
Delayed Sowing Date Improves Rice Cooking and Taste Quality by Regulating the Quantity and Quality of Grains Located on Secondary Branches

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Experimental Design
2.3. Observations and Measurements
2.3.1. Development Stage
2.3.2. Grains Numbers in Different Positions
2.3.3. Grain Filling Characteristics
2.3.4. RVA
2.3.5. Cooking Quality and Texture Properties
2.3.6. Starch Extraction and Amylose Content
2.3.7. Enzyme Extraction and Assay
2.4. Statistics Analyses
3. Results
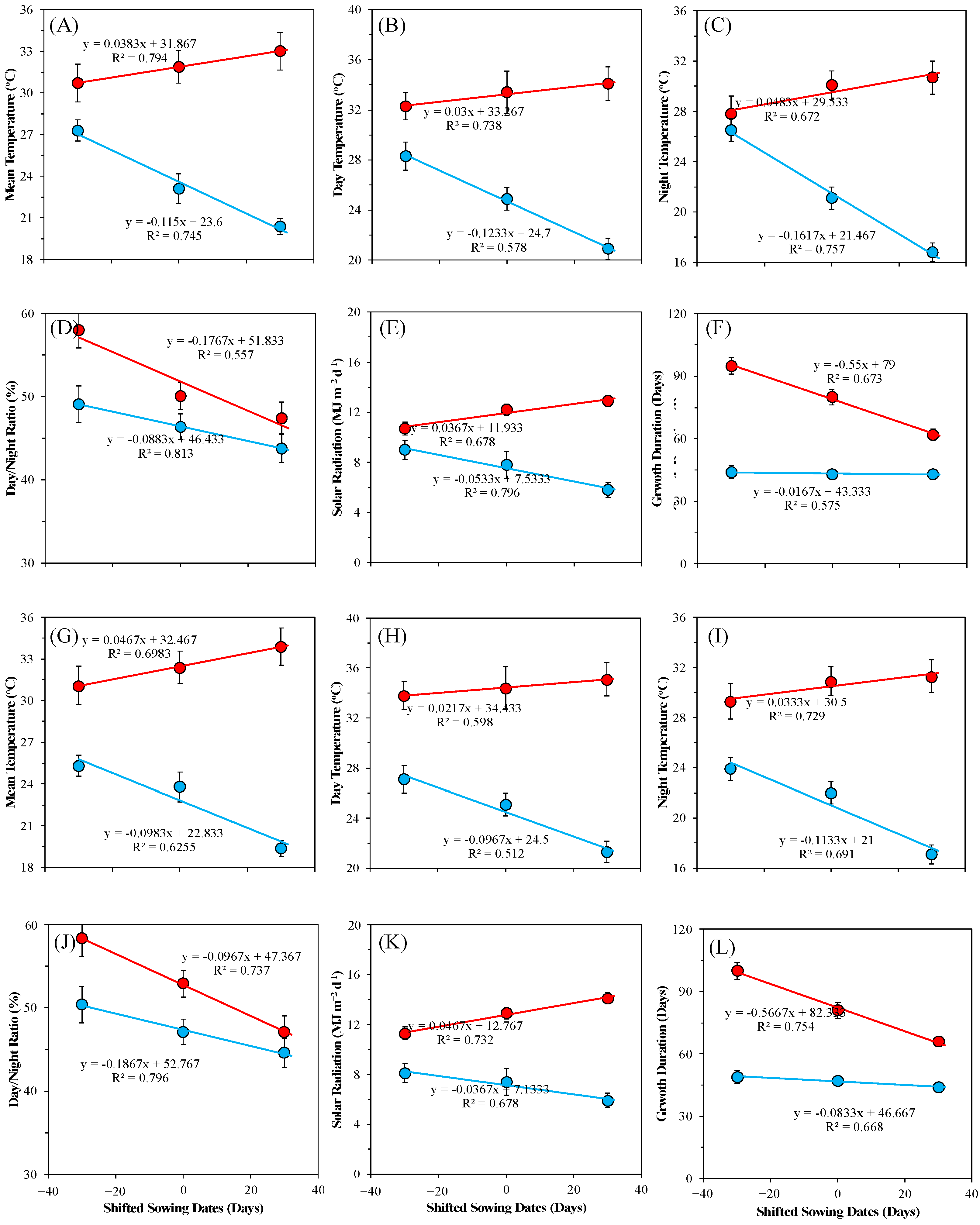
3.1. Effects of Sowing Dates Variation on Thermal Resources during Rice Growing Season
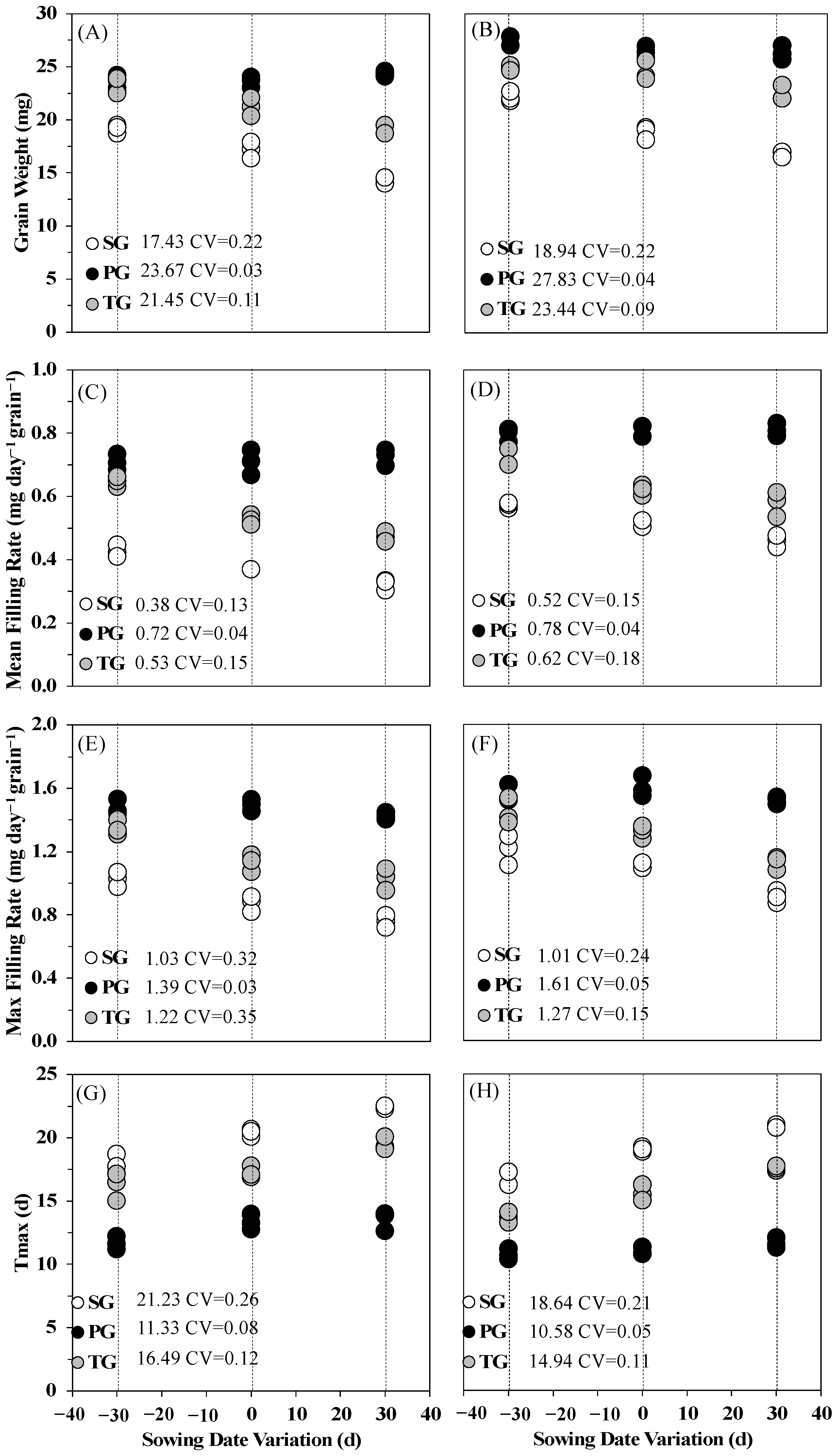
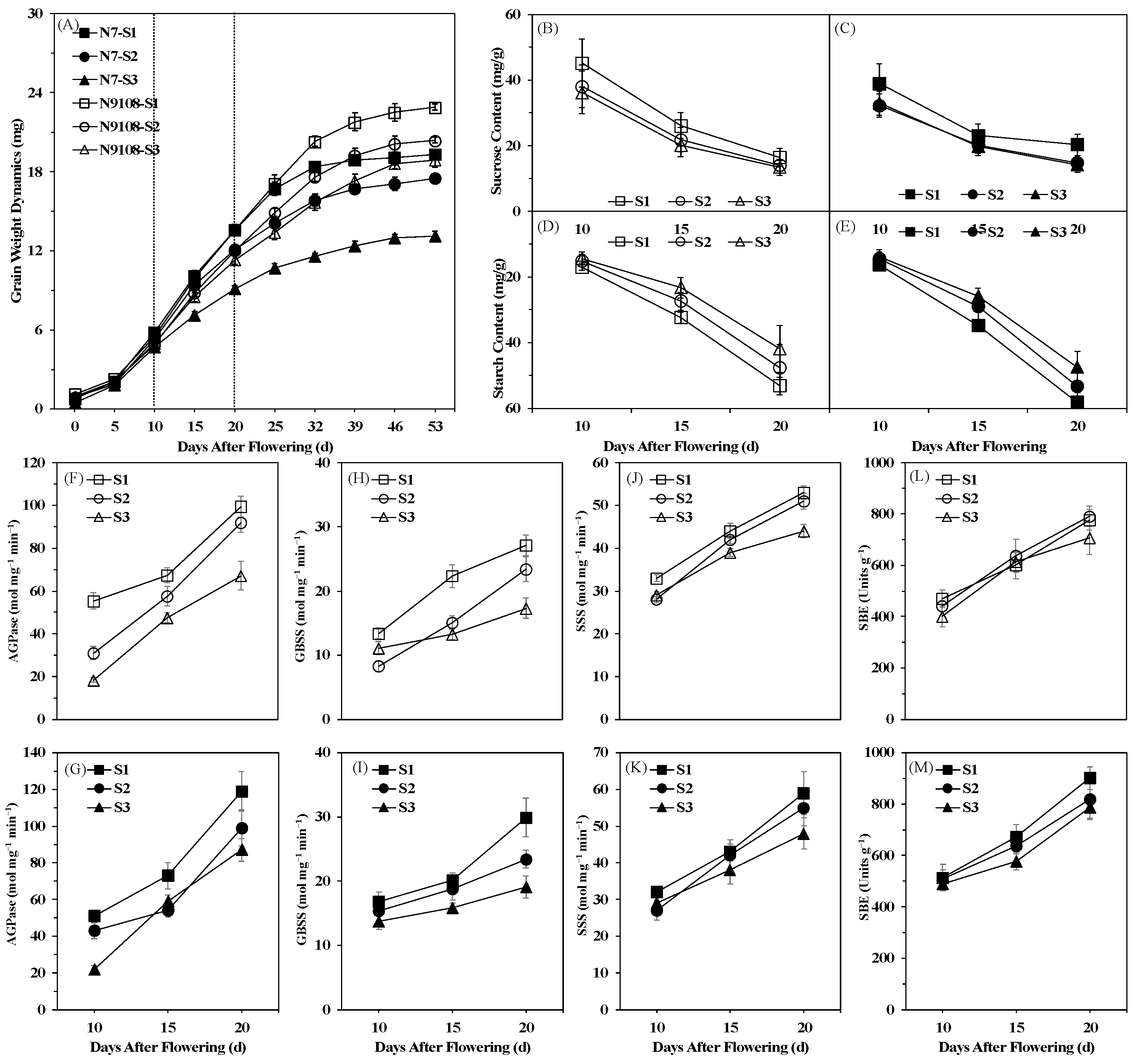
3.2. Effects of Sowing Date Variation on Rice Grain Filling
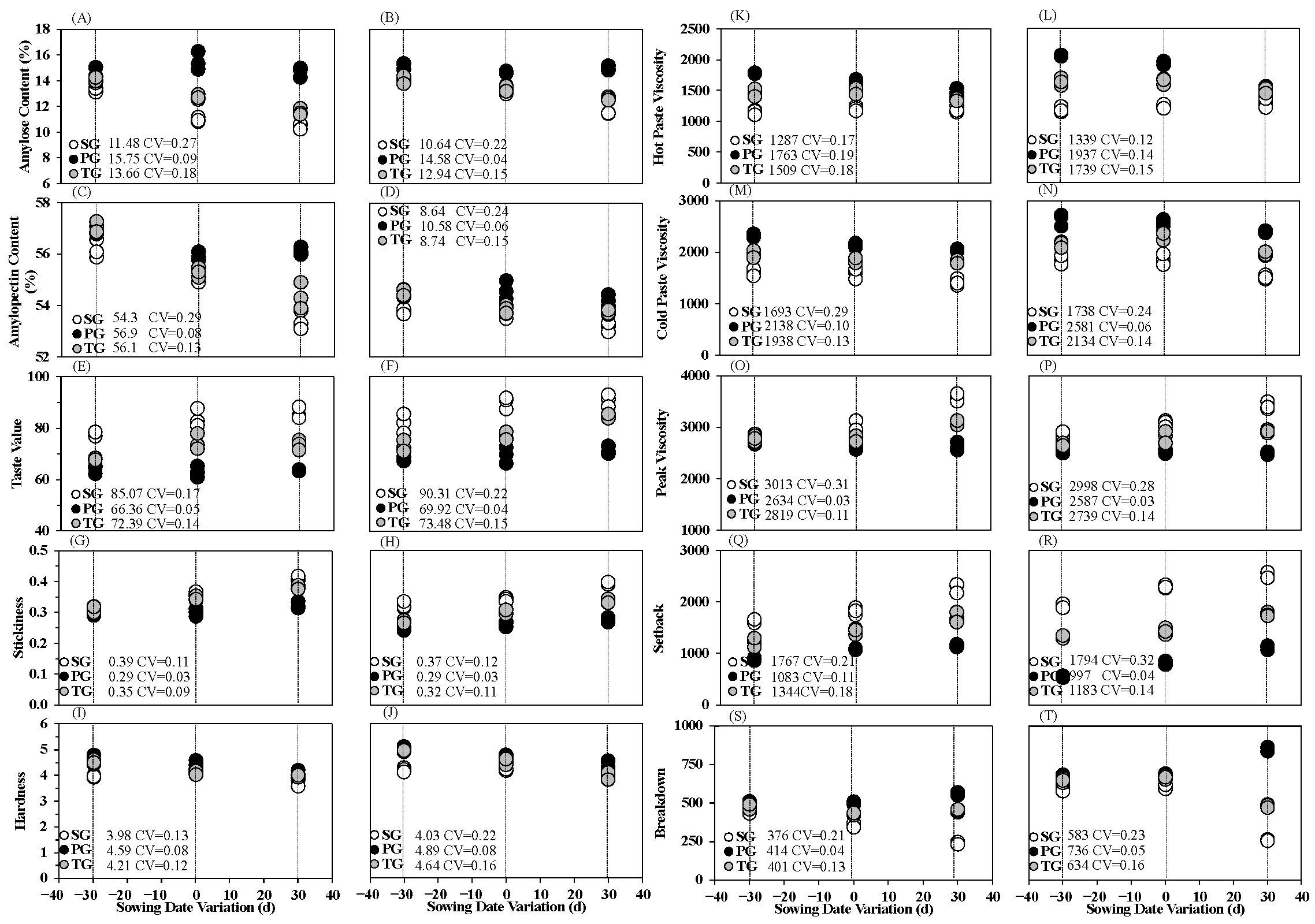
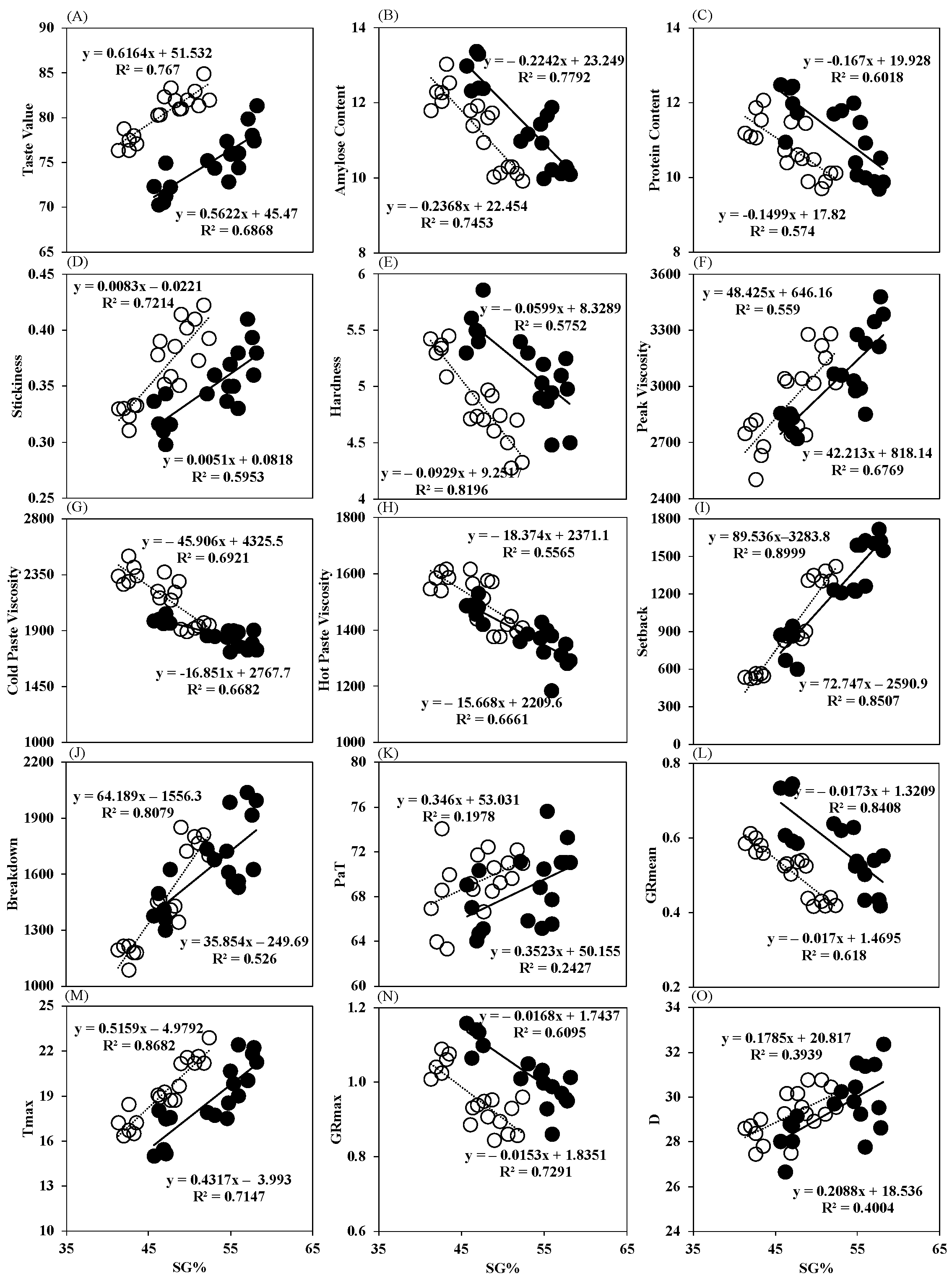
3.3. Effects of Sowing Date Variation on Rice Grain Cooking and Taste Quality
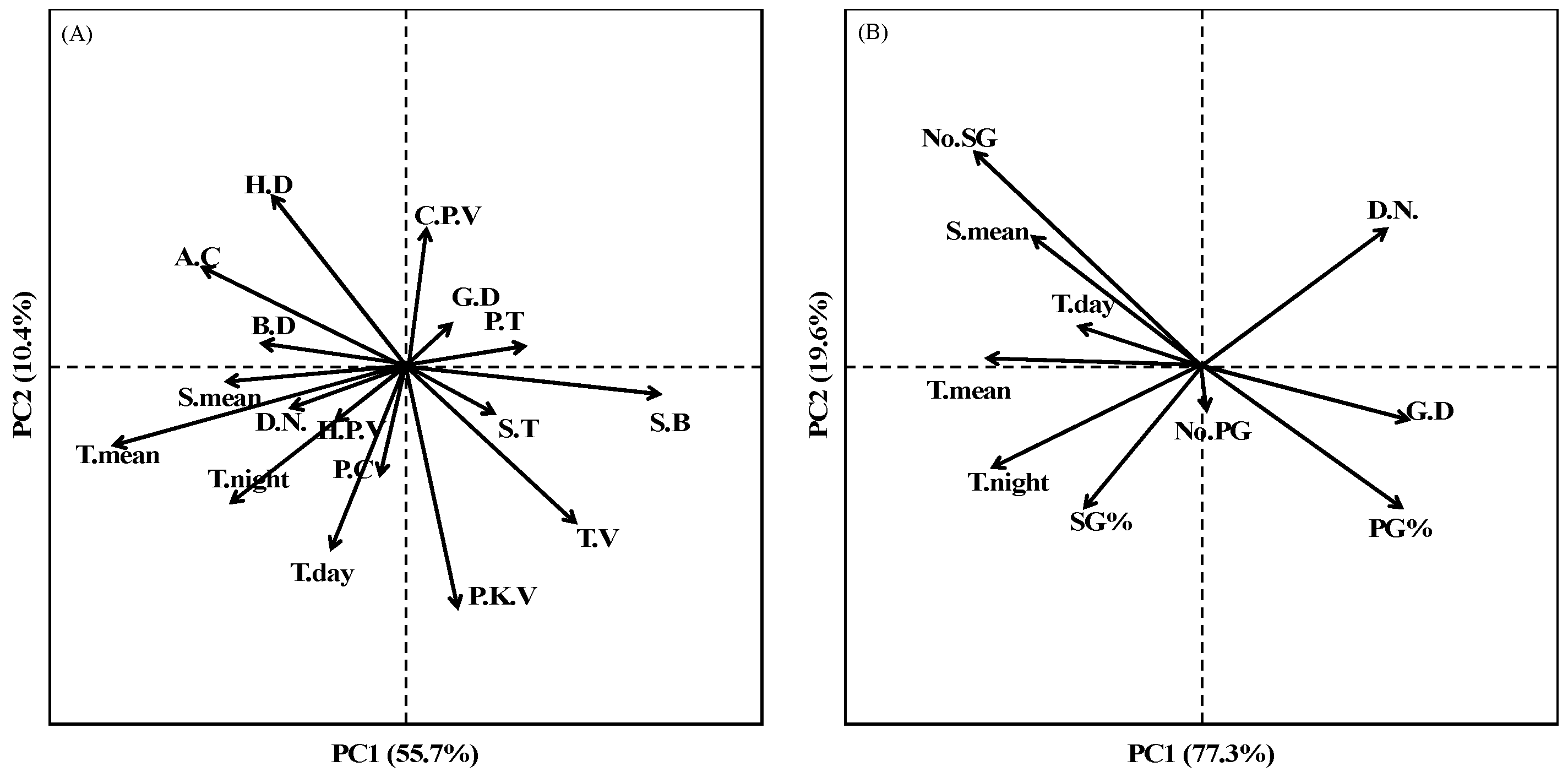
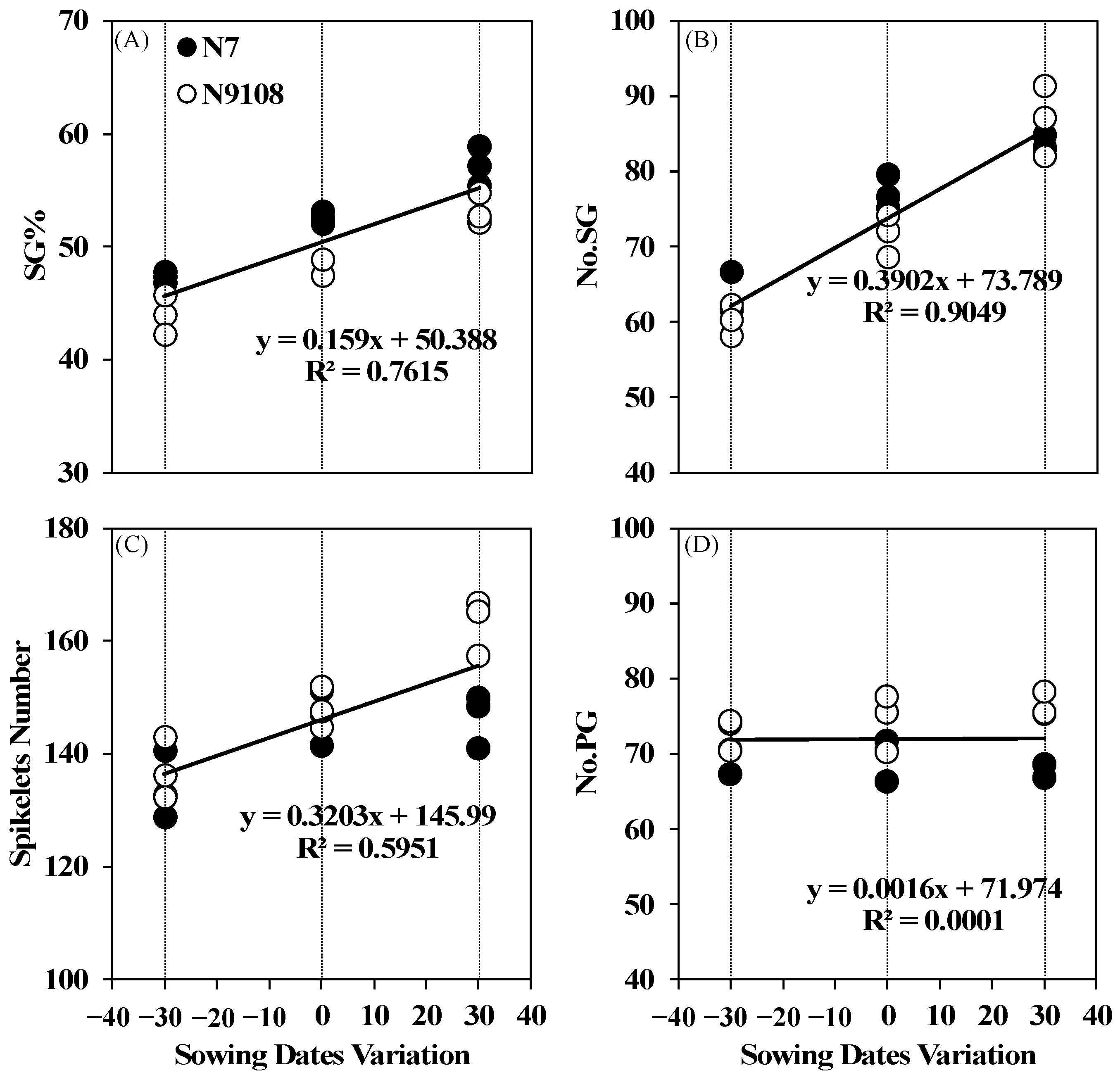
3.4. Effects of Sowing Date Variation on Rice Spikelets Branching Structure
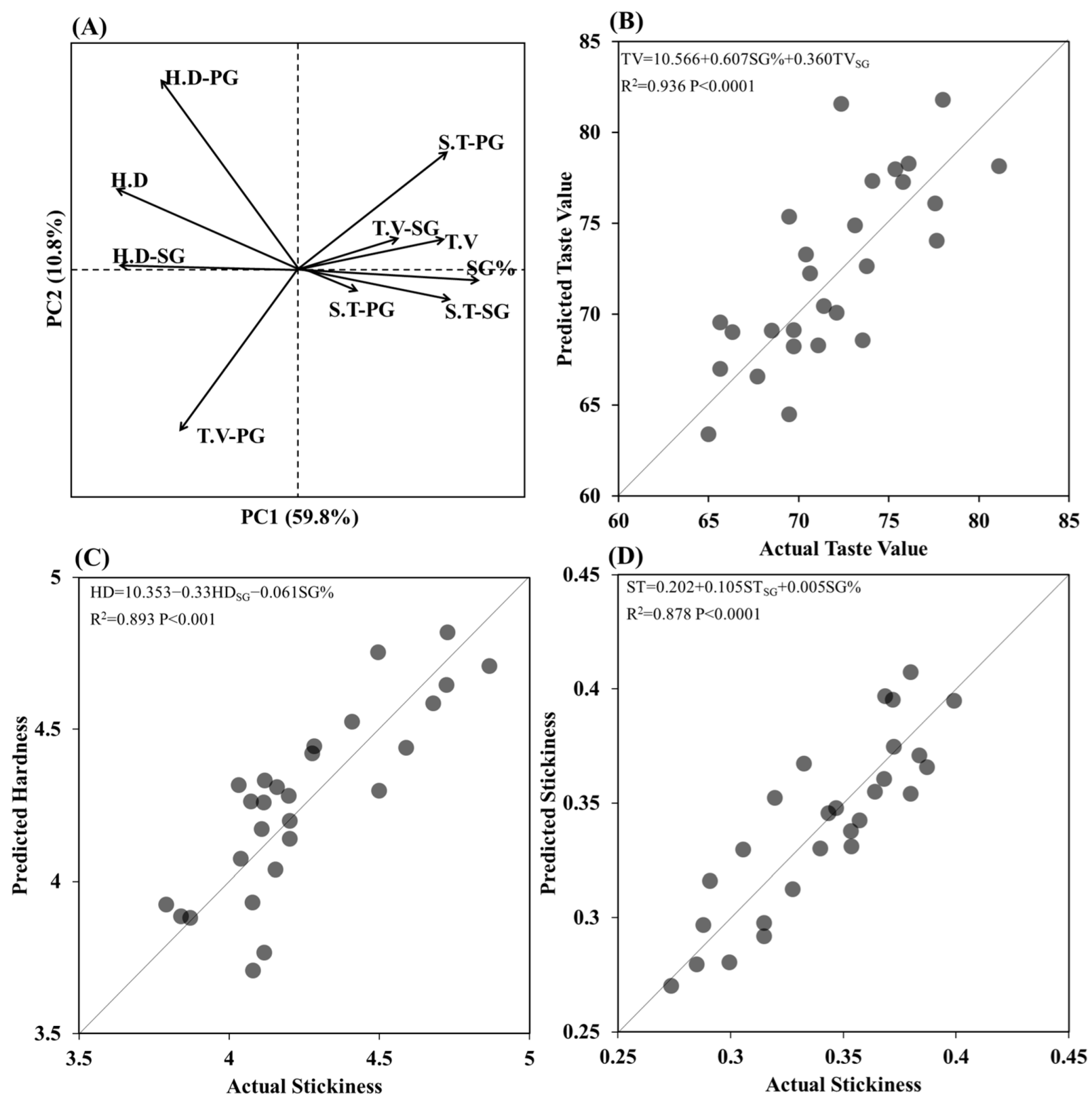
3.5. Correlation Analysis of Structural and Physiological Parameters of Grains Located on SG, PG and TG
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, C.; Yang, F.; Tang, X.; Lu, B.; Li, Z.; Liu, Z.; Ding, Y.; Ding, C.; Li, G. Super Rice With High Sink Activities Has Superior Adaptability to Low Filling Stage Temperature. Front. Plant Sci. 2021, 12, 729021. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Cui, W.; Xu, K.; Gao, H.; Zhang, H. Effects of Early- and Late-Sowing on Starch Accumulation and Associated Enzyme Activities During Grain Filling Stage in Rice. Rice Sci. 2021, 28, 191–199. [Google Scholar]
- Deng, N.; Ling, X.; Sun, Y.; Zhang, C.; Fahad, S.; Peng, S.; Cui, K.; Nie, L.; Huang, J. Influence of temperature and solar radiation on grain yield and quality in irrigated rice system. Eur. J. Agron. 2015, 64, 37–46. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, W.; Zhuang, Q.; Luo, Y. Adaptation of paddy rice in China to climate change: The effects of shifting sowing date on yield and irrigation water requirement. Agric. Water Manag. 2019, 228, 105890. [Google Scholar] [CrossRef]
- Zhou, N.B.; Zhang, J.; Fang, S.L.; Wei, H.Y.; Zhang, H.C. Effects of temperature and solar radiation on yield of good eating-quality rice in the lower reaches of the Huai River Basin, China. Sci. Agric. Sin. 2021, 20, 1762–1774. [Google Scholar] [CrossRef]
- Pan, T.; Zhao, L.; Lin, L.; Wang, J.; Liu, Q.; Wei, C. Changes in kernel morphology and starch properties of high-amylose brown rice during the cooking process. Food Hydrocoll. 2017, 66, 227–236. [Google Scholar] [CrossRef]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and Genetic Analysis of the Effects ofAmylose-Extender Mutation in Rice Endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef]
- Kato, T.; Shinmura, D.; Taniguchi, A. Activities of Enzymes for Sucrose-Starch Conversion in Developing Endosperm of Rice and Their Association with Grain Filling in Extra-Heavy Panicle Types. Plant Prod. Sci. 2007, 10, 442–450. [Google Scholar] [CrossRef]
- Satoh, H.; Nishi, A.; Yamashita, K.; Takemoto, Y.; Tanaka, Y.; Hosaka, Y.; Sakurai, A.; Fujita, N.; Nakamura, Y. Starch-branching enzyme I-deficient mutation specifically affects the structure and properties of starch in rice endosperm. Plant Physiol. 2003, 133, 1111–1121. [Google Scholar] [CrossRef] [Green Version]
- Bu-Hong, Z.; Wen-Jie, Z.; Zhi-Qin, W.; Qin-Sen, Z.; Jian-Chang, Y. Changes in Activities of the Key Enzymes Related to Starch Synthesis in Rice Grains During Grain Filling and Their Relationships with the Filling Rate and Cooking Quality. Sci. Agric. Sin. 2005, 4, 26–33. [Google Scholar]
- Siddik, M.A.; Zhang, J.; Chen, J.; Qian, H.; Jiang, Y.; Raheem, A.K.; Deng, A.; Song, Z.; Zheng, C.; Zhang, W. Responses of indica rice yield and quality to extreme high and low temperatures during the reproductive period. Eur. J. Agron. 2019, 106, 30–38. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Butardo, V.M., Jr.; Misra, G.; Cuevas, R.P.; Anacleto, R.; Kavi Kishor, P.B. Designing climate-resilient rice with ideal grain quality suited for high-temperature stress. J. Exp. Bot. 2015, 66, 1737–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, J. Grain-filling problem in ‘super’ rice. J. Exp. Bot. 2010, 61, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Fang, C.; Qian, Z.; Guo, B.; Huo, Z. Differences in starch structure, physicochemical properties and texture characteristics in superior and inferior grains of rice varieties with different amylose contents. Food Hydrocoll. 2021, 110, 106170. [Google Scholar] [CrossRef]
- Ling, Q.; Zhang, H. Discussion about Rice High Yield Population and Optimization. Sci. Agric. Sin. 1993, 26, 11. (In Chinese) [Google Scholar]
- Limpisut, P.; Jindal, V.K. Comparison of Rice Flour Pasting Properties using Brabender Viscoamylograph and Rapid Visco Analyser for Evaluating Cooked Rice Texture. Starch-Stärke 2002, 54, 350–357. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, L.; Zhu, Z.; Lu, H.; Zhou, X.; Qian, Y.; Li, Q.; Lu, Y.; Gu, M.; Liu, Q. Characterization of Grain Quality and Starch Fine Structure of Two Japonica Rice (Oryza Sativa) Cultivars with Good Sensory Properties at Different Temperatures during the Filling Stage. J. Agric. Food Chem. 2016, 64, 4048–4057. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, H.; Guo, B.; Xu, K.; Dai, Q.; Wei, C.; Wei, H.; Gao, H.; Hu, Y.; Cui, P. Effect of Nitrogen Management on the Structure and Physicochemical Properties of Rice Starch. J. Agric. Food Chem. 2016, 64, 8019–8025. [Google Scholar] [CrossRef]
- Man, J.; Yang, Y.; Zhang, C.; Zhou, X.; Wei, C. Structural Changes of High-Amylose Rice Starch Residues following in Vitro and in Vivo Digestion. J. Agric. Food Chem. 2012, 60, 9332–9341. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Wang, Z.; Zhu, Q.; Liu, L. Activities of enzymes involved in sucrose-to-starch metabolism in rice grains subjected to water stress during filling. Field Crops Res. 2003, 81, 69–81. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yuki, K.; Park, S.Y.; Ohya, T. Carbohydrate Metabolism in the Developing Endosperm of Rice Grains. Plant Cell Physiol. 1989, 30, 833–839. [Google Scholar] [CrossRef]
- Waongo, M.; Laux, P.; Kunstmann, H. Adaptation to climate change: The impacts of optimized planting dates on attainable maize yields under rainfed conditions in Burkina Faso. Agric. For. Meteorol. 2015, 205, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Zhang, C.; He, L.; Liao, S.; Li, Q.; Li, B.; Zhu, S.; Gao, Y.; Tao, Y.; Zhou, W. Delayed sowing date improves the quality of mechanically transplanted rice by optimizing temperature conditions during growth season. Field Crops Res. 2022, 281, 108493. [Google Scholar] [CrossRef]
- Balwinder-Singh; Humphreys, E.; Sudhir-Yadav; Gaydon, D.S. Options for increasing the productivity of the rice–wheat system of north-west India while reducing groundwater depletion. Part 1. Rice variety duration, sowing date and inclusion of mungbean. Field Crops Res. 2015, 173, 68–80. [Google Scholar] [CrossRef] [Green Version]
- Rurinda, J.; Wijk, M.; Mapfumo, P.; Descheemaeker, K.; Supit, I.; Giller, K. Climate change and maize yield in southern Africa: What can farm management do? Glob. Chang. Biol. 2016, 21, 4588–4601. [Google Scholar] [CrossRef]
- Shi, W.; Yin, X.; Struik, P.C.; Solis, C.; Xie, F.; Schmidt, R.C.; Huang, M.; Zou, Y.; Ye, C.; Jagadish, S.V.K. High day- and night-time temperatures affect grain growth dynamics in contrasting rice genotypes. J. Exp. Bot. 2017, 18, 5233–5245. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Q.; Li, H.X.; Feng, L.; Chen, M.X.; Meng, S.; Ye, N.H.; Zhang, J. Transcriptomic analysis of grain filling in rice inferior grains under moderate soil drying. J. Exp. Bot. 2019, 70, 1597–1611. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.H.; Chen, P.F.; Qiao, Z.Y.; Xiang-Zhou, W.U.; Zhao, B.H.; Jiang, Y.Y.; Yang, J.C. Quality Response of Grains in Different Spikelet Positions to Temperature Stress During Grain Filling of Rice. Acta Agron. Sin. 2011, 37, 506–513. [Google Scholar] [CrossRef]
- Xfa, B.; Yl, A.; Yun, Z.A.; Jw, A.; Jie, Z.A.; Xs, A.; Yp, A.; Xb, A.; Cza, B.; Dza, B. Characterization of physicochemical qualities and starch structures of two indica rice varieties tolerant to high temperature during grain filling. J. Cereal Sci. 2022, 93, 102966. [Google Scholar]
- Xia, N.; Zhao, H.W.; Yan-Chao, L.V.; Zhao, Z.D.; Zou, D.T.; Liu, H.L.; Wang, J.G.; Jia, Y. Effect of Cold-water Stress at Grain-filling Stage on Starch Accumulation and Related Enzyme Activities in Grains of japonica Rice in Cold-region. Chin. J. Rice Sci. 2016, 30, 62–74. [Google Scholar]
- Jing, L.; Chen, C.; Lu, Q.; Wang, Y.; Yang, L. How do elevated atmosphere CO2 and temperature alter the physiochemical properties of starch granules and rice taste? Sci. Total Environ. 2020, 766, 142592. [Google Scholar] [CrossRef] [PubMed]
- Cza, B.; Wh, B.; Yan, L.B.; Yong, Y.B.; Zc, B.; Ql, B.; Xf, B.; Jl, C.; Qla, B. A comparative evaluation of the effect of SSI and Wx allelic variation on rice grain quality and starch physicochemical properties. Food Chem. 2022, 371, 131205. [Google Scholar]
- Chen, H.; Chen, D.; He, L.; Wang, T.; Ren, W. Correlation of taste values with chemical compositions and Rapid Visco Analyser profiles of 36 indica rice (Oryza sativa L.) varieties. Food Chem. 2021, 349, 129176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, Y.Y.; Song, N.Y.; Chen, Q.L.; Zhao, Q.Z. Response of grain-filling rate and grain quality of mid-season indica rice to nitrogen application. J. Integr. Agric. 2021, 20, 1465–1473. [Google Scholar] [CrossRef]
- Le, X.; Zhan, X.; Yu, T.; Nie, L.; Huang, J.; Cui, K.; Fei, W.; Yong, L.; Peng, S. Yield performance of direct-seeded, double-season rice using varieties with short growth durations in central China. Field Crops Res. 2018, 227, 49–55. [Google Scholar]
- Cai, Y.X.; Zhu, Q.S.; Wei, X.U.; Wang, W.; Yang, J.C.; Zhang, Z.J.; Lang, Y.Z. Effects of Water Stress on the Main Characters of Superior and Inferior Grains Quality and the Properties of RVA Profile during Grain-filling Stage. Acta Agron. Sin. 2004, 30, 241–247. [Google Scholar]
- Chen, P.; Gu, J.; Han, L.; Qiao, Z.; Wang, J.; Zhu, Y.; Dong, M. Effects of returning wheat straw to farmland and irrigation pattern on grain setting traits and quality of super rice. Chin. J. Eco-Agric. 2014, 22, 543–550. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Correlations between Filling Characteristics and Taste Quality Parameters | |||||
|---|---|---|---|---|---|
| Taste Quality Characteristics | GRmean | Grmax | Tmax | D | |
| AC | 0.881 ** | 0.790 ** | −0.782 * | −0.782 * | |
| PC | 0.902 | 0.879 * | −0.834 | −0.678 * | |
| ST | −0.906 ** | −0.900 * | 0.851 ** | 0.617 ** | |
| HD | 0.787 ** | 0.823 * | −0.740 * | −0.604 * | |
| TV | −0.961 ** | −0.859 * | 0.847 * | 0.745 ** | |
| PKV | −0.917 ** | −0.838 ** | 0.866 ** | 0.571 * | |
| HPV | −0.498 * | −0.591 | 0.303 | 0.466 * | |
| BD | 0.719 * | 0.670 * | −0.752 * | −0.343 | |
| CPV | −0.916 * | −0.836 ** | 0.835 ** | 0.577 * | |
| SB | 0.946 ** | 0.820 * | −0.880 * | −0.705 * | |
| PT | −0.072 | −0.144 | 0.246 | −0.135 | |
| (b) Correlations between Filling Characteristics and Enzyme Activities | |||||
| Starch Related Enzymes | Days After Flowering | GRmean | GRmax | Tmax | D |
| AGPase | 5 | 0.962 ** | 0.842 * | −0.885 * | −0.763 * |
| 10 | 0.939 * | 0.837 ** | −0.884 ** | −0.653 * | |
| 15 | 0.886 ** | 0.851 * | −0.880 ** | −0.554 | |
| SSS | 5 | 0.356 | 0.202 | −0.216 | −0.539 |
| 10 | 0.901 ** | 0.763 ** | −0.777** | −0.803 ** | |
| 15 | 0.937 * | 0.878 * | −0.918 ** | −0.624 * | |
| GBSS | 5 | 0.631 * | 0.483 | −0.522 ** | −0.789 * |
| 10 | 0.501 | 0.518 | −0.568 * | −0.238 | |
| 15 | 0.720 ** | 0.609 * | −0.694 * | −0.409 | |
| SBE | 5 | 0.874 ** | 0.868 * | −0.828 * | −0.671 ** |
| 10 | −0.175 | −0.149 * | 0.005 | 0.306 | |
| 15 | 0.293 ** | 0.379 * | −0.327 | −0.021 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Lu, B.; Liang, L.; Yang, F.; Ding, C.; Yan, F.; Zhou, Y.; Jiang, Z.; Liu, Z.; Ding, Y.; et al. Delayed Sowing Date Improves Rice Cooking and Taste Quality by Regulating the Quantity and Quality of Grains Located on Secondary Branches. Agronomy 2022, 12, 1316. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061316
Xu C, Lu B, Liang L, Yang F, Ding C, Yan F, Zhou Y, Jiang Z, Liu Z, Ding Y, et al. Delayed Sowing Date Improves Rice Cooking and Taste Quality by Regulating the Quantity and Quality of Grains Located on Secondary Branches. Agronomy. 2022; 12(6):1316. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061316
Chicago/Turabian StyleXu, Congshan, Bo Lu, Limin Liang, Fei Yang, Chao Ding, Feiyu Yan, Yan Zhou, Zhengrong Jiang, Zhenghui Liu, Yanfeng Ding, and et al. 2022. "Delayed Sowing Date Improves Rice Cooking and Taste Quality by Regulating the Quantity and Quality of Grains Located on Secondary Branches" Agronomy 12, no. 6: 1316. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061316