1. Introduction
The diamondback moth,
Plutella xylostella (L.) (Lepidoptera: Plutellidae), is a widely distributed and economically important oligophagous pest of cruciferous vegetable crops [
1,
2]. Presently the control of
P. xylostella is mainly dependent on chemical insecticides. However, the injudicious application of insecticides has caused
P. xylostella to rapidly develop resistance to almost all classes of old fashioned insecticides [
3,
4,
5] as well as varying degrees of resistance to novel chemicals, such as chlorantraniliprole [
6], spinosad [
7], emamectin benzoate [
8] and spinetoram [
9]. Besides, owing to its high fecundity and short life span, very few insecticides are currently available that remain effective in controlling
P. xylostella in India [
9]. Therefore, new active compounds with unique mechanisms of action are urgently required to participate in insecticide resistance management programmes.
Fluxametamide is a novel isoxazoline insecticide that differs chemically from fiproles, avermectins and diamides. It has a new target site for arthropods, interfering with GABA Cl
− and Glu Cl
− channels [
10,
11], and is listed in group 30 in the IRAC mode of action classification [
12]. Fluxametamide has an exceptional insecticidal activity on a range of insect pest species, such as lepidoptera, thysanoptera, coleoptera and diptera [
13] and the advantage of negligible non-target toxicity, including pollinators [
10,
14]. The unique binding site of fluxametamide in GABA-gated chloride channels, different from those for existing antagonists, makes this molecule effective against fipronil-resistant pest populations [
15]. After Japan, Australia and China, fluxametamide has been launched in India very recently and is expected to be used effectively to control various lepidopteran insects. Besides lethal effects, the sublethal effects must be considered while making an insecticide selection [
16,
17]. The effect of sublethal exposure to an insecticide could impair the growth, developmental rate, feeding, fecundity, and various physiological and biochemical processes of an insect pest, and sometimes causes the rapid evolution of insecticide resistance [
18]. The extensive application of fluxametamide in the coming years may cause sublethal effects on
P. xylostella gradually. Therefore, estimation of the toxicity and sublethal effects of fluxametamide on
P. xylostella is very important for coordinating chemical and biological control strategies in an integrated pest management program. However, there is no report on the sublethal effects of fluxametamide on
P. xylostella, and very little literature is available for their comparison. Previous studies reported that fluxametamide has excellent efficacy against
Musca domestica,
Laodelphax striatellus,
Drosophila melanogaster,
Tetranichus urticae and
Cylas formicarius [
10,
19]. On the other hand, the sublethal concentrations of various insecticides such as chlorantraniliprole, metaflumizone, fenoxycarb, spinosad, spinetoram and broflanilide significantly reduced the longevity and fecundity of
P. xylostella [
20,
21,
22,
23,
24]. The life-table bioassay is a precise tool to estimate the effect of sublethal exposure to an insecticide on major biological factors and demographic traits of an insect population in the laboratory [
25,
26]. In addition to the pest’s biology, the sublethal effect of a new chemical on the biochemistry of a target insect must be considered for a comprehensive study [
27]. These pieces of information can be crucial in decision-making on pest management by predicting the possible impacts that are likely to occur in field strains.
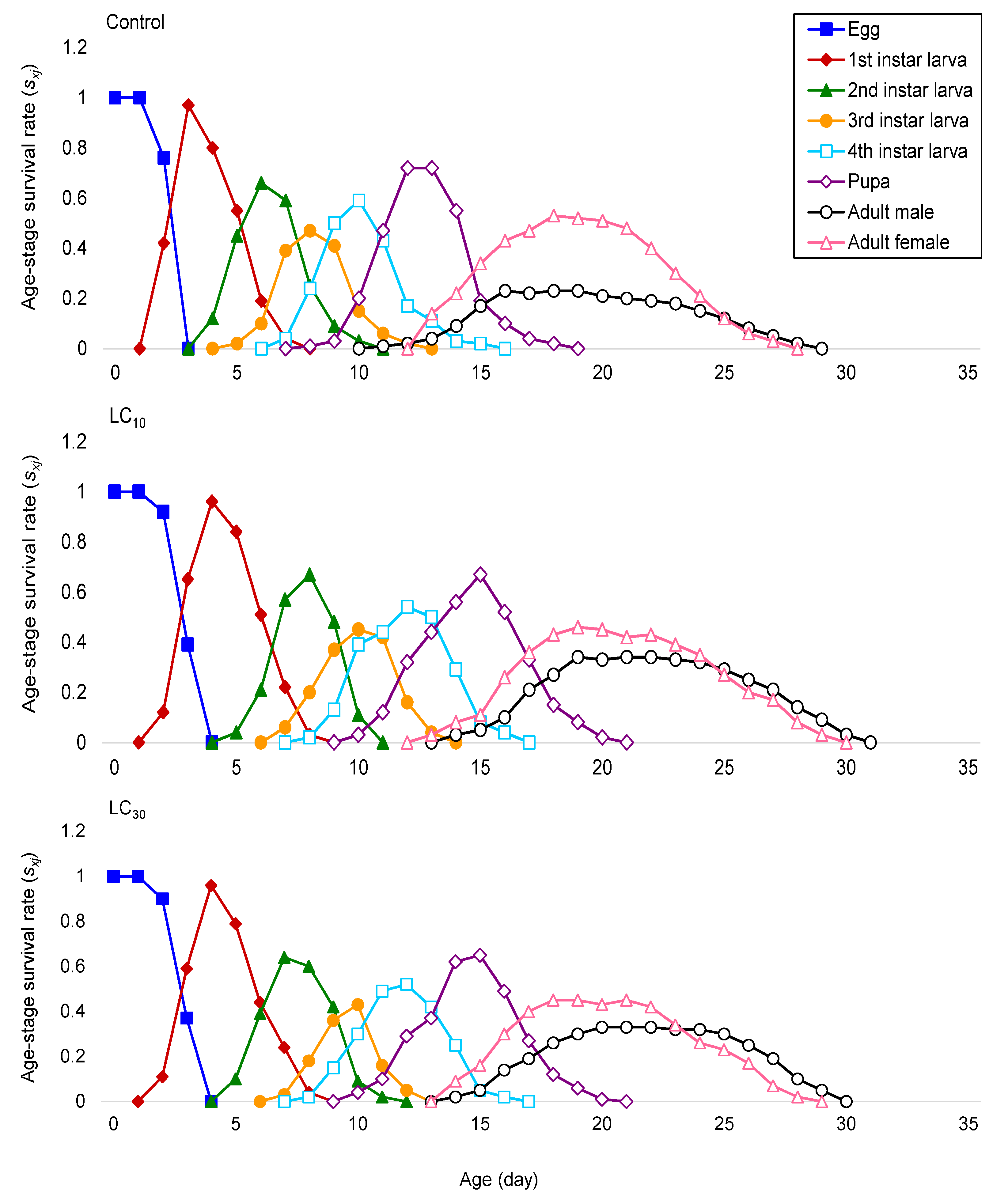
The present study aimed to evaluate the toxicity and baseline susceptibility of a novel isoxazoline insecticide fluxametamide against third instar larvae of a susceptible population of P. xylostella. Additionally, the age-stage and two-sex life table methods were used to analyze the demographic parameters of P. xylostella treated by sublethal concentrations (LC10 and LC30) of fluxametamide, with a particular focus on the potential toxicological and possible transgenerational effects in the subsequent generations. The activities of cytochrome P450 monooxygenase, glutathione S-transferase and esterases were also quantified to assess the biochemical responses and metabolic enzyme-mediated detoxification mechanisms to sublethal concentrations of fluxametamide which could provide crucial data for the field application guidelines of this insecticide to control P. xylostella in the future.
4. Discussion
Fluxametamide is a novel isoxazoline insecticide that can effectively control various lepidopteran and coleopteran insect pests [
19,
40]. In the present study, the authors tried to explore the potential influence of the sublethal concentrations of fluxametamide on
P. xylostella for the first time. A high insecticidal activity of fluxametamide was observed against the third-instar larvae of
P. xylostella with an LC
50 value of 0.18 mg L
−1. The result indicated that the fluxametamide toxicity to
P. xylostella was higher than that of abamectin, spinosad, indoxacarb, chlorantraniliprole and fenoxycarb [
21,
41,
42]. In addition, the second instar larvae of
P. xylostella exhibited an LC
50 value of 56.41 mg L
−1 when treated with farnesyl acetate, was higher than the present results with a lower toxicity [
43]. However, the LC
50 of the technical grade fluxametamide, obtained from the present study, was also lower than that of
rac-fluxametamide (1.90 mg L
−1) and its enantiomers;
S-(+)-fluxametamide (0.62 mg L
−1), or
R-(-)-fluxametamide (34.18 mg L
−1) at 48 h [
40].
Considering that most of the long-lasting insecticides could degrade gradually after their initial field application, turning lethal dosages into sublethal levels, we determined the effects of the sublethal concentrations of fluxametamide on
P. xylostella. Very few workers have reported the laboratory and field efficacy of fluxametamide against
Tetranichus urticae,
Laodelphax striatellus and sweet potato weevil [
10,
19]. In addition to larval toxicity, the present study demonstrated a variety of sublethal effects of fluxametamide on key biological traits of
P. xylostella. The LC
10 and LC
30 concentrations of fluxametamide not only prolonged the fourth larval duration of F
0 of
P. xylostella but also extended the larval period of F
1 progeny in a dose-dependent manner. This observation was corroborated by the previous reports on the sublethal effects of spinosad [
20] or chlorantraniliprole [
21] or spinetoram [
23] on
P. xylostella larvae. Prolongation of the larval duration in fluxametamide treatments could be attributed to retarded growth due to a reduced appetite, feeding disruption, abnormal metabolism, starvation stress, or the imbalance between metabolic detoxification and physiological development [
44,
45]. However, it is important to note that the extended larval duration could make a significant contribution to the field management of
P. xylostella by increasing the probability of natural parasitism or predation [
46] and by forcing immature larvae to feed on suboptimal nutritional foliage for completing their life cycle, which leads to reduced fecundity and survival [
47]. In contrast, the sublethal concentrations of cyantraniliprole and beta-cypermethrin did not show any significant effect on the life history of
Helicoverpa assulta [
48] and
P. xylostella [
49], respectively. Besides, da Silva et al. [
50] reported that chlorantraniliprole (<10% mortality) and methomyl (0% mortality) registered very negligible toxicity to
P. xylostella at their sublethal levels. This phenomenon might be attributable to the typical symptoms of positive fitness effects of sublethal insecticides exposure to insect pests [
51].
Fluxametamide at sublethal concentrations could persistently affect the metamorphosis and biology of treated
P. xylostella. For example, the LC
10 and LC
30 fluxametamide treatments significantly decreased the pupation rate in both F
0 and F
1 generations, which could, in turn, reduce the probability that the treated strain completed the developmental process. In addition, a significant increase in the deformed adult rate was observed at sublethal doses of fluxametamide in both generations. The adverse effect of chlorantraniliprole, spinosad and tebufenozide on the pupation rate of
Spodoptera exigua was already reported [
52]. Tamilselvan et al. [
23] also observed a significant reduction and increase in the pupation rate and the adults with wings deformity, respectively, in F
0 generation of
P. xylostella when third instar larvae were treated with sublethal doses of spinetoram. In the present study, exposure to LC
10 and LC
30 concentrations of fluxametamide significantly reduced the mean pupal weight of
P. xylostella in the F
0 and F
1 generations. This result was consistent with the previous report on sublethal doses of metaflumizone or spinetoram on
P. xylostella [
22,
23] and broflanilide on
Spodoptera litura [
53]. The reduction in pupal weight might be due to the decrease in food ingestion for optimal larval growth and the poor recovery efficiency of fluxametamide-treated larvae.
The adult male and female longevity of
P. xylostella F
0 generation were not significantly differed in fluxametamide treatments, but the adult longevity and total longevity of both males and females and the fecundity of gravid females were affected in the F
1 generation. Similar results were obtained by Yin et al. [
54], Su and Xia [
55] and Tamilselvan et al. [
23] who reported that sublethal concentrations of chlorantraniliprole, methylthio-diafenthiuron and spinetoram reduced the fecundity of
P. xylostella in F
1 progeny. We hypothesized that the production of viable female eggs and male sperms, required for normal fertilization, was less in the fluxametamide-treated
P. xylostella adults due to the cessation of feeding and lower food intake by the larval instars. The involvement of similar mechanisms in reduced fecundity has been reported for other lepidopteran pests such as
Platynota idaeusalis [
56] and
S. litura [
57]. Although the copulation and mating behavior of the adults that emerge after sublethal exposure is unknown, our findings indicate the potential for sublethal effects of fluxametamide on the reproductive physiology of
P. xylostella. On the other hand, it should be noted that endocrine signaling pathways regulate the fecundity of insects [
53]. Unlike the present study, Hu [
58] observed that two GABA-receptor-targeting insecticides, endosulfan and butane-fipronil, enhanced the fecundity of
Drosophila melanogaster by upregulation of the signaling pathway-related genes of juvenile hormone (
JHATT), ecdysone (
CYP307A1) and insulin-like (
ILP2,
ILP5 and
ILP7), and of the insulin-like (
ILP2 and
ILP5), respectively. In addition, the non-significant increase in the mean fecundity of
S. litura was observed at the sublethal concentrations of a meta-diamide insecticide broflanilide, but the exact reason has yet to be confirmed [
53]. However, neither any positive effect nor insecticide hormoligosis was reported in the fecundity of broflanilide-treated
Tetranychus urticae [
59], and this substantiated the present results. Besides, the sublethal effects of spinosad [
60] and spinetoram [
23] on the pre-oviposition period of
P. xylostella have been investigated. In the current study, exposure to LC
10 and LC
30 concentrations of fluxametamide also increased the TPOP of
P. xylostella F
1 generation, which is consistent with the previous authors.
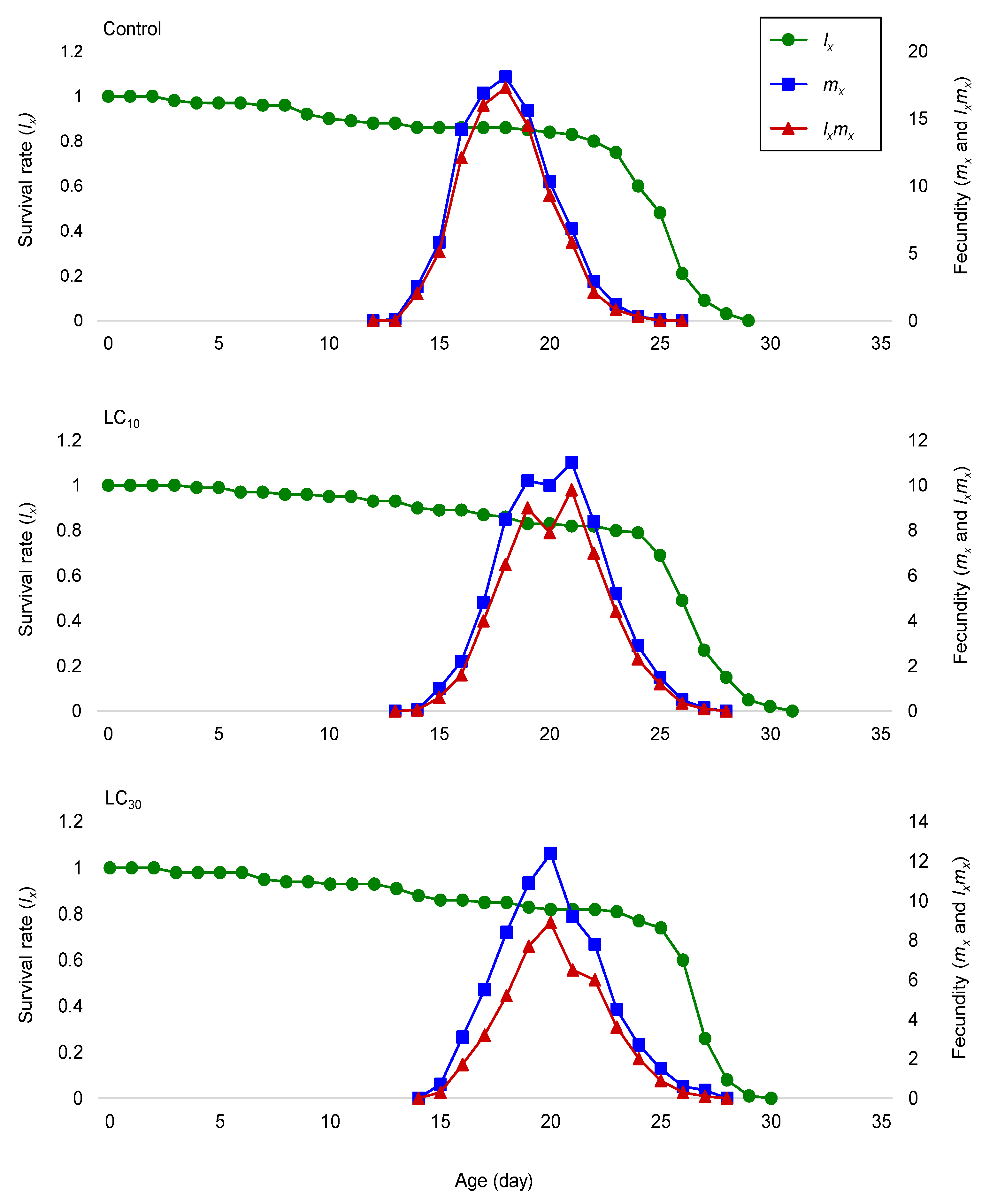
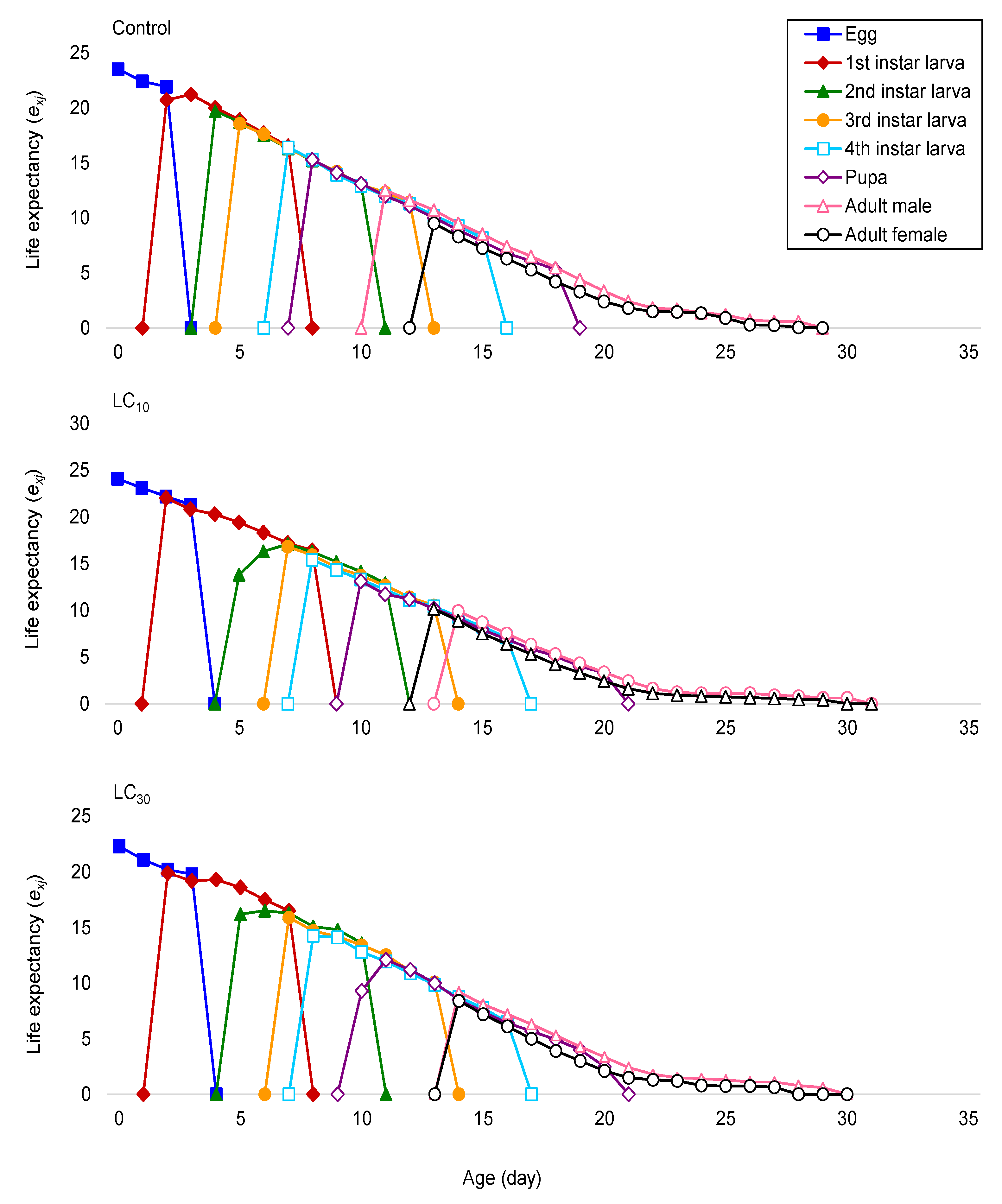
Demographic traits have been recommended as a more suitable approach for examining the total effects of an active ingredient on an insect population [
17,
61]. In addition, the effects of an insecticide on an insect population level could be predicted better by combining toxicological and ecological parameters. However, it is essential to study the effects of sublethal insecticide exposure on the fitness costs associated with insecticide resistance for the development of potential resistance management strategies [
62,
63]. In the present study, the mean values of
r,
λ and
R0 tended to be lower in the sublethal treatments of fluxametamide than in the control, while
T was significantly increased. The results indicate that the sublethal doses of fluxametamide can slow down the population dynamics of
P. xylostella through the reduction in their reproduction and survival. Furthermore, after the sublethal exposure, the significant changes in development times and fecundity caused a decrease in the
r and
R0 and resulted in a lower
Rf and longer
T, which was identical to the results seen with of
P. xylostella treated with chlorantraniliprole [
21], spinetoram [
23] and broflanilide [
24]. The population parameters of
P. xylostella, such as intrinsic rate of increase (
r), finite rate of increase (
λ), age-specific fecundity (
mx) and life expectancy (
exj) were also found to be lower in the sublethal exposure of indoxacarb [
64], pyriproxyfen [
65] and teflubenzuron [
66], and are consistent with the present findings.
The sublethal effects of an active ingredient on the fecundity and life stages of insects resulted from biochemical responses of various metabolic enzymes [
67,
68]. In the present study, the increased activities of GST may be contributing to the biochemical effects of the LC
30 treatment of fluxametamide in
P. xylostella. Based on the nature, dosage, and timing of chemical exposure, fluctuating activities of GST are possible in chemically stressed insects [
69,
70]. Similarly, in
Haemaphysalis longicornis and
Spodoptera litura, upregulation and expression of GST genes were detected after sublethal treatments with flumethrin [
71] and chlorpyrifos [
72], respectively, indicating a possible role in insect biochemical responses under sublethal insecticide exposure.


 ,
, 



{kind=link}
{kind=link}
{kind=link}