosis
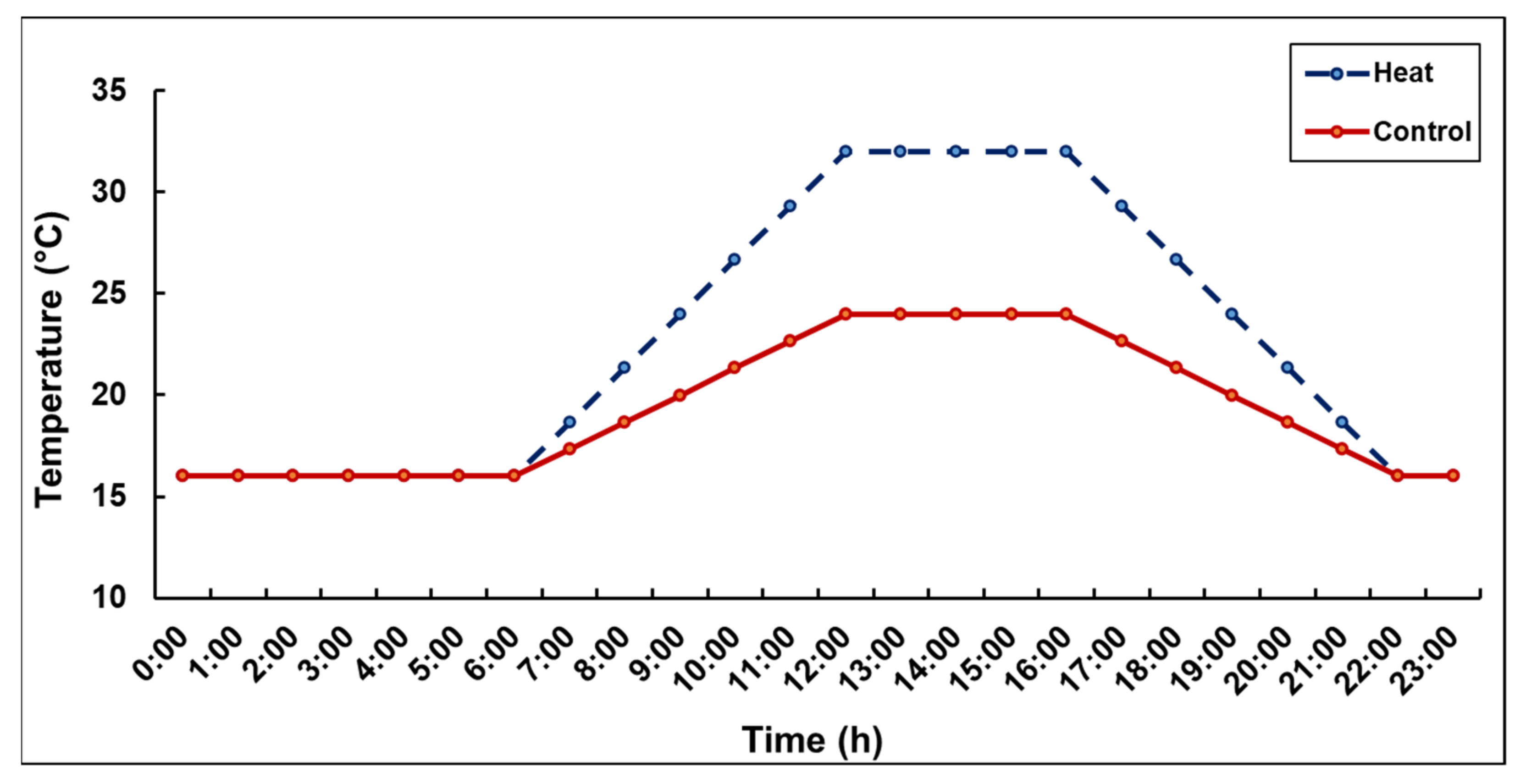
2.4. Temperature Treatments during Meiosis
2.5. Measurement of Traits during and Immediately after Temperature Treatment
2.6. Measurements of Traits at Maturity
2.7. Statistical Analyses
3. Results
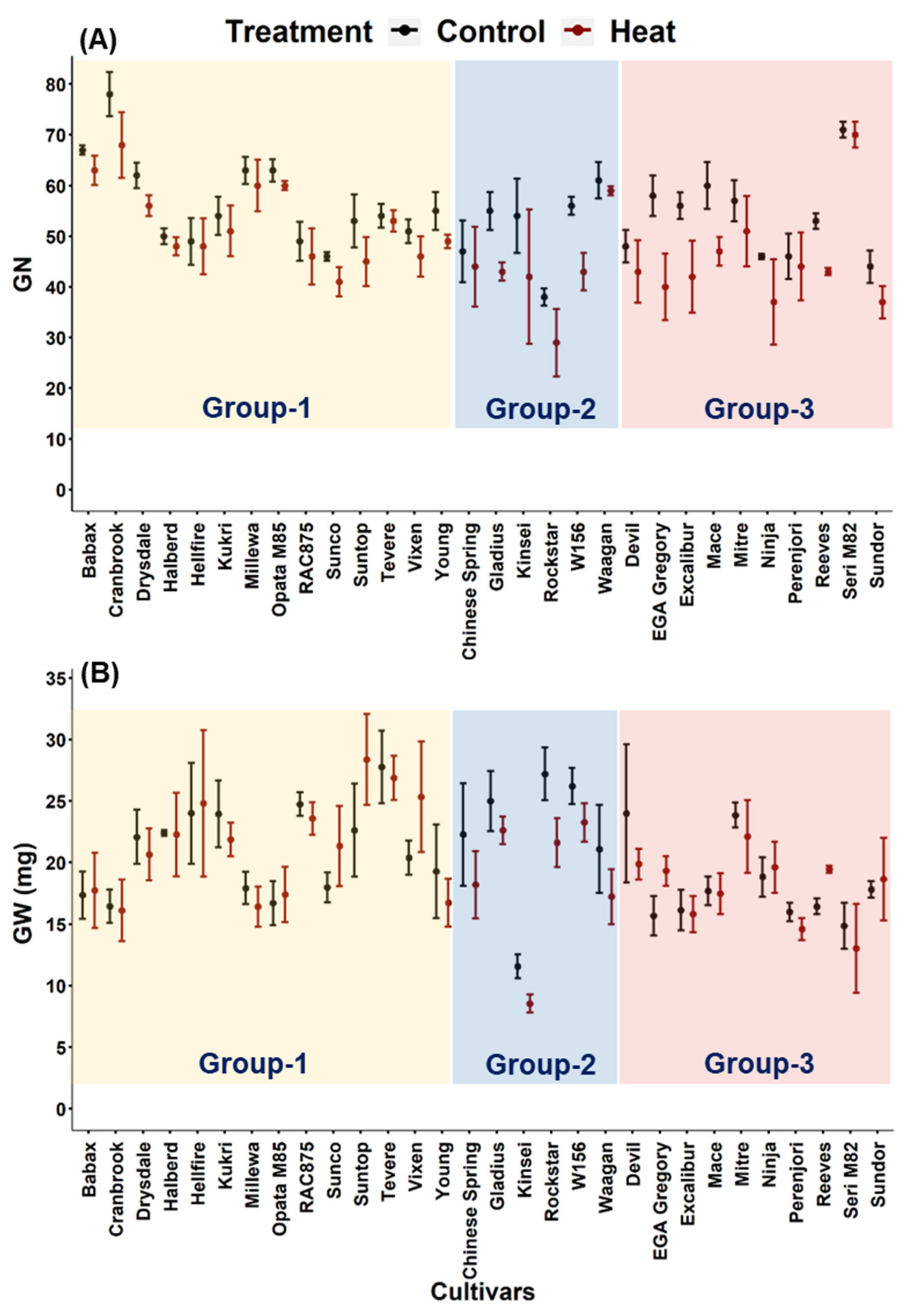
3.1. Effect of Heat Stress on Traits Measured Immediately after Temperature Treatment or at Maturity
3.2. Effect of Heat Stress on Physiological Traits Measured during the Heat Stress Treatment
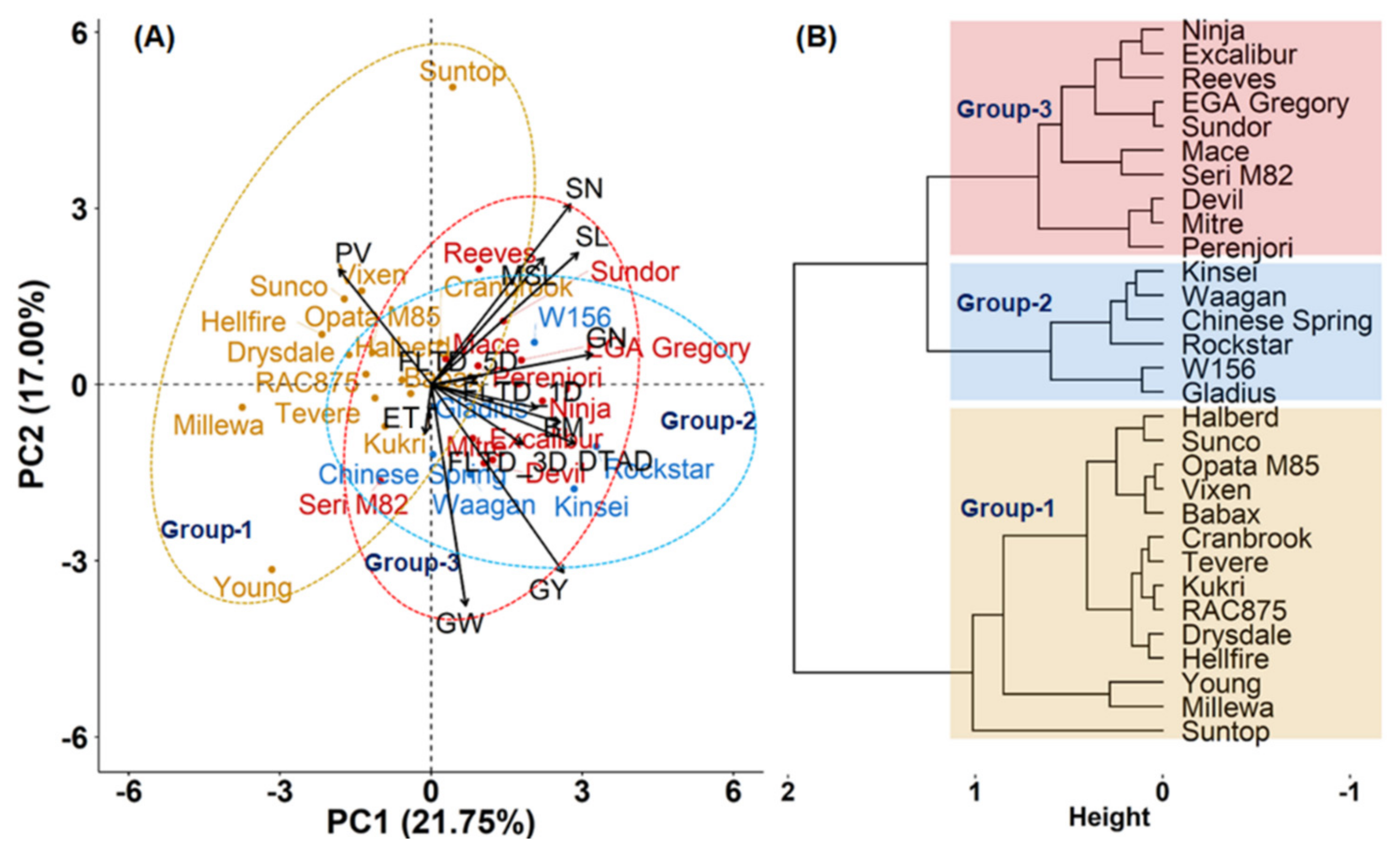
3.3. Principal Component Analysis and Clustering of Cultivars Based on Response to Heat Stress
3.4. Correlations across Cultivars for Percent Reduction in Physiological and Yield-Related Traits
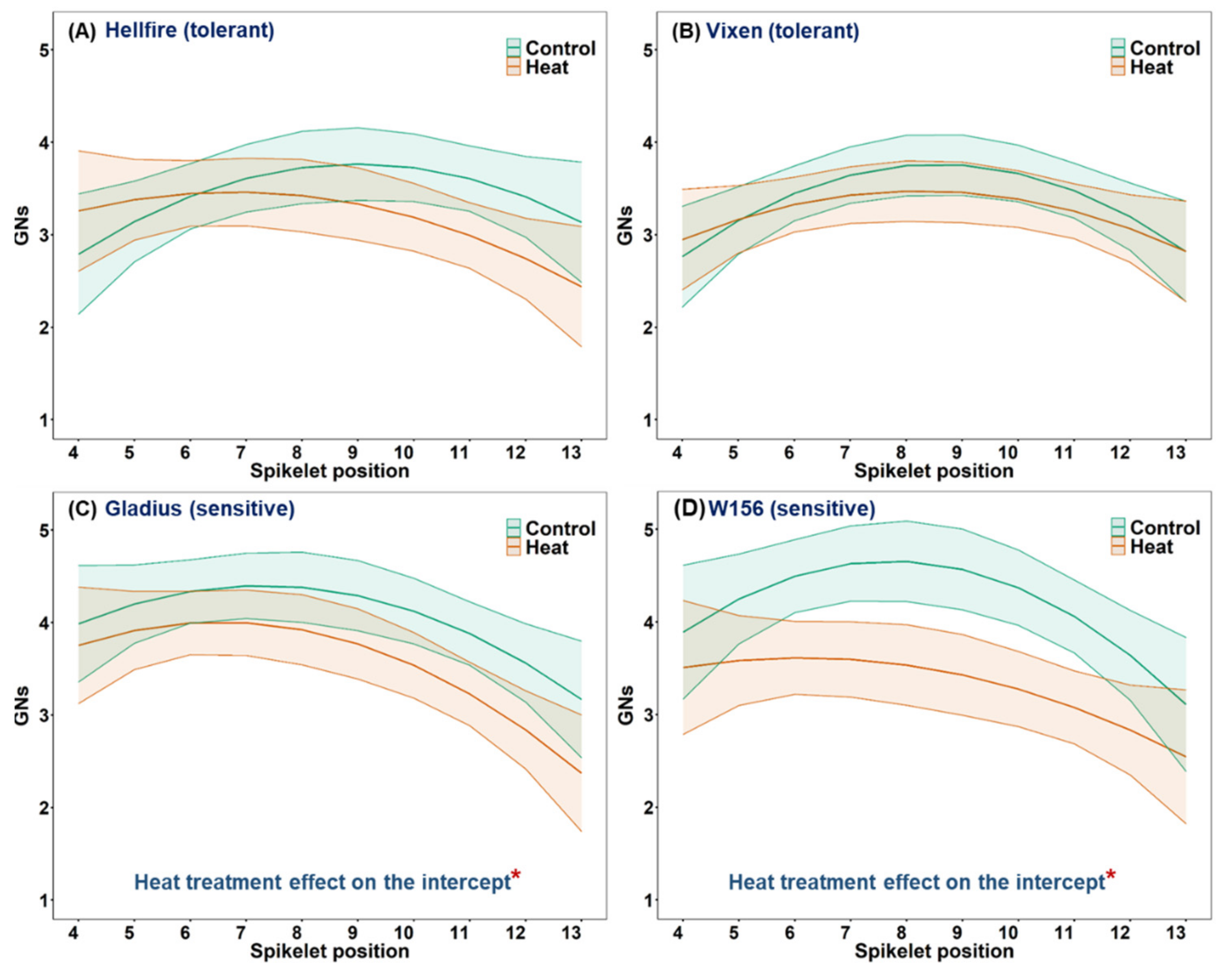
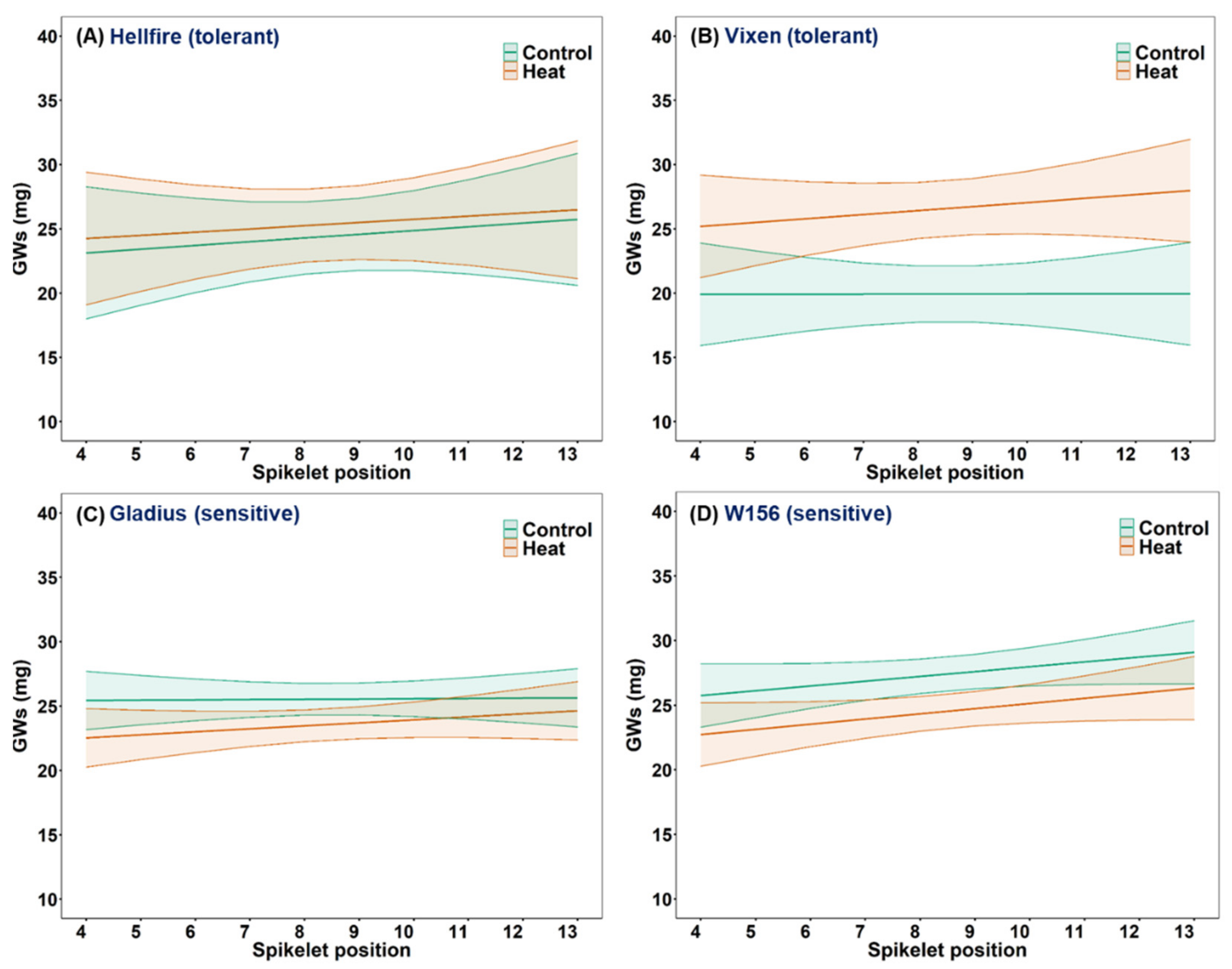
3.5. Effect of Heat Stress on Grain Number and Average Individual Grain Weight at Different Spikelet Positions on the Main Stem Spike
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AD | auricle distance |
| BM | above-ground biomass |
| CERs | controlled environment rooms |
| DTA | days from sowing to anthesis |
| DTAD | days from sowing to AD 1 cm |
| ENT | effective number of tillers |
| ET | percentage effective tillers |
| FLTD | flag leaf temperature depression |
| GN | grain number (main stem spike) |
| GNs | grain number per spikelet (main stem spike) |
| GWs | individual grain weight per spikelet (main stem spike) |
| GY | grain yield (main stem spike) |
| GW | individual grain weight (main stem spike) |
| MSL | main stem length |
| PV | pollen viability |
| SL | spike length (main stem spike) |
| SN | spikelet number (main stem spike) |
| TGY | total plant grain yield |
| TNT | total number of tillers |
References
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. FAOSTAT Database. Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 19 April 2022).
- Fischer, R.A.; Byerlee, D.; Edmeades, G. Crop Yields and Global Food Security: Will Yield Increase Continue to Feed the World? ACIAR Monograph No. 158; Australian Centre for International Agricultural Research: Canberra, Australia, 2014; p. 634. Available online: https://www.aciar.gov.au/publication/books-and-manuals/crop-yields-and-global-food-security-will-yield-increase-continue-feed-world (accessed on 10 August 2021).
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; Hanumantha, R.B.; Nair, R.M.; Prasad, P.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for policymakers 2021. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate, Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; Available online: https://www.ipcc.ch/report/ar6/wg1/#FullReport (accessed on 30 March 2022).
- Lorenz, R.; Stalhandske, Z.; Fischer, E.M. Detection of a Climate Change Signal in Extreme Heat, Heat Stress, and Cold in Europe from Observations. Geophys. Res. Lett. 2019, 46, 8363–8374. [Google Scholar] [CrossRef] [Green Version]
- NOAA. National Centers for Environmental Information. State of the Climate: Global Climate Report for Annual 2020. 2021. Available online: https://www.ncdc.noaa.gov/sotc/global/202013 (accessed on 12 September 2021).
- Australian Bureau of Agricultural and Resource Economics and Sciences. Agricultural Commodities: September Quarter 2021; Australian Bureau of Agricultural and Resource Economics and Sciences: Canberra, Australia, 2021. [Google Scholar] [CrossRef]
- Hochman, Z.; Gobbett, D.; Horan, H. Climate trends account for stalled wheat yields in Australia since 1990. Glob. Chang. Biol. 2017, 23, 2071–2081. [Google Scholar] [CrossRef]
- Warrington, I.; Dunstone, R.; Green, L. Temperature effects at three development stages on the yield of the wheat ear. Aust. J. Agric. Res. 1977, 28, 11–27. [Google Scholar] [CrossRef]
- Porter, J.R.; Gawith, M. Temperatures and the growth and development of wheat: A review. Eur. J. Agron. 1999, 10, 23–36. [Google Scholar] [CrossRef]
- Wang, E.; Martre, P.; Zhao, Z.; Ewert, F.; Maiorano, A.; Rötter, R.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. The uncertainty of crop yield projections is reduced by improved temperature response functions. Nat. Plants 2017, 3, 17102. [Google Scholar] [CrossRef] [Green Version]
- Deryng, D.; Conway, D.; Ramankutty, N.; Price, J.; Warren, R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 2014, 9, 034011. [Google Scholar] [CrossRef] [Green Version]
- Demirevska-Kepova, K.; Holzer, R.; Simova-Stoilova, L.; Feller, U. Heat stress effects on ribulose-1,5-bisphosphate carboxylase/oxygenase, Rubisco binding protein and Rubisco activase in wheat leaves. Biol. Plant. 2005, 49, 521–525. [Google Scholar] [CrossRef]
- Kadam, N.N.; Xiao, G.; Melgar, R.J.; Bahuguna, R.N.; Quinones, C.; Tamilselvan, A.; Prasad, P.V.; Jagadish, K.S. Agronomic and Physiological Responses to High Temperature, Drought, and Elevated CO2 Interactions in Cereals. Adv. Agron. 2014, 127, 111–156. [Google Scholar] [CrossRef]
- Ullah, A.; Nadeem, F.; Nawaz, A.; Siddique, K.H.M.; Farooq, M. Heat stress effects on the reproductive physiology and yield of wheat. J. Agron. Crop Sci. 2021, 208, 1–17. [Google Scholar] [CrossRef]
- Mishra, D.; Shekhar, S.; Chakraborty, S.; Chakraborty, N. High temperature stress responses and wheat: Impacts and alleviation strategies. Environ. Exp. Bot. 2021, 190, 104589. [Google Scholar] [CrossRef]
- Farooq, M.; Bramley, H.; Palta, J.A.; Siddique, K.H.M. Heat Stress in Wheat during Reproductive and Grain-Filling Phases. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- Grant, R.F.; Kimball, B.A.; Conley, M.M.; White, J.W.; Wall, G.W.; Ottman, M.J. Controlled Warming Effects on Wheat Growth and Yield: Field Measurements and Modeling. Agron. J. 2011, 103, 1742–1754. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Pal, M.; Sukumaran, S.; Parani, M.; Siddique, K.H.M. Heat stress resilient crops for future hotter environments. Plant Physiol. Rep. 2020, 25, 529–532. [Google Scholar] [CrossRef]
- Zahra, N.; Wahid, A.; Hafeez, M.B.; Ullah, A.; Siddique, K.H.; Farooq, M. Grain development in wheat under combined heat and drought stress: Plant responses and management. Environ. Exp. Bot. 2021, 188, 104517. [Google Scholar] [CrossRef]
- Viswanathan, C.; Khanna-Chopra, R. Effect of Heat Stress on Grain Growth, Starch Synthesis and Protein Synthesis in Grains of Wheat (Triticum aestivum L.) Varieties Differing in Grain Weight Stability. J. Agron. Crop Sci. 2001, 186, 1–7. [Google Scholar] [CrossRef]
- Saini, H.S.; Aspinall, D. Abnormal Sporogenesis in Wheat (Triticum aestivum L.) Induced by Short Periods of High Temperature. Ann. Bot. 1982, 49, 835–846. [Google Scholar] [CrossRef]
- Porter, J.R.; Semenov, M. Crop responses to climatic variation. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 2021–2035. [Google Scholar] [CrossRef]
- Zinn, K.E.; Tunc-Ozdemir, M.; Harper, J.F. Temperature stress and plant sexual reproduction: Uncovering the weakest links. J. Exp. Bot. 2010, 61, 1959–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolferus, R.; Ji, X.; Richards, R.A. Abiotic stress and control of grain number in cereals. Plant Sci. 2011, 181, 331–341. [Google Scholar] [CrossRef]
- Semenov, M.; Stratonovitch, P.; Alghabari, F.; Gooding, M. Adapting wheat in Europe for climate change. J. Cereal Sci. 2014, 59, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaei, E.E.; Webber, H.; Gaiser, T.; Naab, J.; Ewert, F. Heat stress in cereals: Mechanisms and modelling. Eur. J. Agron. 2015, 64, 98–113. [Google Scholar] [CrossRef]
- Bennett, M.D.; Chapman, V.; Riley, R. The duration of meiosis in pollen mother cells of wheat, rye and Triticale. Proc. R. Soc. London. Ser. B Biol. Sci. 1971, 178, 259–275. [Google Scholar] [CrossRef]
- Bennett, M.D.; Finch, R.A.; Smith, J.B.; Rao, M.K. The time and duration of female meiosis in wheat, rye and barley. Proc. R. Soc. London. Ser. B Biol. Sci. 1973, 183, 301–319. [Google Scholar] [CrossRef]
- Bennett, M.D.; Rao, M.K.; Smith, J.B.; Bayliss, M.W. Cell development in the anther, the ovule, and the young seed of Triticum aestivum L. var. Chinese Spring. Philos. Trans. R. Soc. B Biol. Sci. 1973, 266, 39–81. [Google Scholar] [CrossRef]
- Wardlaw, I.; Dawson, I.; Munibi, P. The tolerance of wheat to hight temperatures during reproductive growth. 2. Grain development. Aust. J. Agric. Res. 1989, 40, 15–24. [Google Scholar] [CrossRef]
- Prasad, P.V.; Pisipati, S.R.; Ristic, Z.; Bukovnik, U.; Fritz, A.K. Impact of Nighttime Temperature on Physiology and Growth of Spring Wheat. Crop Sci. 2008, 48, 2372–2380. [Google Scholar] [CrossRef]
- Talukder, A.; McDonald, G.K.; Gill, G.S. Effect of short-term heat stress prior to flowering and early grain set on the grain yield of wheat. Field Crops Res. 2014, 160, 54–63. [Google Scholar] [CrossRef]
- Kaur, V.; Behl, R. Grain yield in wheat as affected by short periods of high temperature, drought and their interaction during pre- and post-anthesis stages. Cereal Res. Commun. 2010, 38, 514–520. [Google Scholar] [CrossRef]
- Wheeler, T.R.; Batts, G.R.; Ellis, R.H.; Hadley, P.; Morison, J.I.L. Growth and yield of winter wheat (Triticum aestivum) crops in response to CO2and temperature. J. Agric. Sci. 1996, 127, 37–48. [Google Scholar] [CrossRef]
- Draeger, T.; Moore, G. Short periods of high temperature during meiosis prevent normal meiotic progression and reduce grain number in hexaploid wheat (Triticum aestivum L.). Theor. Appl. Genet. 2017, 130, 1785–1800. [Google Scholar] [CrossRef] [Green Version]
- Balla, K.; Karsai, I.; Bónis, P.; Kiss, T.; Berki, Z.; Horváth, Á.; Mayer, M.; Bencze, S.; Veisz, O. Heat stress responses in a large set of winter wheat cultivars (Triticum aestivum L.) depend on the timing and duration of stress. PLoS ONE 2019, 14, e0222639. [Google Scholar] [CrossRef] [PubMed]
- Dowrick, G.J. The influence of temperature on meiosis. Heredity 1957, 11, 37–49. [Google Scholar] [CrossRef]
- Rezaei, M.; Arzani, A.; Sayed-Tabatabaei, B.E. Meiotic behaviour of tetraploid wheats (Triticum turgidum L.) and their synthetic hexaploid wheat derivates influenced by meiotic restitution and heat stress. J. Genet. 2010, 89, 401–407. [Google Scholar] [CrossRef]
- Omidi, M.; Siahpoosh, M.R.; Mamghani, R.; Modarresi, M. The Influence of Terminal Heat Stress on Meiosis Abnormalities in Pollen Mother Cells of Wheat. CYTOLOGIA 2014, 79, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Saini, H.; Sedgley, M.; Aspinall, D. Effect of Heat Stress During Floral Development on Pollen Tube Growth and Ovary Anatomy in Wheat (Triticum aestivum L.). Funct. Plant Biol. 1983, 10, 137–144. [Google Scholar] [CrossRef]
- Loss, S.; Kirby, E.; Siddique, K.; Perry, M. Grain growth and development of old and modern Australian wheats. Field Crops Res. 1989, 21, 131–146. [Google Scholar] [CrossRef]
- Ji, X.; Shiran, B.; Wan, J.; Lewis, D.C.; Jenkins, C.; Condon, A.; Richards, R.A.; Dolferus, R. Importance of pre-anthesis anther sink strength for maintenance of grain number during reproductive stage water stress in wheat. Plant Cell Environ. 2010, 33, 926–942. [Google Scholar] [CrossRef]
- Barton, D.A.; Cantrill, L.C.; Law, A.M.K.; Phillips, C.G.; Sutton, B.G.; Overall, R.L. Chilling to zero degrees disrupts pollen formation but not meiotic microtubule arrays in Triticum aestivum L. Plant Cell Environ. 2014, 37, 2781–2794. [Google Scholar] [CrossRef] [PubMed]
- Onyemaobi, I.; Liu, H.; Siddique, K.H.M.; Yan, G. Both Male and Female Malfunction Contributes to Yield Reduction under Water Stress during Meiosis in Bread Wheat. Front. Plant Sci. 2017, 7, 2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.M. Possible role of abscisic acid in reducing seed set in water-stressed wheat plants. Nature 1980, 285, 655–657. [Google Scholar] [CrossRef]
- Agar, O.; Tan, D.K.; Barton, D.A.; Trethowan, R.M. Genetic variation in wheat pollen heat tolerance. In Proceedings of the 17th ASA Conference, Hobart, Australia, 20–24 September 2015; Available online: http://agronomyaustraliaproceedings.org/images/sampledata/2015_Conference/pdf/agronomy2015final00011.pdf (accessed on 5 April 2021).
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirdelmoghanloo, H.; Lohraseb, I.; Rabie, H.S.; Brien, C.; Parent, B.; Collins, N.C. Heat susceptibility of grain filling in wheat (Triticum aestivum L.) linked with rapid chlorophyll loss during a 3-day heat treatment. Acta Physiol. Plant. 2016, 38, 208. [Google Scholar] [CrossRef]
- Butler, D.G.; Gogel, B.G.; Cullis, B.R.; Thompson, R. Navigating from ASReml-R Version 3 to 4; VSN Int. Ltd.: Hemel Hempstead, UK, 2017; Available online: https://asreml.kb.vsni.co.uk/wp-content/uploads/sites/3/2018/07/Navigating-from-ASReml-R-3-to-4.pdf.pdf (accessed on 15 November 2021).
- Cao, Y.Y.; Duan, H.; Yang, L.N.; Wang, Z.Q.; Zhou, S.C.; Yang, J.C. Effect of Heat Stress During Meiosis on Grain Yield of Rice Cultivars Differing in Heat Tolerance and Its Physiological Mechanism. Acta Agron. Sin. 2008, 34, 2134–2142. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Delgado, M.I.B.; Gutiérrez-Rodríguez, M.; Saavedra, A.L. Photosynthesis of wheat in a warm, irrigated environment: I: Genetic diversity and crop productivity. Field Crops Res. 2000, 66, 37–50. [Google Scholar] [CrossRef]
- Bányai, J.; Karsai, I.; Balla, K.; Kiss, T.; Bedő, Z.; Lang, L. Heat stress response of wheat cultivars with different ecological adaptation. Cereal Res. Commun. 2014, 42, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Mondal, S.; Mason, R.E.; Huggins, T.; Hays, D.B. QTL on wheat (Triticum aestivum L.) chromosomes 1B, 3D and 5A are associated with constitutive production of leaf cuticular wax and may contribute to lower leaf temperatures under heat stress. Euphytica 2015, 201, 123–130. [Google Scholar] [CrossRef]
- Sharma, D.K.; Andersen, S.B.; Ottosen, C.-O.; Rosenqvist, E. Wheat cultivars selected for high Fv/Fm under heat stress maintain high photosynthesis, total chlorophyll, stomatal conductance, transpiration and dry matter. Physiol. Plant. 2015, 153, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Shirdelmoghanloo, H.; Cozzolino, D.; Lohraseb, I.; Collins, N.C. Truncation of grain filling in wheat (Triticum aestivum) triggered by brief heat stress during early grain filling: Association with senescence responses and reductions in stem reserves. Funct. Plant Biol. 2016, 43, 919–930. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M. Rapid photosynthetic adaptation to heat stress triggered in potato leaves by moderately elevated temperatures. Plant Cell Environ. 1993, 16, 461–467. [Google Scholar] [CrossRef]
- Reddy, K.R.; Bheemanahalli, R.; Saha, S.; Singh, K.; Lokhande, S.B.; Gajanayake, B.; Read, J.J.; Jenkins, J.N.; Raska, D.A.; De Santiago, L.M.; et al. High-Temperature and Drought-Resilience Traits among Interspecific Chromosome Substitution Lines for Genetic Improvement of Upland Cotton. Plants 2020, 9, 1747. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Sharma, P.; Sareen, S.; Sarial, A.K. Mapping QTLs for chlorophyll content and chlorophyll fluorescence in wheat under heat stress. Biol. Plant. 2018, 62, 721–731. [Google Scholar] [CrossRef]
- Feng, B.; Liu, P.; Li, G.; Dong, S.T.; Wang, F.H.; Kong, L.A.; Zhang, J.W. Effect of Heat Stress on the Photosynthetic Characteristics in Flag Leaves at the Grain-Filling Stage of Different Heat-Resistant Winter Wheat Varieties. J. Agron. Crop Sci. 2014, 200, 143–155. [Google Scholar] [CrossRef]
- Ali, M.; Ibrahim, A.M.H.; Hays, D.B.; Ristic, Z.; Fu, J. Wild Tetraploid Wheat (Triticum turgidum L.) Response to Heat Stress. J. Crop Improv. 2010, 24, 228–243. [Google Scholar] [CrossRef]
- Bergkamp, B.; Impa, S.; Asebedo, A.; Fritz, A.; Jagadish, S.K. Prominent winter wheat varieties response to post-flowering heat stress under controlled chambers and field based heat tents. Field Crops Res. 2018, 222, 143–152. [Google Scholar] [CrossRef]
- Bennett, D.; Reynolds, M.; Mullan, D.; Izanloo, A.; Kuchel, H.; Langridge, P.; Schnurbusch, T. Detection of two major grain yield QTL in bread wheat (Triticum aestivum L.) under heat, drought and high yield potential environments. Theor. Appl. Genet. 2012, 125, 1473–1485. [Google Scholar] [CrossRef]
- Telfer, P.; Edwards, J.; Bennett, D.; Ganesalingam, D.; Able, J.; Kuchel, H. A field and controlled environment evaluation of wheat (Triticum aestivum) adaptation to heat stress. Field Crops Res. 2018, 229, 55–65. [Google Scholar] [CrossRef]
- Fábián, A.; Sáfrán, E.; Szabó-Eitel, G.; Barnabás, B.; Jäger, K. Stigma Functionality and Fertility Are Reduced by Heat and Drought Co-stress in Wheat. Front. Plant Sci. 2019, 10, 244. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, L.; Shi, P.; Yang, B.; Sun, T.; Cao, W.; Zhu, Y. Effects of short-term high temperature on grain quality and starch granules of rice (Oryza sativa L.) at post-anthesis stage. Protoplasma 2017, 254, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Erena, M.F. Genetic and Physiological Bases of Heat-Induced Floret Sterility in Wheat. Ph.D. Thesis, The University of Adelaide, Adelaide, Australia, 2018. Available online: https://hdl.handle.net/2440/122076 (accessed on 15 July 2021).
- Balla, K.; Karsai, I.; Kiss, T.; Horváth, A.; Berki, Z.; Cseh, A.; Bónis, P.; Árendás, T.; Veisz, O. Single versus repeated heat stress in wheat: What are the consequences in different developmental phases? PLoS ONE 2021, 16, e0252070. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Han, Y.; Wang, S.; Yin, S.; Peng, Z.; Zhou, M.; Gao, W.; Wen, X.; Qin, X.; Siddique, K. The Effect of Grain Position on Genetic Improvement of Grain Number and Thousand Grain Weight in Winter Wheat in North China. Front. Plant Sci. 2018, 9, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philipp, N.; Weichert, H.; Bohra, U.; Weschke, W.; Schulthess, A.W.; Weber, H. Grain number and grain yield distribution along the spike remain stable despite breeding for high yield in winter wheat. PLoS ONE 2018, 13, e0205452. [Google Scholar] [CrossRef] [Green Version]
- Pinto, R.S.; Lopes, M.S.; Collins, N.C.; Reynolds, M.P. Modelling and genetic dissection of staygreen under heat stress. Theor. Appl. Genet. 2016, 129, 2055–2074. [Google Scholar] [CrossRef] [Green Version]
- Qin, D.; Wu, H.; Peng, H.; Yao, Y.; Ni, Z.; Li, Z.; Zhou, C.; Sun, Q. Heat stress-responsive transcriptome analysis in heat susceptible and tolerant wheat (Triticum aestivum L.) by using Wheat Genome Array. BMC Genom. 2008, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Mufti, M.U. Screening Wheat Seedlings for Heat and Drought Tolerance by In Vivo Chlorophyll Fluorescence. Ph.D. Thesis, The University of Adelaide, Adelaide, Australia, 2005. Available online: https://hdl.handle.net/2440/22299 (accessed on 15 October 2021).
- Maphosa, L.; Langridge, P.; Taylor, H.; Parent, B.; Emebiri, L.; Kuchel, H.; Reynolds, M.P.; Chalmers, K.J.; Okada, A.; Edwards, J.; et al. Genetic control of grain yield and grain physical characteristics in a bread wheat population grown under a range of environmental conditions. Theor. Appl. Genet. 2014, 127, 1607–1624. [Google Scholar] [CrossRef]
- Tura, H.; Edwards, J.; Gahlaut, V.; Garcia, M.; Sznajder, B.; Baumann, U.; Shahinnia, F.; Reynolds, M.P.; Langridge, P.; Balyan, H.S.; et al. QTL analysis and fine mapping of a QTL for yield-related traits in wheat grown in dry and hot environments. Theor. Appl. Genet. 2020, 133, 239–257. [Google Scholar] [CrossRef]
- Mason, R.E.; Mondal, S.; Beecher, F.W.; Pacheco, A.; Jampala, B.; Ibrahim, A.M.H.; Hays, D.B. QTL associated with heat susceptibility index in wheat (Triticum aestivum L.) under short-term reproductive stage heat stress. Euphytica 2010, 174, 423–436. [Google Scholar] [CrossRef]
- Alsamadany, H. Diversity and Genetic Studies of Heat Tolerance in Wheat. Ph.D. Thesis, The University of Western Australia, Perth, Australia, 2015. Available online: https://research-repository.uwa.edu.au/en/publications/diversity-and-genetic-studies-of-heat-tolerance-in-wheat (accessed on 15 March 2021).
- Thistlethwaite, R.; Bokshi, A.; He, S.; Joukhadar, R.; Tan, D.; Daetwyler, H.; Trethowan, R. How Heat Tolerant are Our Current Wheat Varieties? 2021. GRDC Update Paper. Available online: https://grdc.com.au/resources-and-publications/grdc-update-papers/tab-content/grdc-update-papers/2021/02/how-heat-tolerant-are-our-current-wheat-varieties (accessed on 19 April 2022).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Abbreviation (Units) | Random Effects (Variance Components) | Fixed Effect (Wald Statistic) | Temperature Treatment Means | ||||
|---|---|---|---|---|---|---|---|---|
| Cultivar | Cultivar × Temperature Treatment | Error | Temperature Treatment | Control | Heat | Percent Reduction | ||
| Main stem traits | ||||||||
| Degrees of freedom | 29 | 29 | 116 | 1 | ||||
| Grain number | GN | 63.74 *** | 3.67 × 10−6 | 55.72 *** | 33.63 *** | 54.65 | 48.23 | 11.8 |
| Grain yield | GY (g) | 0.04 ** | 4.11 × 10−7 | 0.05 *** | 20.90 *** | 1.09 | 0.94 | 14.1 |
| Individual grain weight | GW (mg) | 12.5 ** | 9.85 × 10−7 | 16.89 *** | 0.88 | 20.28 | 19.70 | 2.9 |
| Spike length | SL (cm) | 1.46 *** | 1.44 × 10−7 | 0.56 *** | 2.09 | 9.54 | 9.37 | 1.8 |
| Spikelet number | SN | 4.99 *** | 1.25 × 10−6 | 2.01 *** | 1.77 | 18.38 | 18.09 | 1.6 |
| Main stem length | MSL (cm) | 37.33 *** | 1.59 | 23.66 *** | 0.93 | 51.77 | 52.64 | −1.7 |
| Pollen viability | PV (%) | 0.90 ** | 0.05 | 1.15 *** | 212 *** | 98.72 | 96.21 | 2.5 |
| Whole plant traits | ||||||||
| Degrees of freedom | 29 | 29 | 116 | |||||
| Above-ground biomass | BM (g) | 36.39 *** | 1.65 × 10−6 | 33.91 *** | 9.98 ** | 30.19 | 27.38 | 9.3 |
| Total no. tillers | TNT | 21.14 ** | 6.39 × 10−6 | 24.70 *** | 1.07 | 22.6 | 21.84 | 3.6 |
| Effective no. tillers | ENT | 27.47 *** | 1.27 × 10−6 | 21. 23 *** | 0.31 | 18.83 | 18.43 | 2.1 |
| % effective tillers | ET (%) | 98.24 *** | 2.97 × 10−5 | 119.67 *** | 0.23 | 82.85 | 83.65 | −1.0 |
| Total plant grain yield | TGY (g) | 5.82 ** | 2.12 × 10−6 | 11.07 *** | 4.99 * | 9.48 | 8.3 | 11.7 |
| Physiological traits | ||||||||
| Flag leaf chlorophyll content | ||||||||
| Degrees of freedom | 29 | 29 | 116 | |||||
| Day 1 | SPAD_1D | 4.01 *** | 2.82 × 10−7 | 5.10 *** | 0.90 | 46.13 | 46.47 | −0.7 |
| Day 3 | SPAD_3D | 3.69 ** | 0.35 | 4.69 *** | 3.10 | 50.47 | 51.12 | −1.3 |
| Day 5 | SPAD_5D | 3.37 ** | 0.57 | 4.31 *** | 2.40 | 53.54 | 54.08 | −1.0 |
| Flag leaf temperature depression | ||||||||
| Degrees of freedom | 29 | 29 | 116 | |||||
| Day 1 | FLTD_1D (°C) | 1.79 × 10−7 | 4.79 × 10−7 | 0.39 *** | 120.17 *** | 1.75 | 2.80 | −60.2 |
| Day 3 | FLTD_3D (°C) | 2.52 × 10−7 | 0.05 | 0.36 *** | 74.27 *** | 1.95 | 2.86 | −46.6 |
| Day 5 | FLTD_5D (°C) | 8.89 × 10−8 | 0.05 | 0.29 *** | 29.51 *** | 2.17 | 2.71 | −25.3 |
| Quantum yield of photosynthesis | ||||||||
| Degrees of freedom | 29 | 29 | 116 | |||||
| Day 1 | Fv/Fm_1D | 1.62 × 10−5 * | 1.35 × 10−8 | 5.67 × 10−5 *** | 192 *** | 0.817 | 0.802 | 1.8 |
| Day 3 | Fv/Fm_3D | 4.83 × 10−6 * | 8.15 × 10−9 | 2.05 × 10−5 *** | 300 *** | 0.819 | 0.806 | 1.6 |
| Day 5 | Fv/Fm_5D | 6.35 × 10−9 | 9.11 × 10−6 * | 1.88 × 10−5 *** | 145 *** | 0.821 | 0.809 | 1.5 |
| Stomatal conductance | ||||||||
| Degrees of freedom | 7 | 1 | 30 | 1 | ||||
| Day 3 | SC (mmol m−2 s−1) | 1182.91 | 1.16 × 10−3 | 1741.32 *** | 37.76 *** | 259.48 | 333.48 | −28.72 |
| Traits | Group 1 (Tolerant) | Group 2 (Sensitive) | Group 3 (Sensitive) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Heat | Percent Reduction | Control | Heat | Percent Reduction | Control | Heat | Percent Reduction | |
| GN | 56.57 | 52.43 | 7.3 | 51.78 | 43.44 | 16.1 | 53.68 | 45.22 | 15.8 |
| GY | 1.17 | 1.10 | 5.5 | 1.13 | 0.79 | 29.8 | 0.96 | 0.80 | 17.5 |
| GW | 20.9 | 21.4 | –2.0 | 22.2 | 18.6 | 16.4 | 18.1 | 18.0 | 0.8 |
| PV | 98.43 | 95.69 | 2.8 | 98.58 | 96.45 | 2.2 | 99.21 | 96.79 | 2.4 |
| BM | 29.40 | 29.38 | 0.1 | 32.33 | 28.45 | 12.0 | 30.0 | 23.95 | 20.2 |
| SL | 9.72 | 9.70 | 0.2 | 9.12 | 8.78 | 3.7 | 9.55 | 9.27 | 2.9 |
| SN | 17.81 | 17.79 | 0.1 | 17.78 | 17.00 | 4.4 | 19.55 | 19.18 | 1.9 |
| MSL | 51.92 | 52.72 | –1.5 | 55.19 | 54.14 | 1.9 | 49.51 | 51.62 | –4.3 |
| TGY | 10.2 | 10.3 | –2.6 | 9.3 | 7.0 | 22.7 | 8.1 | 6.1 | 17.4 |
| TNT | 22.0 | 23.7 | –7.5 | 23.7 | 20.9 | 11.3 | 22.6 | 19.5 | 13.0 |
| ENT | 19.6 | 21.2 | –8.8 | 17.9 | 15.4 | 12.8 | 18.0 | 16.0 | 9.3 |
| ET | 88.35 | 89.26 | –1.0 | 75.00 | 73.83 | 1.6 | 79.85 | 81.68 | –2.3 |
| SPAD_1D | 46.56 | 46.27 | 0.6 | 45.51 | 45.58 | –0.2 | 45.92 | 47.27 | –2.9 |
| SPAD_3D | 51.35 | 51.17 | 0.4 | 49.65 | 50.09 | –0.9 | 49.73 | 51.67 | –3.9 |
| SPAD_5D | 54.12 | 54.66 | –1.0 | 52.52 | 52.83 | –0.6 | 53.32 | 54.01 | –1.3 |
| FLTD_1D | 1.70 | 2.83 | –66.8 | 1.93 | 2.67 | –37.9 | 1.72 | 2.85 | –66.2 |
| FLTD_3D | 1.81 | 2.87 | –58.4 | 1.84 | 3.01 | –63.0 | 2.20 | 2.75 | –24.8 |
| FLTD_5D | 2.04 | 2.74 | –34.0 | 1.99 | 3.22 | –61.6 | 2.44 | 2.37 | 2.8 |
| Fv/Fm_1D | 0.817 | 0.802 | 1.9 | 0.815 | 0.802 | 1.6 | 0.817 | 0.802 | 1.8 |
| Fv/Fm_3D | 0.819 | 0.806 | 1.6 | 0.818 | 0.804 | 1.6 | 0.819 | 0.806 | 1.6 |
| Fv/Fm_5D | 0.822 | 0.810 | 1.5 | 0.820 | 0.809 | 1.3 | 0.820 | 0.806 | 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choudhary, M.; Yan, G.; Siddique, K.H.M.; Cowling, W.A. Heat Stress during Meiosis Has Lasting Impacts on Plant Growth and Reproduction in Wheat (Triticum aestivum L.). Agronomy 2022, 12, 987. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12050987
Choudhary M, Yan G, Siddique KHM, Cowling WA. Heat Stress during Meiosis Has Lasting Impacts on Plant Growth and Reproduction in Wheat (Triticum aestivum L.). Agronomy. 2022; 12(5):987. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12050987
Chicago/Turabian StyleChoudhary, Mukesh, Guijun Yan, Kadambot H. M. Siddique, and Wallace A. Cowling. 2022. "Heat Stress during Meiosis Has Lasting Impacts on Plant Growth and Reproduction in Wheat (Triticum aestivum L.)" Agronomy 12, no. 5: 987. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12050987