
Diet Composition Influences Growth Performance, Bioconversion of Black Soldier Fly Larvae: Agronomic Value and In Vitro Biofungicidal Activity of Derived Frass

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diet Preparation
2.2. Plant Pathogens
2.3. Rearing and Harvesting
2.4. Biochemical Analysis of Diets, Larvae, and Frass
2.5. Effect of Frass Extracts on Mycelial Growth of Plant Pathogens (Dual-Culture Overlay Assay)
2.6. Statistical Analyses
3. Results
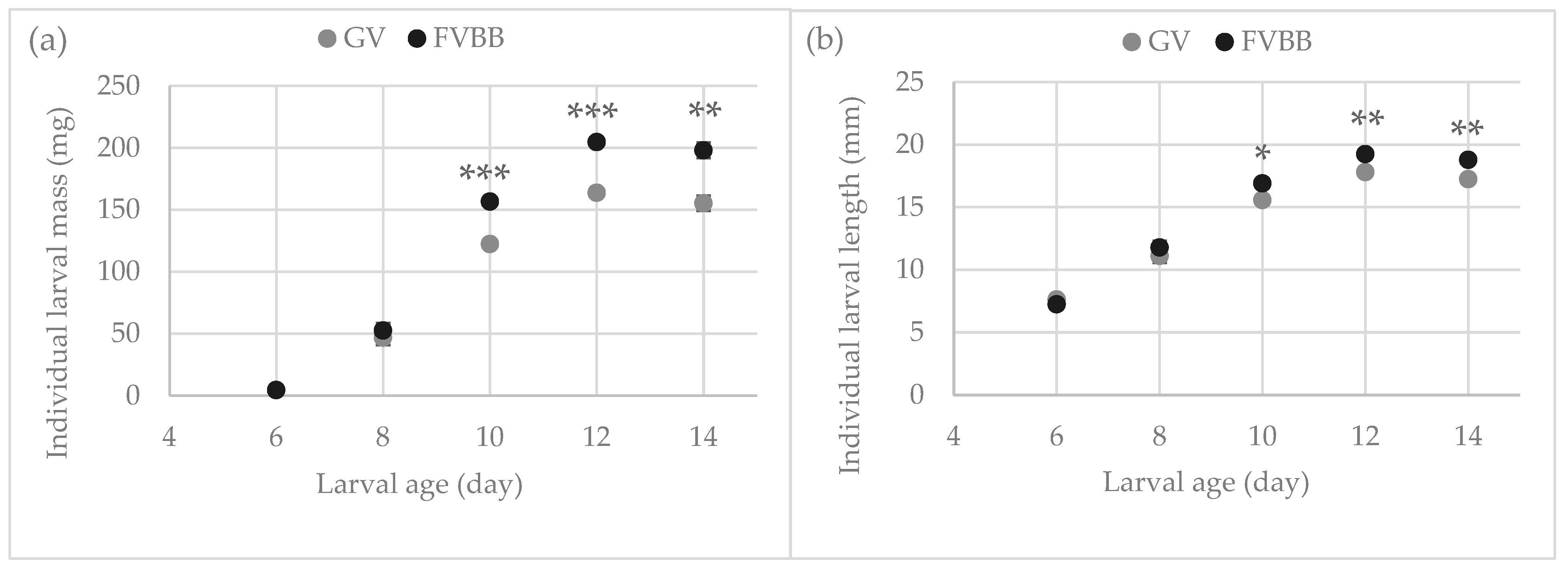
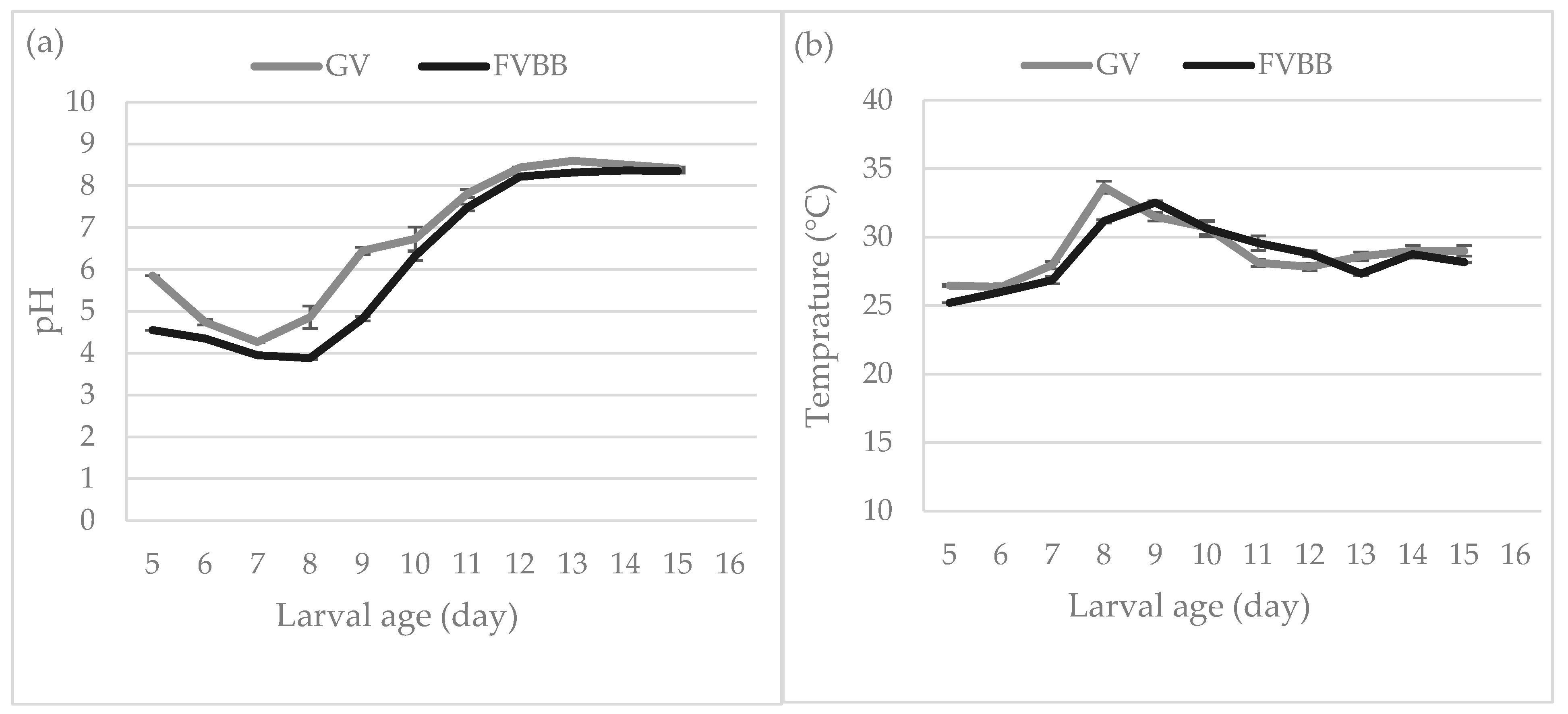
3.1. Larval Development
3.2. Chemical Composition of BSFL
3.3. Mineral Composition of Frass
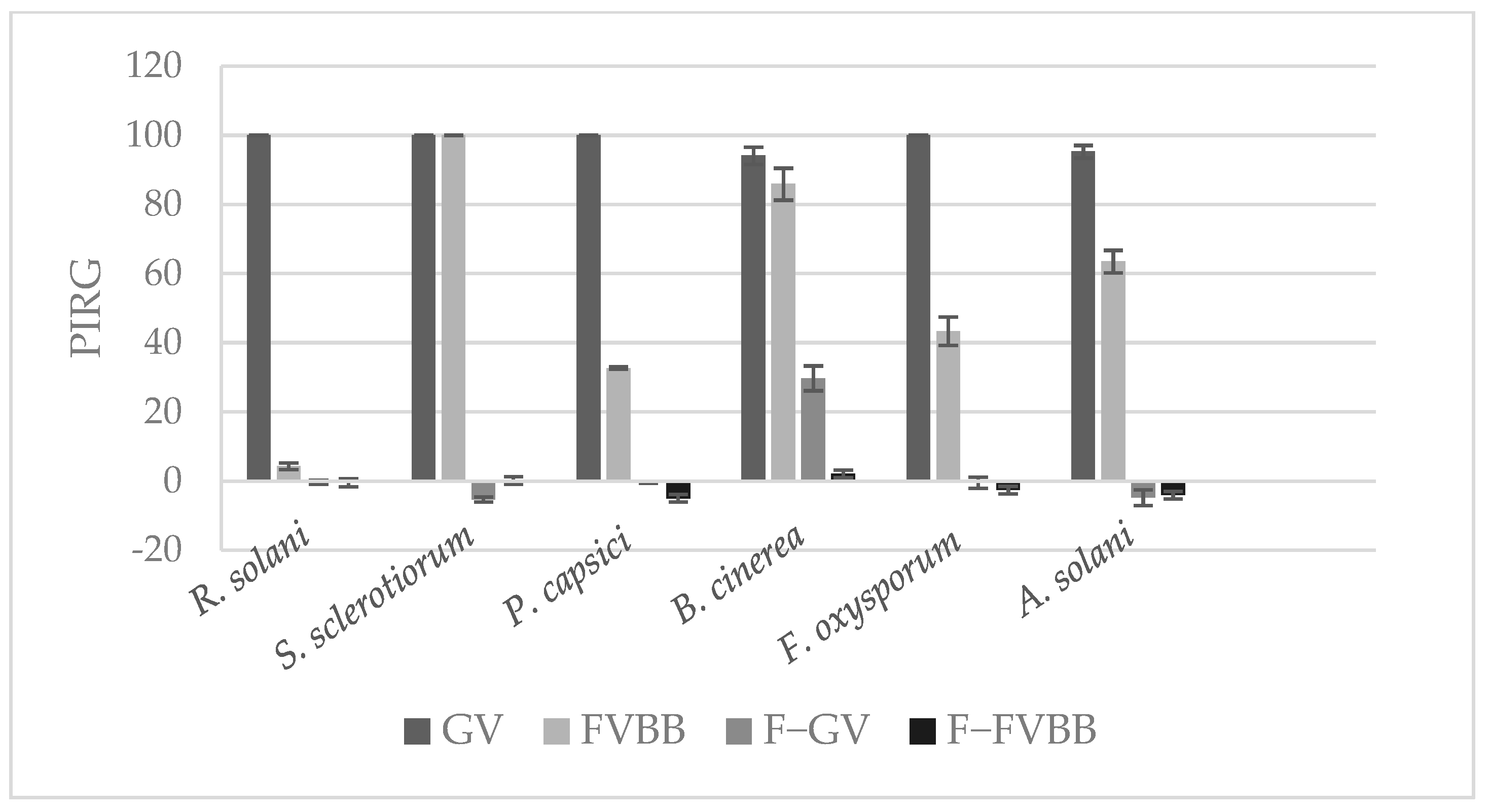
3.4. Effect of Frass Extracts on Mycelial Growth of Plant Pathogens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. Global Population Growth and Sustainable Development; Department of Economic and Social Affairs, Population Division: New York, NY, USA, 2021; p. 5. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). Technical Platform on the Measurement and Reduction of Food Loss and Waste; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2020. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). Global Food Losses and Food Waste–Extent, Causes and Prevention; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2011; p. 29. [Google Scholar]
- Melikoglu, M.; Lin, C.S.K.; Webb, C. Analysing Global Food Waste Problem: Pinpointing the Facts and Estimating the Energy Content. Cent. Eur. J. Eng. 2013, 3, 157–164. [Google Scholar] [CrossRef]
- Arsova, L. Anaerobic Digestion of Food Waste: Current Status, Problems and an Alternative Product. Master’s Thesis, Columbia University, New York, NY, USA, 2010. [Google Scholar]
- Lalander, C.; Diener, S.; Zurbrügg, C.; Vinnerås, B. Effects of Feedstock on Larval Development and Process Efficiency in Waste Treatment with Black Soldier Fly (Hermetia illucens). J. Clean. Prod. 2019, 208, 211–219. [Google Scholar] [CrossRef]
- Gold, M.; Cassar, C.M.; Zurbrügg, C.; Kreuzer, M.; Boulos, S.; Diener, S.; Mathys, A. Biowaste Treatment with Black Soldier Fly Larvae: Increasing Performance through the Formulation of Biowastes Based on Protein and Carbohydrates. Waste Manag. 2020, 102, 319–329. [Google Scholar] [CrossRef] [PubMed]
- van Huis, A. Manure and Flies: Biodegradation and/or Bioconversion? J. Insects Food Feed 2019, 5, 55–58. [Google Scholar] [CrossRef]
- Liu, T.; Awasthi, M.K.; Chen, H.; Duan, Y.; Awasthi, S.K.; Zhang, Z. Performance of Black Soldier Fly Larvae (Diptera: Stratiomyidae) for Manure Composting and Production of Cleaner Compost. J. Environ. Manag. 2019, 251, 109593. [Google Scholar] [CrossRef]
- Gao, Z.; Wang, W.; Lu, X.; Zhu, F.; Liu, W.; Wang, X.; Lei, C. Bioconversion Performance and Life Table of Black Soldier Fly (Hermetia illucens) on Fermented Maize Straw. J. Clean. Prod. 2019, 230, 974–980. [Google Scholar] [CrossRef]
- Koné, M. Étude de La Composition Des Matières Organiques Végétales Résiduelles Récoltées Chez Les Détaillants Alimentaires Sur Les Performances de Croissance, Les Bilans de Bioconversion et La Qualité Nutritionnelle Des Larves de Mouches Soldats Noires Destinées à l’alimentation Du Bétail. Master’s Thesis, Laval University, Québec City, QC, Canada, 2019. [Google Scholar]
- Lähteenmäki-Uutela, A.; Hénault-Ethier, L.; Marimuthu, S.B.; Talibov, S.; Allen, R.N.; Nemane, V.; Vandenberg, G.W.; Józefiak, D. The Impact of the Insect Regulatory System on the Insect Marketing System. J. Insects Food Feed 2018, 4, 187–198. [Google Scholar] [CrossRef]
- Staff, B. New Insect Protein Gains Approval for Use in Animal Feed: Regulatory Approval First of Its Kind in Canada; Marketwired: Langley, BC, Canada, 2016. [Google Scholar]
- Leung, V. Enterra Received CFIA Approval to Sell Insect Larvae to Aquaculture Industry; Enterra Feed Corporation: Langley, BC, Canada, 2017. [Google Scholar]
- Leung, V. Enterra Receives New Approvals to Sell Sustainable Insect Ingredients for Animal Feed in USA, Canada and EU; Enterra Feed Corporation: Langley, BC, Canada, 2018. [Google Scholar]
- Jucker, C.; Erba, D.; Leonardi, M.G.; Lupi, D.; Savoldelli, S. Assessment of Vegetable and Fruit Substrates as Potential Rearing Media for Hermetia illucens (Diptera: Stratiomyidae) Larvae. Environ. Entomol. 2017, 46, 1415–1423. [Google Scholar] [CrossRef]
- Parra Paz, A.S.; Carrejo, N.S.; Gómez Rodríguez, C.H. Effects of Larval Density and Feeding Rates on the Bioconversion of Vegetable Waste Using Black Soldier Fly Larvae Hermetia illucens (L.), (Diptera: Stratiomyidae). Waste Biomass Valoriz. 2015, 6, 1059–1065. [Google Scholar] [CrossRef]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black Soldier Fly Biowaste Treatment–Assessment of Global Warming Potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef]
- Diener, S.; Zurbrügg, C.; Tockner, K. Conversion of Organic Material by Black Soldier Fly Larvae: Establishing Optimal Feeding Rates. Waste Manag. Res. J. Sustain. Circ. Econ. 2009, 27, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Giannetto, A.; Oliva, S.; Ceccon Lanes, C.F.; de Araújo Pedron, F.; Savastano, D.; Baviera, C.; Parrino, V.; Lo Paro, G.; Spanò, N.C.; Cappello, T.; et al. Hermetia illucens (Diptera: Stratiomydae) Larvae and Prepupae: Biomass Production, Fatty Acid Profile and Expression of Key Genes Involved in Lipid Metabolism. J. Biotechnol. 2020, 307, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Chun, C.Y.; Yoong, L.S.; Kim, L.P.; Hock, T.L.; Ling, L.J. Comparison of Hermetia illucens Larvae and Pre-Pupae as Potential Aqua Feed Derived from the Biotransformation of Organic Waste. AIP Conf. Proc. 2019, 2157, 020008. [Google Scholar] [CrossRef]
- Wang, S.Y.; Wu, L.; Li, B.; Zhang, D. Reproductive Potential and Nutritional Composition of Hermetia illucens (Diptera: Stratiomyidae) Prepupae Reared on Different Organic Wastes. J. Econ. Entomol. 2020, 113, 527–537. [Google Scholar] [CrossRef]
- Jucker, C.; Lupi, D.; Moore, C.D.; Leonardi, M.G.; Savoldelli, S. Nutrient Recapture from Insect Farm Waste: Bioconversion with Hermetia illucens (L.) (Diptera: Stratiomyidae). Sustainability 2020, 12, 362. [Google Scholar] [CrossRef] [Green Version]
- Temple, W.D.; Radley, R.; Baker-French, J.; Richardson, F. Use of Enterra Natural Fertilizer (Black Soldier Fly Larvae Digestate) as a Soil Amendment; Enterra Feed Corporation: Vancouver, BC, Canada, 2013. [Google Scholar]
- Klammsteiner, T.; Turan, V.; Fernández-Delgado Juárez, M.; Oberegger, S.; Insam, H. Suitability of Black Soldier Fly Frass as Soil Amendment and Implication for Organic Waste Hygienization. Agronomy 2020, 10, 1578. [Google Scholar] [CrossRef]
- Beesigamukama, D.; Mochoge, B.; Korir, N.K.; Fiaboe, K.K.M.; Nakimbugwe, D.; Khamis, F.M.; Subramanian, S.; Dubois, T.; Musyoka, M.W.; Ekesi, S.; et al. Exploring Black Soldier Fly Frass as Novel Fertilizer for Improved Growth, Yield, and Nitrogen Use Efficiency of Maize Under Field Conditions. Front. Plant Sci. 2020, 11, 574592. [Google Scholar] [CrossRef]
- Beesigamukama, D.; Mochoge, B.; Korir, N.K.; Fiaboe, K.K.M.; Nakimbugwe, D.; Khamis, F.M.; Subramanian, S.; Wangu, M.M.; Dubois, T.; Ekesi, S.; et al. Low-Cost Technology for Recycling Agro-Industrial Waste into Nutrient-Rich Organic Fertilizer Using Black Soldier Fly. Waste Manag. 2021, 119, 183–194. [Google Scholar] [CrossRef]
- Setti, L.; Francia, E.; Pulvirenti, A.; Gigliano, S.; Zaccardelli, M.; Pane, C.; Caradonia, F.; Bortolini, S.; Maistrello, L.; Ronga, D. Use of Black Soldier Fly (Hermetia illucens (L.), Diptera: Stratiomyidae) Larvae Processing Residue in Peat-Based Growing Media. Waste Manag. 2019, 95, 278–288. [Google Scholar] [CrossRef]
- Shaviv, A. Advances in Controlled-Release Fertilizers. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2001; Volume 71, pp. 1–49. [Google Scholar]
- Penhallegon, R. Nitrogen-Phosphorus-Potassium Values of Organic Fertilizers; Oregon State University: Corvallis, OR, USA, 2003. [Google Scholar]
- Liu, T.; Awasthi, S.K.; Qin, S.; Liu, H.; Awasthi, M.K.; Zhou, Y.; Jiao, M.; Pandey, A.; Varjani, S.; Zhang, Z. Conversion Food Waste and Sawdust into Compost Employing Black Soldier Fly Larvae (Diptera: Stratiomyidae) under the Optimized Condition. Chemosphere 2021, 272, 129931. [Google Scholar] [CrossRef]
- Beesigamukama, D.; Mochoge, B.; Korir, N.; Musyoka, M.W.; Fiaboe, K.K.M.; Nakimbugwe, D.; Khamis, F.M.; Subramanian, S.; Dubois, T.; Ekesi, S.; et al. Nitrogen Fertilizer Equivalence of Black Soldier Fly Frass Fertilizer and Synchrony of Nitrogen Mineralization for Maize Production. Agronomy 2020, 10, 1395. [Google Scholar] [CrossRef]
- Fischer, H.; Romano, N. Fruit, Vegetable, and Starch Mixtures on the Nutritional Quality of Black Soldier Fly (Hermetia illucens) Larvae and Resulting Frass. J. Insects Food Feed 2021, 7, 319–327. [Google Scholar] [CrossRef]
- Beesigamukama, D.; Mochoge, B.; Korir, N.K.; Fiaboe, K.K.M.; Nakimbugwe, D.; Khamis, F.M.; Dubois, T.; Subramanian, S.; Wangu, M.M.; Ekesi, S.; et al. Biochar and Gypsum Amendment of Agro-Industrial Waste for Enhanced Black Soldier Fly Larval Biomass and Quality Frass Fertilizer. PLoS ONE 2020, 15, e0238154. [Google Scholar] [CrossRef]
- Quilliam, R.S.; Nuku-Adeku, C.; Maquart, P.; Little, D.; Newton, R.; Murray, F. Integrating Insect Frass Biofertilisers into Sustainable Peri-Urban Agro-Food Systems. J. Insects Food Feed 2020, 6, 315–322. [Google Scholar] [CrossRef]
- Arabzadeh, G.; Delisle-Houde, M.; Dorais, M.; Deschamps, M.-H.; Derome, N.; Tweddell, R.; Vandenberg, G. Evaluation of the Antifungal Activity of Black Soldier Fly Larval Frass Using the Double-Layer Agar Technique. In Proceedings of the Insects to Feed the World Conference, Quebec City, QC, Canada, 23–26 November 2020. [Google Scholar]
- Aktar, M.W.; Sengupta, D.; Chowdhury, A. Impact of Pesticides Use in Agriculture: Their Benefits and Hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hogsette, J.A. New Diets for Production of House Flies and Stable Flies (Diptera: Muscidae) in the Laboratory. J. Econ. Entomol. 1992, 85, 2291–2294. [Google Scholar] [CrossRef] [Green Version]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected Life-History Traits of Black Soldier Flies (Diptera: Stratiomyidae) Reared on Three Artificial Diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Nguyen, T.T.X.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of Black Soldier Fly (Diptera: Stratiomyidae) Larvae to Recycle Food Waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef]
- Bosch, G.; Oonincx, D.G.A.B.; Jordan, H.R.; Zhang, J.; van Loon, J.J.A.; van Huis, A.; Tomberlin, J.K. Standardisation of Quantitative Resource Conversion Studies with Black Soldier Fly Larvae. J. Insects Food Feed 2020, 6, 95–109. [Google Scholar] [CrossRef]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of Rearing Substrate on Growth Performance, Waste Reduction Efficiency and Chemical Composition of Black Soldier Fly (Hermetia illucens) Larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Official Methods of Analysis of AOAC International. AOAC Official Method 2001.11; AOAC international: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio Molitor, Alphitobius Diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Official Methods of Analysis of AOAC International. AOAC Official Method 942.05; AOAC international: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Official Methods of Analysis of AOAC International. AOAC Official Method 973.18; AOAC international: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Hall, M.B. Determination of Starch, Including Maltooligosaccharides, in Animal Feeds: Comparison of Methods and a Method Recommended for AOAC Collaborative Study. J. AOAC Int. 2009, 92, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Official Methods of Analysis of AOAC International. AOAC Official Method 2003.06; AOAC international: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Spinelli, J.; Lehman, L.; Wieg, D. Composition, Processing, and Utilization of Red Crab (Pleuroncodes Planipes) as an Aquacultural Feed Ingredient. J. Fish. Res. Board Can. 1974, 31, 1025–1029. [Google Scholar] [CrossRef]
- Isaac, R.A.; Johnson, W.C. Determination of Total Nitrogen in Plant Tissue, Using a Block Digestor. J. Assoc. Off. Anal. Chem. 1976, 59, 98–100. [Google Scholar] [CrossRef]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant Tissue by Nitration of Salicylic Acid. Commun. Soil Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Conseil des Productions Végétales du Québec. Méthodes d’Analyse Des Sols, Des Fumiers et Des Tissus Végétaux., Méthode FO-1; Conseil des Productions Végétales du Québec: Quebec City, QC, Canada, 1988. [Google Scholar]
- Conseil des Productions Végétales du Québec. Méthodes d’Analyse Des Sols, Des Fumiers et Des Tissus Végétaux., Méthode MA-2; Conseil des Productions Végétales du Québec: Quebec City, QC, Canada, 1988. [Google Scholar]
- Conseil des Productions Végétales du Québec. Méthodes d’Analyse Des Sols, Des Fumiers et Des Tissus Végétaux., Méthode MA-1; Conseil des Productions Végétales du Québec: Quebec City, QC, Canada, 1988. [Google Scholar]
- Reidinger, S.; Ramsey, M.H.; Hartley, S.E. Rapid and Accurate Analyses of Silicon and Phosphorus in Plants Using a Portable X-ray Fluorescence Spectrometer. New Phytol. 2012, 195, 699–706. [Google Scholar] [CrossRef]
- Marguí, E.; Queralt, I.; Hidalgo, M. Application of X-ray Fluorescence Spectrometry to Determination and Quantitation of Metals in Vegetal Material. TrAC Trends Anal. Chem. 2009, 28, 362–372. [Google Scholar] [CrossRef]
- Surendra, K.C.; Tomberlin, J.K.; van Huis, A.; Cammack, J.A.; Heckmann, L.-H.L.; Khanal, S.K. Rethinking Organic Wastes Bioconversion: Evaluating the Potential of the Black Soldier Fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae) (BSF). Waste Manag. 2020, 117, 58–80. [Google Scholar] [CrossRef]
- Mutafela, R.N. High Value Organic Waste Treatment via Black Soldier Fly Bioconversion: Onsite Pilot Study. Master’s Thesis, KTH Royal Institute of Technology, Stockholm, Sweden, 2015. [Google Scholar]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional Composition of Black Soldier Fly (Hermetia illucens) Prepupae Reared on Different Organic Waste Substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Hard, J.J.; Bradshaw, W.E.; Malarkey, D.J. Resource- and Density-Dependent Development in Tree-Hole Mosquitoes. Oikos 1989, 54, 137–144. [Google Scholar] [CrossRef]
- Cammack, J.A.; Tomberlin, J.K. The Impact of Diet Protein and Carbohydrate on Select Life-History Traits of The Black Soldier Fly Hermetia illucens (L.) (Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Influence of Larval Density and Dietary Nutrient Concentration on Performance, Body Protein, and Fat Contents of Black Soldier Fly Larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Barragán-Fonseca, K.; Pineda-Mejia, J.; Dicke, M.; van Loon, J.J.A. Performance of the Black Soldier Fly (Diptera: Stratiomyidae) on Vegetable Residue-Based Diets Formulated Based on Protein and Carbohydrate Contents. J. Econ. Entomol. 2018, 111, 2676–2683. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; Broekhoven, S.V.; Huis, A.V.; Loon, J.J.A. van Feed Conversion, Survival and Development, and Composition of Four Insect Species on Diets Composed of Food By-Products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barragán-Fonseca, K.B. Flies Are What They Eat. Ph.D. Thesis, Wageningen University & Research, Wageningen, The Netherlands, 2018. [Google Scholar]
- Gold, M.; Tomberlin, J.K.; Diener, S.; Zurbrügg, C.; Mathys, A. Decomposition of Biowaste Macronutrients, Microbes, and Chemicals in Black Soldier Fly Larval Treatment: A Review. Waste Manag. 2018, 82, 302–318. [Google Scholar] [CrossRef]
- Lee, C.-M.; Lee, Y.-S.; Seo, S.-H.; Yoon, S.-H.; Kim, S.-J.; Hahn, B.-S.; Sim, J.-S.; Koo, B.-S. Screening and Characterization of a Novel Cellulase Gene from the Gut Microflora of Hermetia illucens Using Metagenomic Library. J. Microbiol. Biotechnol. 2014, 24, 1196–1206. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.; Park, S.; Choi, J.; Jeong, G.; Lee, S.-B.; Choi, Y.; Lee, S.-J. The Intestinal Bacterial Community in the Food Waste-Reducing Larvae of Hermetia illucens. Curr. Microbiol. 2011, 62, 1390–1399. [Google Scholar] [CrossRef] [PubMed]
- Beniers, J.J.A.; Graham, R.I. Effect of Protein and Carbohydrate Feed Concentrations on the Growth and Composition of Black Soldier Fly (Hermetia illucens) Larvae. J. Insects Food Feed 2019, 5, 193–199. [Google Scholar] [CrossRef]
- Bonelli, M.; Bruno, D.; Caccia, S.; Sgambetterra, G.; Cappellozza, S.; Jucker, C.; Tettamanti, G.; Casartelli, M. Structural and Functional Characterization of Hermetia illucens Larval Midgut. Front. Physiol. 2019, 10, 204. [Google Scholar] [CrossRef]
- Sealey, W.M.; Gaylord, T.G.; Barrows, F.T.; Tomberlin, J.K.; McGuire, M.A.; Ross, C.; St-Hilaire, S. Sensory Analysis of Rainbow Trout, Oncorhynchus Mykiss, Fed Enriched Black Soldier Fly Prepupae, Hermetia illucens. J. World Aquac. Soc. 2011, 42, 34–45. [Google Scholar] [CrossRef]
- Liu, X.; Chen, X.; Wang, H.; Yang, Q.; Rehman, K.U.; Li, W.; Cai, M.; Li, Q.; Mazza, L.; Zhang, J.; et al. Dynamic Changes of Nutrient Composition throughout the Entire Life Cycle of Black Soldier Fly. PLoS ONE 2017, 12, e0182601. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; ur Rehman, K.; Yu, Y.; Liu, X.; Wang, H.; Tomberlin, J.K.; Sze, S.-H.; Cai, M.; Zhang, J.; Yu, Z.; et al. De Novo Transcriptome Sequencing and Analysis Revealed the Molecular Basis of Rapid Fat Accumulation by Black Soldier Fly (Hermetia illucens, L.) for Development of Insectival Biodiesel. Biotechnol. Biofuels 2019, 12, 194. [Google Scholar] [CrossRef]
- Inagaki, S.; Yamashita, O. Metabolic Shift from Lipogenesis to Glycogenesis in the Last Instar Larval Fat Body of the Silkworm, Bombyx Mori. Insect Biochem. 1986, 16, 327–331. [Google Scholar] [CrossRef]
- DOWNER, R.G.H.; MATTHEWS, J.R. Patterns of Lipid Distribution and Utilisation in Insects. Am. Zool. 1976, 16, 733–745. [Google Scholar] [CrossRef]
- Tschirner, M.; Simon, A. Influence of Different Growing Substrates and Processing on the Nutrient Composition of Black Soldier Fly Larvae Destined for Animal Feed. J. Insects Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Parodi, A.; De Boer, I.J.M.; Gerrits, W.J.J.; Van Loon, J.J.A.; Heetkamp, M.J.W.; Van Schelt, J.; Bolhuis, J.E.; Van Zanten, H.H.E. Bioconversion Efficiencies, Greenhouse Gas and Ammonia Emissions during Black Soldier Fly Rearing–A Mass Balance Approach. J. Clean. Prod. 2020, 271, 122488. [Google Scholar] [CrossRef]
- Caruso, D.; Devic, E.; Subamia, I.W.; Talamond, P.; Baras, E. Technical Handbook of Domestication and Production of Diptera Black Soldier Fly (BSF) Hermetia illucens, Stratiomyidae; PT Penerbit IPB Press: Bogor, Indonesia, 2014. [Google Scholar]
- Kawasaki, K.; Kawasaki, T.; Hirayasu, H.; Matsumoto, Y.; Fujitani, Y. Evaluation of Fertilizer Value of Residues Obtained after Processing Household Organic Waste with Black Soldier Fly Larvae (Hermetia illucens). Sustainability 2020, 12, 4920. [Google Scholar] [CrossRef]
- Watson, C.; Schlösser, C.; Vögerl, J.; Wichern, F. Excellent Excrement? Frass Impacts on a Soil’s Microbial Community, Processes and Metal Bioavailability. Appl. Soil Ecol. 2021, 168, 104110. [Google Scholar] [CrossRef]
- Green, T.R.; Popa, R. Enhanced Ammonia Content in Compost Leachate Processed by Black Soldier Fly Larvae. Appl. Biochem. Biotechnol. 2012, 166, 1381–1387. [Google Scholar] [CrossRef]
- Awasthi, M.K.; Wang, Q.; Chen, H.; Wang, M.; Ren, X.; Zhao, J.; Li, J.; Guo, D.; Li, D.-S.; Awasthi, S.K.; et al. Evaluation of Biochar Amended Biosolids Co-Composting to Improve the Nutrient Transformation and Its Correlation as a Function for the Production of Nutrient-Rich Compost. Bioresour. Technol. 2017, 237, 156–166. [Google Scholar] [CrossRef]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of Animal Manures and Chemical Criteria for Compost Maturity Assessment. A Review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef]
- Sarpong, D.; Oduro-Kwarteng, S.; Gyasi, S.F.; Buamah, R.; Donkor, E.; Awuah, E.; Baah, M.K. Biodegradation by Composting of Municipal Organic Solid Waste into Organic Fertilizer Using the Black Soldier Fly (Hermetia illucens) (Diptera: Stratiomyidae) Larvae. Int. J. Recycl. Org. Waste Agric. 2019, 8, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Lin-ki, J.W.C. Drift of Black-Fly Larvae and the Infuence of Water Velocity, Substrate Roughness and Incident Light Intensity on Their Microdistribution (Diptera: Simuliidae). Ph.D. Thesis, MCMaster University, Hamilton, ON, Canada, 1975. [Google Scholar]
- Chang, R.; Yao, Y.; Cao, W.; Wang, J.; Wang, X.; Chen, Q. Effects of Composting and Carbon Based Materials on Carbon and Nitrogen Loss in the Arable Land Utilization of Cow Manure and Corn Stalks. J. Environ. Manage. 2019, 233, 283–290. [Google Scholar] [CrossRef]
- Anyega, A.O.; Korir, N.K.; Beesigamukama, D.; Changeh, G.J.; Nkoba, K.; Subramanian, S.; van Loon, J.J.A.; Dicke, M.; Tanga, C.M. Black Soldier Fly-Composted Organic Fertilizer Enhances Growth, Yield, and Nutrient Quality of Three Key Vegetable Crops in Sub-Saharan Africa. Front. Plant Sci. 2021, 12, 680312. [Google Scholar] [CrossRef]
- Song, S.; Ee, A.W.L.; Tan, J.K.N.; Cheong, J.C.; Chiam, Z.; Arora, S.; Lam, W.N.; Tan, H.T.W. Upcycling Food Waste Using Black Soldier Fly Larvae: Effects of Further Composting on Frass Quality, Fertilising Effect and Its Global Warming Potential. J. Clean. Prod. 2021, 288, 125664. [Google Scholar] [CrossRef]
- Lim, S.-S.; Park, H.-J.; Hao, X.; Lee, S.-I.; Jeon, B.-J.; Kwak, J.-H.; Choi, W.-J. Nitrogen, Carbon, and Dry Matter Losses during Composting of Livestock Manure with Two Bulking Agents as Affected by Co-Amendments of Phosphogypsum and Zeolite. Ecol. Eng. 2017, 102, 280–290. [Google Scholar] [CrossRef]
- Dioha, I.J.; Ikeme, C.H.; Nafi’u, T.; Soba, N.I.; Yusuf, M.B.S. Effect of Carbon to Nitrogen Ratio on Biogas Production. Int. Res. J. Nat. Sci. 2013, 1, 1–10. [Google Scholar]
- Wang, L.; Li, Y.; Prasher, S.O.; Yan, B.; Ou, Y.; Cui, H.; Cui, Y. Organic Matter, a Critical Factor to Immobilize Phosphorus, Copper, and Zinc during Composting under Various Initial C/N Ratios. Bioresour. Technol. 2019, 289, 121745. [Google Scholar] [CrossRef]
- Goyal, S.; Dhull, S.K.; Kapoor, K.K. Chemical and Biological Changes during Composting of Different Organic Wastes and Assessment of Compost Maturity. Bioresour. Technol. 2005, 96, 1584–1591. [Google Scholar] [CrossRef]
- Pan, I.; Dam, B.; Sen, S.K. Composting of Common Organic Wastes Using Microbial Inoculants. 3 Biotech 2012, 2, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Dorais, M. Chapter 05-Advances in Organic Greenhouse Cultivation. In Achieving Sustainable Greenhouse Cultivation; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 121–176. ISBN 978-0-429-26674-4. [Google Scholar]
- Dorais, M.; Alsanius, B. Advances and Trends in Organic Fruit and Vegetable Farming Research. In Horticultural Reviews: Volume 43; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 185–268. ISBN 978-1-119-10778-1. [Google Scholar]
- de Bertoldi, M.; Vallini, G.; Pera, A. The Biology of Composting: A Review. Waste Manag. Res. 1983, 1, 157–176. [Google Scholar] [CrossRef]
- ur Rehman, K.; Rehman, A.; Cai, M.; Zheng, L.; Xiao, X.; Somroo, A.A.; Wang, H.; Li, W.; Yu, Z.; Zhang, J. Conversion of Mixtures of Dairy Manure and Soybean Curd Residue by Black Soldier Fly Larvae (Hermetia illucens L.). J. Clean. Prod. 2017, 154, 366–373. [Google Scholar] [CrossRef]
- Meneguz, M.; Gasco, L.; Tomberlin, J.K. Impact of PH and Feeding System on Black Soldier Fly (Hermetia illucens, L.; Diptera: Stratiomyidae) Larval Development. PLoS ONE 2018, 13, e0202591. [Google Scholar] [CrossRef]
- Lemaire, F.; Dartigues, A.; Rivière, L.M. Properties of Substrate Made with Spent Mushroom Compost. Acta Hortic. 1985, 172, 13–30. [Google Scholar] [CrossRef]
- Noguera, P.; Abad, M.; Puchades, R.; Maquieira, A.; Noguera, V. Influence of Particle Size on Physical and Chemical Properties of Coconut Coir Dust as Container Medium. Commun. Soil Sci. Plant Anal. 2003, 34, 593–605. [Google Scholar] [CrossRef]
- Wu, X.; Cai, R.; Wang, X.; Wu, N.; Xu, X. Study on Effects of Black Soldier Fly Feces on Rice Growth. IOP Conf. Ser. Earth Environ. Sci. 2020, 450, 012099. [Google Scholar] [CrossRef]
- Chiam, Z.; Lee, J.T.E.; Tan, J.K.N.; Song, S.; Arora, S.; Tong, Y.W.; Tan, H.T.W. Evaluating the Potential of Okara-Derived Black Soldier Fly Larval Frass as a Soil Amendment. J. Environ. Manag. 2021, 286, 112163. [Google Scholar] [CrossRef]
- Joshi, R.; Singh, J.; Vig, A.P. Vermicompost as an Effective Organic Fertilizer and Biocontrol Agent: Effect on Growth, Yield and Quality of Plants. Rev. Environ. Sci. Biotechnol. 2015, 14, 137–159. [Google Scholar] [CrossRef]
- St. Martin, C.C.G.; Brathwaite, R.A.I. Compost and Compost Tea: Principles and Prospects as Substrates and Soil-Borne Disease Management Strategies in Soil-Less Vegetable Production. Biol. Agric. Hortic. 2012, 28, 1–33. [Google Scholar] [CrossRef]
- Ibrahim, H.A.K.; El-Fiki, I.A.I. Study on the Effect of Yeast in Compost Tea Efficiency in Controlling Chocolate Leaf Spot Disease in Broad Bean (Vicia Faba). Org. Agric. 2019, 9, 175–188. [Google Scholar] [CrossRef]
- Yasir, M.; Aslam, Z.; Kim, S.W.; Lee, S.-W.; Jeon, C.O.; Chung, Y.R. Bacterial Community Composition and Chitinase Gene Diversity of Vermicompost with Antifungal Activity. Bioresour. Technol. 2009, 100, 4396–4403. [Google Scholar] [CrossRef] [PubMed]
- Manandhar, T.; Yami, K.D. Biological Control of Foot Rot Disease of Rice Using Fermented Products of Compost and Vermicompost. Sci. World 2008, 6, 52–57. [Google Scholar] [CrossRef]
- Simsek Ersahin, Y.; Haktanir, K.; Yanar, Y. Vermicompost Suppresses Rhizoctonia Solani Kühn in Cucumber Seedlings. J. Plant Dis. Prot. 2009, 116, 182–188. [Google Scholar] [CrossRef]
- Pane, C.; Spaccini, R.; Piccolo, A.; Scala, F.; Bonanomi, G. Compost Amendments Enhance Peat Suppressiveness to Pythium Ultimum, Rhizoctonia Solani and Sclerotinia Minor. Biol. Control 2011, 56, 115–124. [Google Scholar] [CrossRef]
- Scheuerell, S.J.; Mahaffee, W.F. Compost Tea as a Container Medium Drench for Suppressing Seedling Damping-Off Caused by Pythium Ultimum. Phytopathology 2004, 94, 1156–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samet, M.; Charfeddine, M.; Kamoun, L.; Nouri-Ellouze, O.; Gargouri-Bouzid, R. Effect of Compost Tea Containing Phosphogypsum on Potato Plant Growth and Protection against Fusarium Solani Infection. Environ. Sci. Pollut. Res. Int. 2018, 25, 18921–18937. [Google Scholar] [CrossRef]
- On, A.; Wong, F.; Ko, Q.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Antifungal Effects of Compost Tea Microorganisms on Tomato Pathogens. Biol. Control 2015, 80, 63–69. [Google Scholar] [CrossRef]
- Pane, C.; Celano, G.; Villecco, D.; Zaccardelli, M. Control of Botrytis Cinerea, Alternaria Alternata and Pyrenochaeta Lycopersici on Tomato with Whey Compost-Tea Applications. Crop Prot. 2012, 38, 80–86. [Google Scholar] [CrossRef]
- Koné, S.B.; Dionne, A.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Suppressive Effect of Non-Aerated Compost Teas on Foliar Fungal Pathogens of Tomato. Biol. Control 2010, 52, 167–173. [Google Scholar] [CrossRef]
- Dionne, A.; Tweddell, R.J.; Antoun, H.; Avis, T.J. Effect of Non-Aerated Compost Teas on Damping-off Pathogens of Tomato. Can. J. Plant Pathol. 2012, 34, 51–57. [Google Scholar] [CrossRef]
- Arabzadeh, G.; Delisle-Houde, M.; Nguyen, T.T.A.; Tweddell, R.; Dorais, M.; Deschamps, M.-H.; Vandenberg, G. Evaluation of the Antifungal Activity of Black Soldier Fly Larval Frass Using the Double-Layer Agar Technique. In Proceedings of the Insects to Feed the World Conference, Quebec City, QC, Canada, 12 June 2022. [Google Scholar]
- Brakspear, C. Biocontrol Potential of Black Soldier Fly (Hermetia illucens L.) Frass against Fusarium and Verticillium Wilt of Tomato. In Proceedings of the Insects to Feed the World Conference, Quebec City, QC, Canada, 12 June 2022. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Parameter | GV | FVBB | p-Value |
|---|---|---|---|
| DM (%) | 29.94 ± 0.31 a | 30.27 ± 0.44 | 0.57 |
| Crude protein (% of DM) | 12.46 ± 0.08 | 19.66 ± 0.11 | <0.001 |
| Crude lipid (% of DM) | 3.29 ± 1.21 | 8.26 ± 1.01 | <0.001 |
| Crude ash (% of DM) | 4.98 ± 0.05 | 4.16 ± 0.00 | <0.001 |
| Starch (% of DM) | 25.52 ± 0.11 | 19.93 ± 0.20 | 0.004 |
| Sucrose (% of DM) | 2.85 ± 0.03 | 6.98 ± 0.03 | <0.001 |
| Glucose (% of DM) | 0.58 ± 0.01 | 7.22 ± 0.03 | <0.001 |
| Galactose (% of DM) | 0.61 ± 0.00 | 0.00 ± 0.00 | 0.000 |
| Fructose (% of DM) | 0.52 ± 0.01 | 7.20 ± 0.03 | <0.001 |
| Cellulose (% of DM) | 10.15 ± 1.04 | 5.66 ± 0.51 | 0.018 |
| Hemicellulose (% of DM) | 16.13 ± 0.64 | 8.87 ± 0.33 | <0.001 |
| Energy (cal g−1) | 3950.15 ± 15.09 | 4373.71 ± 4.19 | <0.001 |
| Organic C (%) | 54.72 ± 0.04 | 55.26 ± 0.01 | 0.004 |
| N (%) | 2.24 ± 0.04 | 3.28 ± 0.04 | <0.001 |
| P (%) | 0.45 ± 0.01 | 0.54 ± 0.00 | 0.003 |
| K (%) | 1.33 ± 0.03 | 1.14 ± 0.01 | 0.013 |
| C:N | 24.46 ± 0.38 | 16.76 ± 0.18 | <0.001 |
| Parameter | GV | FVBB | p-Value |
|---|---|---|---|
| Larval biomass (g as is) | 98.77 ± 3.50 a | 114.7 ± 1.22 | 0.034 |
| Larval biomass (g of DM) | 24.82 ± 0.62 | 32.68 ± 0.47 | <0.001 |
| Individual larval mass (mg as is) | 129.31 ± 2.45 | 162.11 ± 2.22 | <0.001 |
| Individual larval mass (mg of DM) | 32.52 ± 0.58 | 46.18 ± 0.54 | <0.001 |
| Bioconversion rate (%) | 10.05 ± 0.27 | 13.32 ± 0.20 | <0.001 |
| Substrate reduction rate (%) | 70.48 ± 0.85 | 67.76 ± 0.75 | 0.074 |
| Mortality (%) | 4.58 ± 1.57 | 11.5 ± 2.13 | 0.059 |
| BER b | 14.26 ± 0.44 | 19.65 ± 0.09 | <0.001 |
| Protein conversion rate (%) | 42.88 ± 0.89 | 32.39 ± 0.76 | <0.001 |
| Parameter (%) | GV | FVBB | p-Value |
|---|---|---|---|
| Crude protein (6.25) | 51.66 ± 0.24 a | 46.75 ± 0.55 | 0.0012 |
| Crude protein (4.67) | 38.60 ± 0.18 | 34.89 ± 0.41 | 0.0012 |
| Crude lipid | 17.26 ± 1.81 | 35.99 ± 1.02 | <0.001 |
| Crude ash | 9.60 ± 0.09 | 4.22 ± 0.18 | <0.001 |
| Dry matter | 25.17 ± 0.69 | 28.49 ± 0.17 | 0.009 |
| Chitin | 6.92 ± 0.13 | 6.81 ± 0.23 | 0.654 |
| Parameter | GV Frass | FVBB Frass | p-Value |
|---|---|---|---|
| DM (%) | 22.09 ± 0.62 a | 25.57 ± 0.23 | 0.02 |
| OM (%) | 87.50 ± 0.18 | 88.67 ± 0.34 | 0.029 |
| Ash (%) | 11.86 ± 0.16 | 11.05 ± 0.31 | 0.099 |
| Organic C (%) | 50.76 ± 0.10 | 51.44 ± 0.20 | 0.030 |
| N (%) | 2.01 ± 0.08 | 2.70 ± 0.01 | 0.006 |
| NO3 (%) | <0.02 | <0.02 | |
| NH4 (%) | 0.17 ± 0.00 | 0.78 ± 0.02 | 0.002 |
| Mg (%) | 0.82 ± 0.02 | 0.42 ± 0.02 | <0.001 |
| P (%) | 1.94 ± 0.03 | 1.27 ± 0.01 | <0.001 |
| K (%) | 3.65 ± 0.01 | 2.97 ± 0.05 | 0.003 |
| Ca (%) | 0.20 ± 0.01 | 0.04 ± 0.00 | 0.002 |
| S (%) | 0.46 ± 0.00 | 1.35 ± 0.01 | <0.001 |
| C:N | 25.36 ± 1.10 | 19.10 ± 0.13 | 0.022 |
| EC (mS cm−1) | 5.35 ± 0.13 | 10.21 ± 0.11 | <0.001 |
| Zn (mg Kg−1) | 99.18 ± 0.92 | 57.32 ± 1.25 | <0.001 |
| Cu (mg Kg−1) | 18.02 ± 1.57 | <9.00 | |
| Fe (mg Kg−1) | 285.74 ± 9.08 | 150.45 ± 5.54 | 0.0013 |
| Mn (mg Kg−1) | 31.33 ± 6.95 | <17.00 | |
| Na (%) | 0.07 ± 0.00 | 0.97 ± 0.05 | 0.0017 |
| Cl (%) | 0.17 ± 0.00 | 1.53 ± 0.03 | <0.001 |
| Mo (mg Kg−1) | <4.00 | <4.00 | |
| Sr (mg Kg−1) | 13.30 ± 0.61 | 5.56 ± 0.67 | <0.001 |
| Rb (mg Kg−1) | 23.41 ± 0.18 | 23.73 ± 0.57 | 0.952 |
| Pb (mg Kg−1) | <1.00 | <1.00 | |
| Se (mg Kg−1) | <2.00 | <2.00 | |
| As (mg Kg−1) | <2.00 | <2.00 | |
| Hg (mg Kg−1) | <3.00 | <3.00 | |
| Si (%) | 0.26 ± 0.00 | 0.05 ± 0.00 | <0.001 |
| Al (%) | <0.17 | <0.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arabzadeh, G.; Delisle-Houde, M.; Tweddell, R.J.; Deschamps, M.-H.; Dorais, M.; Lebeuf, Y.; Derome, N.; Vandenberg, G. Diet Composition Influences Growth Performance, Bioconversion of Black Soldier Fly Larvae: Agronomic Value and In Vitro Biofungicidal Activity of Derived Frass. Agronomy 2022, 12, 1765. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081765
Arabzadeh G, Delisle-Houde M, Tweddell RJ, Deschamps M-H, Dorais M, Lebeuf Y, Derome N, Vandenberg G. Diet Composition Influences Growth Performance, Bioconversion of Black Soldier Fly Larvae: Agronomic Value and In Vitro Biofungicidal Activity of Derived Frass. Agronomy. 2022; 12(8):1765. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081765
Chicago/Turabian StyleArabzadeh, Ghazaleh, Maxime Delisle-Houde, Russell J. Tweddell, Marie-Hélène Deschamps, Martine Dorais, Yolaine Lebeuf, Nicolas Derome, and Grant Vandenberg. 2022. "Diet Composition Influences Growth Performance, Bioconversion of Black Soldier Fly Larvae: Agronomic Value and In Vitro Biofungicidal Activity of Derived Frass" Agronomy 12, no. 8: 1765. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081765