
Sulla Powdery Mildew: Phylogeny and Host Range



Institute for Sustainable Agriculture, CSIC, Avda. Menéndez Pidal s/n, 14004 Córdoba, Spain
*
Author to whom correspondence should be addressed.
†
The author has passed away.
Agronomy 2022, 12(8), 1852; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081852
Submission received: 16 June 2022
/
Revised: 31 July 2022
/
Accepted: 31 July 2022
/
Published: 4 August 2022
(This article belongs to the Special Issue Crop Powdery Mildew)
Abstract
:Sulla is a biannual forage legume cultivated throughout the Mediterranean Basin. It can be severely damaged by powdery mildew, but there is little understanding on its causal species or its host range. The taxonomic characterization of Erysiphe species is mainly based on the examination of chasmothecia morphology, or on the molecular analysis of ITS1 and ITS2 sequences. However, a description of chasmothecia morphology or ITS sequences is not always available to clearly assign a given isolate to an existing or novel Erysiphe species. In an attempt to clarify the identity of the powdery mildew infecting sulla crop we studied the morphology and ITS of nine populations collected over years and countries and compared them with available ITS sequences. Phylogenetic analysis showed that the powdery mildews collected on sulla clustered together with isolates collected on Coronilla varia in an independent clade between the E. guarinonii/E. trifoliorum and E. palczewskii clades, strongly suggesting that all these sequences correspond to the same Erysiphe species. Measurements of conidia and conidial foot cells of the sulla-infecting powdery mildews partially overlap those of other Erysiphe species, although they clearly differentiate from them, supporting the assumption that powdery mildews from sulla belong to a distinct species as observed from the phylogenetic analysis. As far as we know, our study is the first to report the molecular characterization of powdery mildew isolated on plants from the Hedysarum clade. Cross-inoculations confirmed a high specialization of the powdery mildew of sulla, with the sulla’s isolates infecting only the S. coronaria accessions, and none of the accessions from the other legume genera studied. All studied S. coronaria accessions were heavily infected by the sulla isolate but not by any other isolate used. All this points to E. hedysari as the causal agent of the S. coronaria powdery mildew.
1. Introduction
Sulla (Sulla coronaria [(L.) B.H. Choi and H. Ohashi)] (syn. Hedysarum coronarium L.) is a semiperennial legume cultivated throughout the Mediterranean Basin as a biannual forage crop for grazing and/or hay or silage production [1]. It is well adapted to marginal and drought-prone environments, producing a good-quality, high-protein forage crop with moderate levels of condensed tannins, beneficial to ruminant production [1,2]. It also has potential as green manure [3]. Although sulla has been domesticated rather recently, it is grown all over the Mediterranean Basin, and has been introduced into other areas, notably New Zealand and Australia [4,5]. It was reported as the second largest forage after alfalfa by the 1990s in Southern Italy, grown over 300,000 ha [6]; however, little additional records on the actual cultivated area are available since this crop is mainly grazed. Wide morpho-physiological and adaptive diversity is available in the existing germplasm [1,7,8,9,10] which is being exploited in breeding programs.
Powdery mildew, often cited as “oidium”, is an important airborne fungal disease in many crops, including legumes [11,12,13], affecting all green parts of plants. The first symptoms are small, diffuse spots on leaflets and stipules that grow and become white to pale grey powdery areas that later coalesce and completely cover plant surfaces. Powdery mildew can be important also in sulla, but detailed information on its causal species is missing in most reports [9,14,15]. In fact, powdery mildews (Erysiphales) on legumes represent a taxonomically complicated group of diseases where species differentiation is mainly based on chasmothecia morphology, fruiting bodies that unfortunately are not always formed, hampering species determination. The only detailed taxonomic study reports Erysiphe hedysari (U. Braun) U. Braun and S. Takam. on Hedysarum spp. [16,17] infecting Anthyllis and Hedysarum. A number of powdery mildew species were previously cited on various Hedysarum sp. including Microsphaera trifolii (Grev.) U. Braun [18], Podosphaera leutricha [14], Microsphaera diffusa Cooke. et Peck [19] or Microsphaera hedysari Braun [20]. Former Erysiphe and Microsphaera are now treated as sections of Erysiphe emend. U. Braun and S. Takam. [16]. In addition, molecular characterization of the isolates infecting S. coronaria through examination of their ITS sequences is lacking.
A better understanding of the identity and host range of the possible powdery mildews infecting the cultivated S. coronaria is, therefore, needed to assist powdery mildew management. To this aim, we collected several powdery mildews infecting S. coronaria and characterized them at the cellular and molecular level through comparison of their ITS sequences. We also established their host range and compared them to other legume-infecting powdery mildews, showing that powdery mildew on S. coronaria is caused by distinct and highly specialized Erysiphe species.
2. Materials and Methods
2.1. Plant and Fungal Material
Nine populations of powdery mildew collected on Sulla coronaria in different years and regions were used in this study (Table 1). Three of these samples (Co08, Co09 and Co11) were collected from S. coronaria field nurseries in Córdoba over different years, in 2008, 2009 and 2011. Four isolates were collected from natural populations of S. coronaria in different locations of Cádiz province during 2008 to 2017, two of them (Ub10 and Ub17) in Ubrique, one (Je11) in Jerez de la Frontera and the last one (Ca09) in Tarifa counties. The last two populations were collected in 2017 in Oeiras, Portugal (Po17) and in 2011 in Hammemet, Tunisia (Tu11), respectively. All these samples were used for molecular studies. Four of them (Co11, Tu11, Je11 and Ub17) were also used for morphological studies together with powdery mildew populations collected on naturally infected pea and common bean growing near the collected S. coronaria plants (Table 1).
2.2. Morphology Assessments
For morphological measurement, well-formed powdery mildew colonies were scraped out of symptomatic leaves and transferred onto a microscope slide in a 10 µL drop of 0.1% trypan blue in lactoglycerol before visualization under bright-field microscopy (Leica DMLB, Leica Microsystems Wetzlar GmbH, Wetzlar, Germany) at ×40 and ×100 magnification [11,21]. Chasmothecia were never detected on the powdery mildew samples, not even on old material nor after maintaining infected leaves under a controlled environment at several temperatures between 20 and 25 °C until the end of the powdery mildew cycle. Morphological characters were, therefore, limited to the assessment of the vegetative growth stage and it included the length and width of conidia, and conidiophore foot cells. Measurements were based on the observation of 50 conidia and foot cells per sample. These measurements (Table 2) were compared with a morphological description of related legume-infecting powdery mildew species [22,23,24,25,26].
2.3. DNA Extraction and ITS Sequencing
The mycelium of each S. coronaria powdery mildew samples was obtained by scraping the corresponding symptomatic leaves. The obtained mycelium and conidia mixture were then collected in a 2 mL Eppendorf tube, flash frozen and maintained in −80 °C until DNA extraction. Genomic DNA was extracted following a previously reported protocol [27]. Molecular characterization of each S. coronaria powdery mildew sample was performed by analysis of the internal transcribed spacers (ITS). ITS sequences were obtained by nested PCR amplification with primers ITS-5/P3 and ITS1/P3. Each 50 μL reaction mixture contained 50 ng of template DNA, 2 units of BioTaq DNA polymerase (Bioline, London, UK), 1× PCR buffer, 2 mM MgCl2, 200 μM dNTPs, and 0.3 μM of each primer. The PCR amplifications were performed on a MyCycler (Biorad, Hercules, CA, USA) thermocycler as follows: 94 °C for 2 min, 40 cycles at 94 °C for 1 min, 55 °C for 2 min, and 72 °C for 2.5 min followed by a final step at 72 °C for 10 min. All amplifications were purified with the PCR cleanup kit of QIAgen and cloned into pGEMT vector (Promega, Madison, WI, USA). Two positive clones per amplicon were sequenced by STABVida (Setubal, Portugal) using the pGEMT vector-specific primers SP6 and T7. To characterize the ITS sequence, local pairwise comparison was performed to establish the level of similarity existing between them (Table 3). Pairwise analyses were performed with the Water pairwise algorithm implemented on the EMBOSS webserver [28]. Each sequence was also analyzed by BLAST onto the Genbank nr database to compare them with already-sequenced ITS with the online NCBI BLAST server (http://blast.ncbi.nlm.nih.gov/Blast.cgi; accessed and BLAST searches performed on the 10 November 2021). All sequences have been submitted to Genbank and assigned the reference numbers ON729985 to ON729992 (Table 1).
2.4. Phylogenetic Study
For the reconstruction of the phylogenetic relationship of the S. coronaria powdery mildew samples, the ITS sequences of these samples were aligned together with the ITS sequences of related Erysiphe species (Table 1). ITS sequences were aligned with the MAFFT algorithm [29] implemented in the NGphylogeny webserver (http://www.NGphylogeny.fr; accessed on 10 November 2021) [30] with default setting. The alignment was then manually corrected to resolve poorly aligned regions. To establish the phylogenetic relationship of S. coronaria powdery mildew samples, four different methods were used including neighbor joining (NJ), maximum parsimony (MP), maximum likelihood (ML) and Bayesian inference (BI). To remove gaps and trim alignment ends, alignment was further curated with the BMGE (for NJ and MP) [31] or Gblocks (ML and BI) [32] algorithms implemented in the NGPhylogeny webserver before phylogenetic analysis. The curated alignments were then analyzed with jModelTest 2.1.7 [33] to identify the optimum substitution model for NJ, ML and BI. NJ and MP trees were estimated with MEGA10 software [34] with 1000 bootstraps after partial gap deletions. Prior to NJ reconstruction, the distance matrix was obtained using the TN93 substitution model [35] with gamma distribution of 4 categories and α = 0.36. ML and BI trees were obtained based on 1000 bootstrap replicates following the GTR substitution model with gamma distribution of 4 categories and α = 0.39. The ML tree was obtained with PhyML 3.1 [36]. ML tree topology was optimized based on 5 random trees with the subtree pruning and regrafting (SPR) method. BI tree was obtained with MrBayes 3.2.7a [37]. Two chains were run simultaneously for 1 × 106 generations. Results from each independent run were compared to ensure convergence of the two runs onto a single stationary distribution of trees. The first 250,000 generations were discarded as burn-in, after which 2000 trees were sampled from each replicate run to determine the optimal consensus tree and calculate the posterior probabilities of the clades.
2.5. Host Range
Five accessions of Sulla coronaria, together with 5 to 16 accessions from species of the related genera Pisum, Lathyrus, Vicia, Cicer, Medicago, Trigonella and Phaseolus (Table 4 and Supplementary Table S1), were cross-inoculated with powdery mildew isolates collected on sulla (S. coronaria), pea (P. sativum; E. pisi), grasspea (L. sativus; E. trifoliorum), barrel medic (M. truncatula; E. trifoliorum) and one-flowered vetch (V. articulata; E. trifoliorum). Seeds from all accessions were planted in 0.5 L plastic pots containing a 1:1 sand–peat mixture, under controlled conditions (20 °C, 12 h light/12 h dark photoperiod). Each powdery mildew isolate was purified by single sporing and maintained in isolation at the Institute for Sustainable Agriculture, CSIC, Córdoba, on leaving plants of its respective crop on which it was collected, until use. Three independent inoculation experiments were performed with each isolate. Inoculations were performed on detached leaves from two-week-old seedlings (three to five seedlings per accession, per inoculation experiment) [38]. Inoculations with each isolate were performed on consecutive days to avoid isolate cross-contamination. Inoculations were performed in a settling tower [21,39]. Inoculum density was maintained at approximately 15 and 20 conidia/mm2. Petri dishes containing inoculated leaflets were placed in a growth chamber (20 °C, 12 h light/12 h dark photoperiod). Seven or eight days after inoculation, disease severity was assessed as the percentage of leaflet covered by the mycelium.
3. Results
3.1. Mycelial Morphology
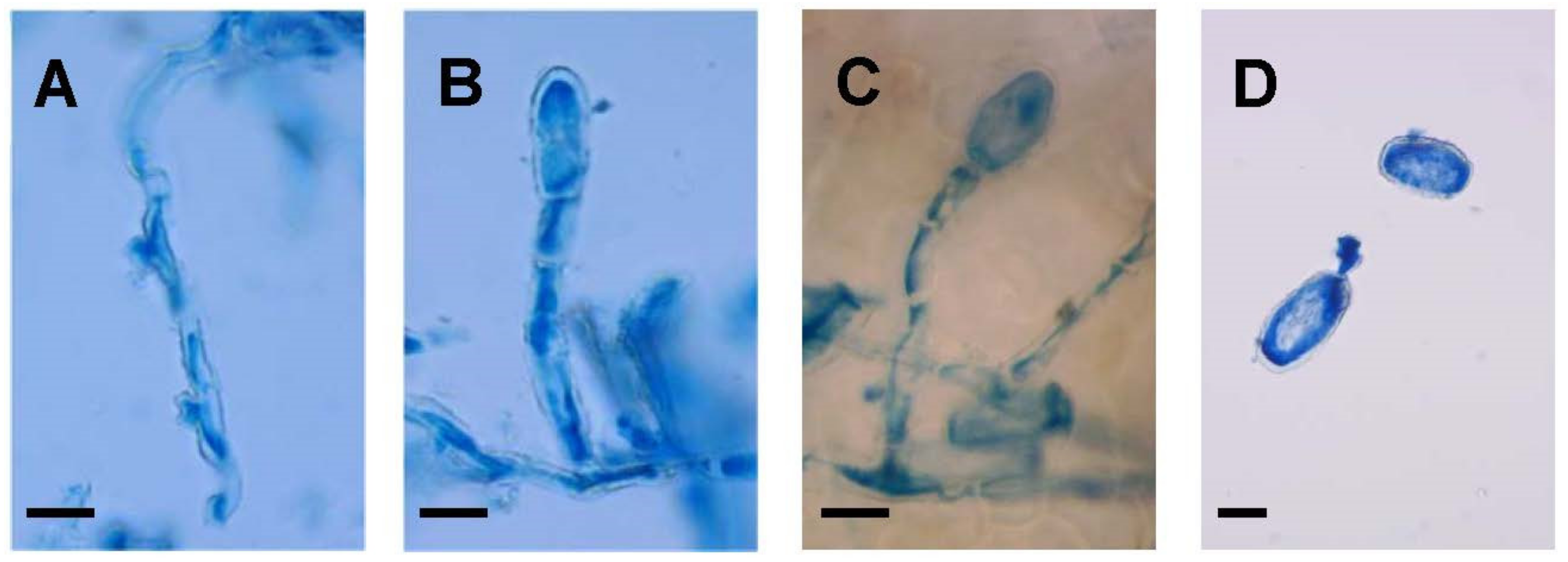
All samples collected on S. coronaria displayed typical powdery mildew symptoms. The mycelia of all samples were mainly epiphyllous in white effuse patches often covering the entire adaxial and abaxial leaf surfaces and stems. Hyphae were branched, septate, hyaline and thin-walled. In addition, infecting colonies developed lobed appressoria singly or in opposite pairs (Figure 1A). Conidiophores were composed of the conidial foot cell followed by 1 to 2 additional cell-forming terminal conidia singly (Figure 1B,C). Foot cells were erect and straight.
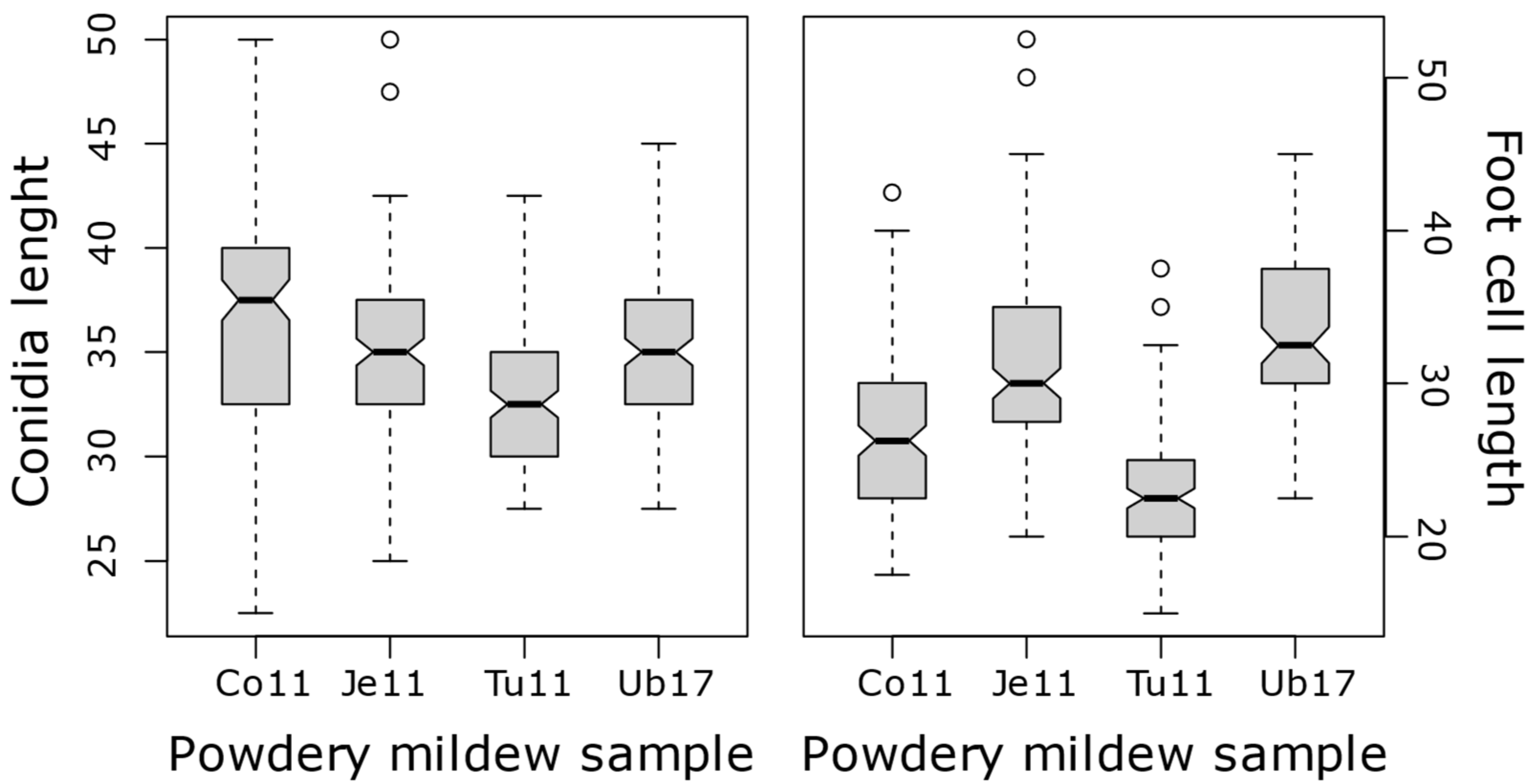
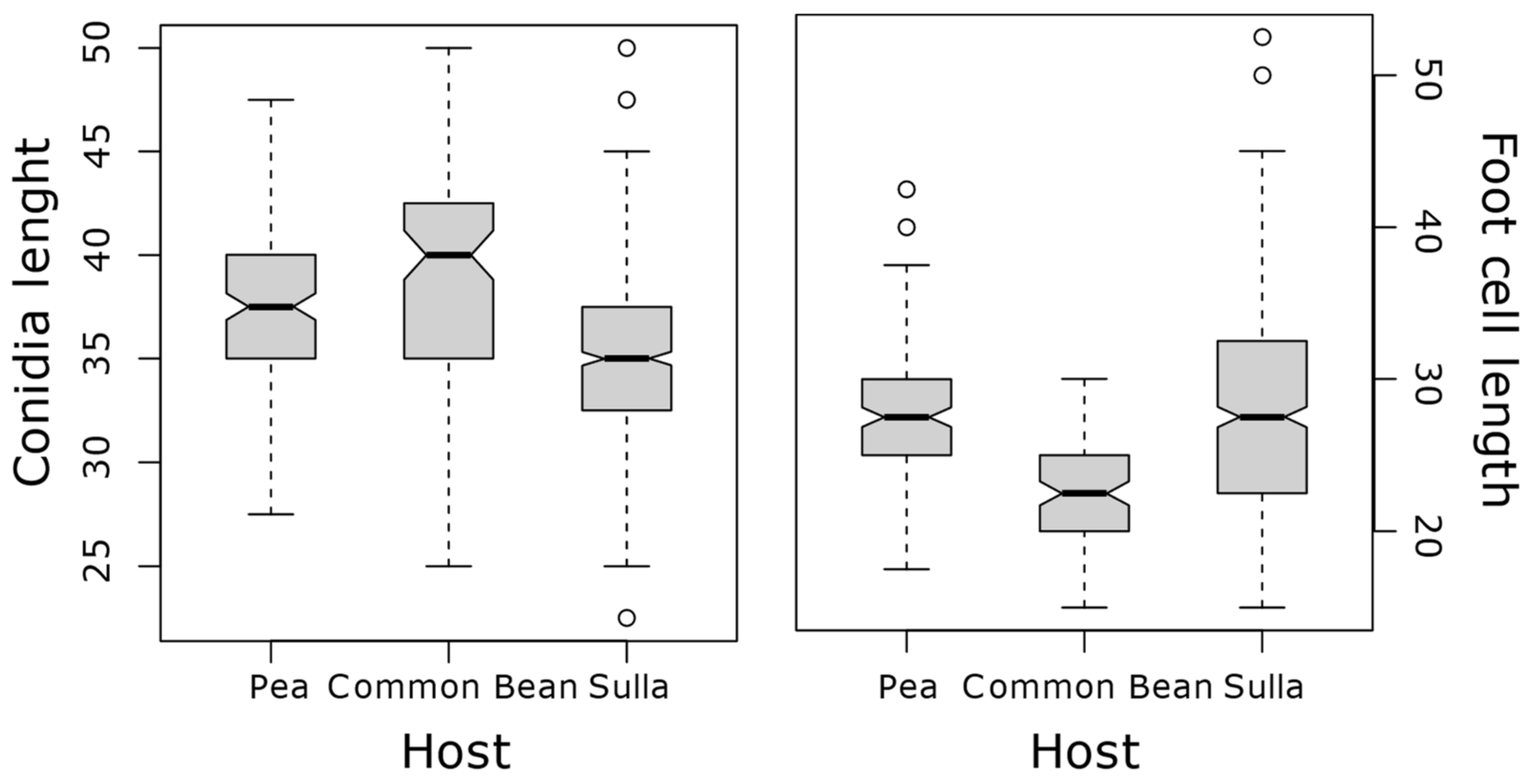
Small variations in conidia and foot cell measurement were detected between S. coronaria-infecting powdery mildew samples, although they were not statistically different (Figure 1D and Figure 2). Conidia and foot cell measurement was largely overlapping the variation detected for powdery mildew collected on nearby pea (E. pisi) and common bean (E. diffusa) plants, although they could be clearly differentiated from them (Figure 3). Altogether, conidia varied from 22.5 to 50 µm in length and from 10 to 22.5 µm in width while foot cell measurements varied from 15 to 52.5 µm in length and 5 to 12.5 µm in width (Table 2). All these features would support the assignation of these S. coronaria-infecting powdery mildew to a distinct Erysiphe species.
3.2. ITS Amplification
PCR amplification was successful for all samples except one (Je11) that turned out as being parasitized by some Ampelomyces sp. For all the other S. coronaria-infecting powdery mildew samples, a single ITS sequence was amplified and sequenced. The different S. coronaria-infecting powdery mildew sequences were almost identical, sharing between 98.8 and 99.9% of identity (Table 3). Blast analysis revealed strong homology with many Erysiphe species ITS sequences. The closest sequence shared 99.2% identity with the ITS from the S. coronaria-infecting samples. This sequence corresponded to a Pseudoidium sp. collected on Coronilla varia, another Fabaceae closely related to S. coronaria. The second and all subsequent matches corresponded to sequences of E. trifoliorum, E. palczewskii, E. longissima and E. guarinonii with homology ranging from 98 to 98.2% identity. Although the level of similarity between these sequences was very high, it was significantly lower than between the sequences of the different S. coronaria-infecting powdery mildews.
3.3. Phylogenetic Relationship
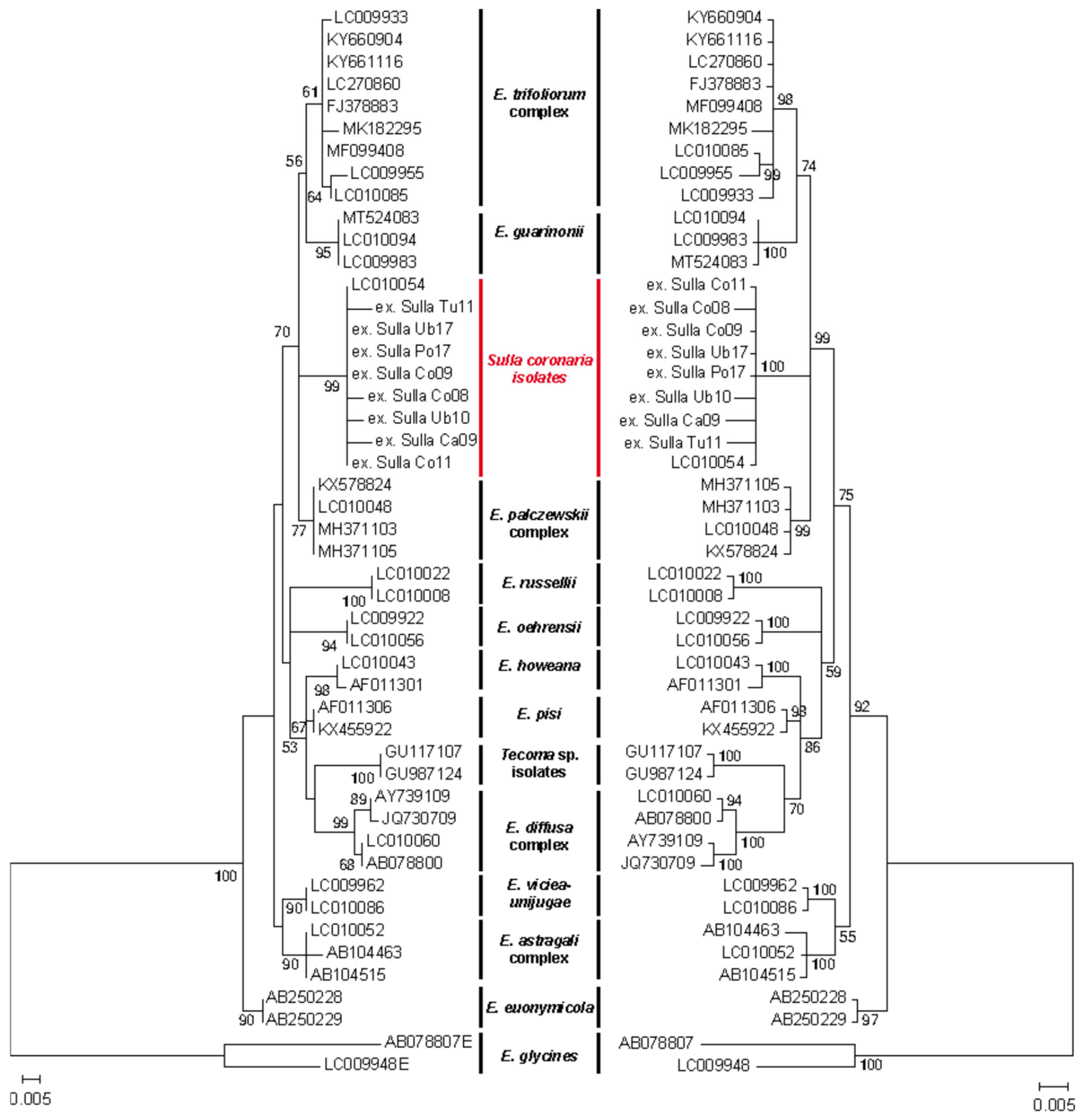
To decipher the identity of the powdery mildew infecting S. coronaria, we established the phylogenetic relationship existing between the ITS sequences of the different S. coronaria-infecting samples and with other Erysiphe species (Table 1). The four phylogenetic approaches performed (NJ, MP, ML and BI) led to very similar, if not identical, phylogenetic trees. To simplify, only the ML and BI trees are shown in Figure 4. The phylogenetic tree separates with strong support the different Erysiphe isolates according to their species or host plant with three exceptions. As expected, members of E. baeumleri, E. hyperici, E. intermedia and E. redens clustered together with the E. trifoliorum isolates forming the E. trifoliorum complex. Similarly, species of E. palczewskii infecting Caragana arborecens or Robinia sp. clustered together with the isolates from E. longissima that infect another related species, Caragana rosae, forming the E. palczewskii complex. Interestingly, all powdery mildew collected on S. coronaria clustered on a well-differentiated branch together with the isolate collected on Coronilla varia. In all phylogenetic trees, the cluster of the S. coronaria-infecting powdery mildew was located between the cluster of the E. trifoliorum complex and E. guarinonii and that of the E. palczewskii complex, but it was clearly differentiated from them which would support that they correspond to a distinct Erysiphe species.
3.4. Host Range
All five S. coronaria accessions were heavily infected by the sulla isolate (DS > 25%) (Table 4 and Supplementary Table S1) but not by any of the other isolates used. Conversely, the sulla isolate only infected the S. coronaria accessions, but not any of the other accessions from the other legume genera studied.
Such a level of specificity was not observed for the other powdery mildew isolates/legume genera studied. The pea (P. sativum) accessions were equally susceptible to pea (E. pisi) and Lathyrus (E. trifoliorum) isolates, some accessions being also slightly infected by E. trifoliorum isolates collected on M. truncatula and V. articulata. Wild pea (P. fulvum) accessions were not infected by the pea isolate, but were highly infected by the Lathyrus one. Lentil accessions were not infected by the pea isolate, but highly infected by Lathyrus and Vicia isolates, and also slightly by the Medicago one. All Lathyrus sp. accessions were heavily infected by the Lathyrus isolate but not by the Medicago one. The response to E. pisi varied, with high susceptibility for some L. cicera and L. sativus accessions, and rather low susceptibility for L. ochrus and L. clymenum accessions. L. ocrhus accessions could be severely infected by the Vicia isolate, by contrast to L. cicera and L. sativus accessions that were resistant.
Accessions of the various Vicia species genus were all resistant to Sulla and Medicago isolates, but could be heavily infected by the Vicia one and slightly infected by pea or Lathyrus isolates. The four alfalfa (M. sativa) accessions were resistant to all isolates whereas the two annual medics (M. truncatula and M. polymorpha) were heavily infected by the Medicago isolate and slightly by the Lathyrus one. The six fenugreek (T. foenum-graecum) accessions studied were highly resistant to Sulla, Pisum and Vicia isolates, whereas a few of them could be infected by Lathyrus or Medicago isolates. All accessions of Cicer spp. and Phaseolus spp. studied were immune to the different isolates tested.
4. Discussions
Powdery mildew is a widespread disease incited by numerous fungal species of the Erysiphaceae family. The taxonomy and host range of these fungal species remain, in many cases, uncertain. This is particularly true for the powdery mildews infecting legumes. In these host species, the disease is mainly caused by fungus belonging to the Erysiphe genera. The large morphological variability existing within and between Erysiphe species makes difficult taxonomic designation. The taxonomic characterization of Erysiphe species is mainly based on the examination of chasmothecia morphology, which is not always formed, or on the molecular analysis of ITS1 and ITS2 sequences. The combination of both approaches largely improves Erysiphe taxonomy, allowing clarification of the situation of many Erysiphe species [16,40]. However, description of chasmothecia morphology or ITS sequences is not always available to clearly assign a given isolate or population to an existing or novel Erysiphe species. The precise characterization of the causal agent of powdery mildew in a given crop is key for its control, particularly for resistance breeding. Therefore, efforts should be continued to expand the understanding of legume powdery mildews. To this aim, we targeted the powdery mildew infecting S. coronaria, a semiperennial legume with high potential as a protein source in pasture and forage which has been largely neglected so far.
Phylogenetic analysis of the powdery mildew samples collected on sulla clustered them together with the sequence of a powdery mildew collected on Coronilla varia on an independent clade between the E. guarinonii/E. trifoliorum and E. palczewskii clades. Interestingly, a detailed analysis of Erysiphe spp. phylogeny placed the C. varia sequence in a similar location [40] confirming that the sequences obtained on S. coronaria and C. varia correspond to a separate clade. The high similarity between the ITS sequences of the powdery mildew collected on C. varia and on S. coronaria and the phylogenetic analysis strongly suggest that all these powdery mildews correspond to the same Erysiphe species. Measurements of conidia and conidial foot cells of the sulla-infecting powdery mildew partially overlap those of other Erysiphe species (Figure 3 and Table 3) although they clearly differentiate from them, supporting the assumption that powdery mildew samples collected on S. coronaria belong to a distinct species as observed from the phylogenetic analysis. However, these data are not enough to ascribe these isolates to a specific Erysiphe species, confirming the limited taxonomic resolution of these morphological parameters [41]. The sole reported species on Hedysarum spp. have been described as E. hedysari based on their chasmothecia morphology [16,20]. As far as we know, our study is the first to report the molecular characterization of powdery mildew isolates on plants from the Hedysarum clade. However, the sexual stage of these isolates could not be detected on the samples examined despite examining older samples and after maintaining infected leaves at distinct temperatures. Therefore, it is impossible to ascertain whether the isolates collected on S. coronaria belong to E. hedysari as previously reported [16,20] or correspond to an as-yet undescribed Erysiphe species.
High specialization was observed for the powdery mildew of sulla. The sulla isolate only infected the S. coronaria accessions. None of the accessions from the other legume genera were infected by the isolate collected on sulla. In turn, all studied S. coronaria accessions were heavily infected by the sulla isolate but not by any other isolates. This level of specificity was not observed for the E. pisi and E. trifoliorum isolates collected on pea, grasspea, vetches or annual medic species, respectively, that could be infected by more than one isolate. None of the studied isolates infected faba bean (V. faba), alfalfa (M. sativa), or any of the Cicer or Phaseolus species studied, but we cannot comment on their eventual specificity against other powdery mildew species not included in this study. This is not surprising, as to the best of our knowledge, no significant powdery mildew damage occurs on alfalfa, faba bean or chickpea. Occasional powdery mildew infections on alfalfa have been reported in research nurseries and greenhouses but not in commercial fields [42]. Similarly, only sporadic infections were observed on faba bean. In both cases, the causal agent has been ascribed to E. pisi [42,43]. Only sporadic infections have also been observed on chickpea and ascribed to Leveillula taurica, E. pisi or other undetermined Erysiphe sp. [44]. By contrast, powdery mildew, caused by E. diffusa isolates, can be important on common bean under specific conditions [45]. Only the E trifoliorum isolates collected on Lathyrus and Medicago could infect some of the fenugreek accessions. In the literature, Leveillula taurica, Erysiphe polygoni and E. trifoliorum have been reported as the causal agents of powdery mildew on fenugreek [46,47]. All this confirms the little understanding on the causal agent and host range of the powdery mildew of many legume species, in spite of the implications in epidemiology and management. Knowledge of the host range is important to determine whether other crops could be affected by a given Erysiphe species. Furthermore, it is possible that alternative hosts could provide a means of overwintering of the pathogen, providing inocula to initiate epidemics in successive years.
A previous report on the host range of rust infecting Vicia species [48] agrees with the differential response to powdery mildew isolates seen here, where faba bean was not infected by any of the tested isolates, whereas several vetch species could be infected by several isolates. Uromyces viciae-fabae isolates from faba bean only infected faba bean, whereas isolates from vetch or lentil infected different Vicia spp. The host range expansion of powdery mildew fungi might be important to the understanding of their phylogenetic history and sometimes sheds light on the evolution of the hosts. The subgenus Vicia is smaller, less variable morphologically and comprises almost exclusively annuals including the most agriculturally important species V. faba and V. sativa. It is relevant that none of the powdery mildew isolates studied infected faba bean, and only a few slightly infected V. sativa, both of which belong to the most evolved subgenus Vicia. In contrast, species such as V. articulata, V. monanthos, V. villosa, and V. cracca of subgenus Vicilla that are considered more primitive and diverse [49] could be infected by several powdery mildew isolates, as found with the less specialized rusts [48]. This trend suggests a possible cospeciation of powdery mildew with host, as earlier suggested for rusts [50,51].
All sulla accessions studied were highly susceptible to the sulla powdery mildew. A small variation in levels of resistance to powdery mildew has been reported in field trials [9] but no details were provided on the actual levels of resistance, apart from stating that the cv. Grimaldi and the ecotype Tarifa were the most resistant among the studied accessions. However, Grimaldi was reported as susceptible under greenhouse conditions [14]. All this reinforces the need to first clarify the causal agent of sulla powdery mildew, and then to standardize resistance screenings to discern the actual levels of resistance available for breeding. Accordingly, we performed, for the first time, the molecular characterization of sulla-infecting powdery mildews and studied their host ranges. Altogether, we demonstrate that the sulla-infecting powdery mildew belongs to a highly specific species, presumably E. hedysari, which showed high specificity to its host. Beside clarifying the status of the sulla-infecting powdery mildew as an independent species, these results will be helpful for future breeding for powdery mildew resistance in sulla.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agronomy12081852/s1, Table S1: Host range: response of legume species to isolates of powdery mildew.
Author Contributions
D.R., A.M. and N.R. designed and performed the trials; D.R. and N.R. analyzed the data and wrote the manuscript. All authors have read and agreed to the published version of the manuscript, except late A.M. who participated in the analytical part the work but deceased before the manuscript was submitted.
Funding
This research was funded by the Junta de Andalucía Project LEGAND P20_00986 and Spanish AEI project PID2020-114668RB-100.
Acknowledgments
The authors are deeply indebted to M.C. Vaz Patto and M. Bani for their assistance in collecting isolates.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ruisi, P.; Siragusa, M.; Di Giorgio, G.; Graziano, D.; Amato, G.; Carimi, F.; Giambalvo, D. Pheno-morphological, agronomic and genetic diversity among natural populations of sulla (Hedysarum coronarium L.) collected in Sicily, Italy. Gen. Res. Crop Evol. 2011, 58, 245–257. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Mazza, F.; De Pasquale, C.; Giosuè, C.; Vitale, F.; Alabiso, M. Effects of ewes grazing sulla or ryegrass pasture for different daily durations on forage intake, milk production and fatty acid composition of cheese. Animal 2016, 10, 2074–2082. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, M.M.; Mojaradi, T.; Shirzadi, T. Effects of Hedysarum coronarium L. (sulla) as a green manure along with nitrogen fertilizer on maize Production. Agritech 2021, 41, 95–106. [Google Scholar] [CrossRef]
- Erwin, M.; Foster, K.; Skinner, P.; Nichols, P.; Nutt, B.; Snowball, R.; Beatty, R. Sulla and Purple Clover as Fodders. Limits on Seed Production. RIRDC Publication No 06/049. 2006. Available online: https://www.agrifutures.com.au/wp-content/uploads/publications/06-049.pdf (accessed on 18 April 2022).
- Crocker, G.; Hackney, B. Sulla. Primefact 745. State of New South Wales through NSW Department of Primary Industries, 2008. ISSN 1832-6668. Available online: www.dpi.nsw.gov.au/primefacts (accessed on 29 April 2022).
- Leto, G.; Todaro, M.; di Noto, A.M.; Alicata, M.L. Comparison of sulla-hay and sulla-silage in the lactating ewes and their effects on milk and cheese characteristics. Small Rum. Res. 2002, 45, 301–306. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Ruisi, P.; Di Miceli, G.; Pecetti, L. Morpho-physiological and adaptive variation of Italian germplasm of sulla (Hedysarum coronarium L.). Crop Pasture Sci. 2014, 65, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Córdoba, E.M.; Nadal, S.; Román, B.; Gónzalez-Verdejo, C.I. Collection, characterization and evaluation of wild (Hedysarum coronarium L.) populations from Andalusia (southern Spain). Aust. J. Crop Sci. 2013, 7, 165–172. [Google Scholar]
- Flores, F.; Gutiérrez, J.; López, J.; Moreno, M.T.; Cubero, M.T. Multivariate analysis approach to evaluate a germplasm collection of Hedysarum coronarium L. Gen. Resour. Crop Evol. 1997, 44, 545–555. [Google Scholar] [CrossRef]
- Marghali, S.; Panaud, O.; Lamy, F.; Ghariani, S.; Sarr, A.; Marrakchi, M.; Trifi-Farah, N. Exploration of intra- and inter-population genetic diversity in Hedysarum coronarium L. by AFLP markers. Gen. Resour. Crop Evol. 2005, 52, 277–284. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Prats, E.; Emeran, A.A.; Rubiales, D. Characterisation of resistance mechanisms to Erysiphe betae in Beta vulgaris. Phytopathology 2009, 99, 385–389. [Google Scholar] [CrossRef] [Green Version]
- Fondevilla, S.; Rubiales, D. Powdery mildew control in pea. A review. Agron. Sustain. Devel. 2012, 32, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Trabanco, N.; Pérez-Vega, E.; Campa, A.; Rubiales, D.; Ferreira, J.J. Genetic resistance to powdery mildew in common bean. Euphytica 2012, 186, 875–882. [Google Scholar] [CrossRef] [Green Version]
- Louati-Namouchi, I.; Louati, M.; Chriki, A. A quantitative study of some agronomic characters in Sulla (Hedysarum coronarium L.). Agronomie 2000, 20, 223–231. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and challenges in legume breeding for pest and disease resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef] [Green Version]
- Braun, U.; Takamatsu, S. Phylogeny of Erysiphe, Microsphaera, Uncinula (Erysipheae) and Cystotheca, Podosphaera, Sphaerotheca (Cystotheceae) inferred from rDNA ITS sequences—Some taxonomic consequences. Schlechtendalia 2000, 4, 1–33. [Google Scholar]
- Braun, U.; Kruse, J.; Wolcan, S.M.; Murace, M. Three new species of the genus Erysiphe (Ascomycota, Erysiphales) on legumes and some new combinations. Mycotaxon 2010, 112, 173–187. [Google Scholar] [CrossRef]
- Watson, M.J. Hedysarum coronarium—A legume with potential for soil conservation and forage. N. Z. J. Agric. Res. 1982, 16, 189–193. [Google Scholar]
- Chen, M.M. Forest Fungi Phytogeography: Forest Fungi Phytogeography of China, North America, and Siberia and International Quarantine of Tree Pathogens; Pacific Mushroom Research & Education Center: Sacramento, CA, USA, 2003; ISBN 0-9723916-6-5. [Google Scholar]
- Braun, U. Descriptions of new species and combinations in Microsphaera and Erysiphe (V). Mycotaxon 1984, 19, 374–383. [Google Scholar]
- Rubiales, D.; Carver, T.W.L. Defence reactions of Hordeum chilense accessions to three formae speciales of cereal powdery mildew fungi. Can. J. Bot. 2000, 78, 1561–1570. [Google Scholar] [CrossRef]
- Attanayake, R.N.; Glawe, D.A.; Dugan, F.M.; Chen, W. Erysiphe trifolii causing powdery mildew of lentil (Lens culinaris). Plant Dis. 2009, 93, 797–803. [Google Scholar] [CrossRef] [Green Version]
- Lebeda, A.; Mieslerova, B.; Sedlarova, M. First report of Erysiphe palczewskii on Caragana arborescens in the Czech Republic. Plant Pathol. 2008, 57, 779. [Google Scholar] [CrossRef]
- Meeboon, J.; Takamatsu, S. Notes on powdery mildews (Erysiphales) in Japan: I. Erysiphe Sect. Erysiphe. Mycoscience 2015, 56, 257–266. [Google Scholar] [CrossRef]
- Meeboon, J.; Takamatsu, S. Notes on powdery mildews (Erysiphales) in Thailand III. Erysiphe species on Fabaceae, Fagaceae, Hydrangeaceae and Lamiaceae. Tropical Plant Pathol. 2017, 42, 239–249. [Google Scholar] [CrossRef]
- Qiu, P.; Tang, S.; Guan, G.; Li, Y.; Takamatsu, S.; Liu, S. Revisiting a poorly known powdery mildew species: Morphology and molecular phylogeny of Erysiphe longissima. Mycoscience 2019, 60, 250–254. [Google Scholar] [CrossRef]
- Raeder, U.; Broda, P. Rapid preparation of DNA from filamentous fungi. Lett. Appl. Microbiol. 1985, 1, 17–20. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, W260–W265. [Google Scholar] [CrossRef] [Green Version]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Prats, E.; Llamas, M.J.; Rubiales, D. Characterisation of resistance mechanisms to Erysiphe pisi in Medicago truncatula. Phytopathology 2007, 97, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Martín, J.; Rubiales, D.; Prats, E. Resistance to powdery mildew (Blumeria graminis f.sp. avenae) in oat seedlings and adult plants. Plant Pathol. 2011, 60, 846–856. [Google Scholar]
- Takamatsu, S.; Ito, H.A.; Shiroya, Y.; Kiss, L.; Heluta, V. First comprehensive phylogenetic analysis of the genus Erysiphe (Erysiphales, Erysiphaceae) I. The Microsphaera lineage. Mycologia 2015, 107, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Takamatsu, S. Studies on the evolution and systematics of powdery mildew fungi. J. Gen. Plant Pathol. 2018, 84, 422–426. [Google Scholar] [CrossRef]
- Mohan, S.K.; Bijman, V.P.; Miller, D.R. Powdery Mildew Caused by Erysiphe pisi on Alfalfa in Idaho and Oregon. Plant Dis. 2001, 85, 678. [Google Scholar] [CrossRef]
- Cook, R.T.A.; Fox, R.T.V. Erysiphe pisi var. pisi on faba beans and other legumes in Britain. Plant Pathol. 1992, 41, 506–512. [Google Scholar] [CrossRef]
- Dugan, F.M.; Attanayake, K.; Glawe, D.; Chen, W. Powdery Mildew of Chickpea. Compendium of Chickpea and Lentil Diseases and Pests; American Phytopathological Society: Saint Paul, MN, USA, 2011; pp. 51–52. [Google Scholar]
- Leitão, S.T.; Almeida, N.F.; Moral, A.; Rubiales, D.; Vaz Patto, M.C. Identification of resistance to rust (Uromyces appendiculatus) and powdery mildew (Erysiphe difussa) in Portuguese common bean germplasm. Plant Breed. 2013, 132, 654–657. [Google Scholar] [CrossRef]
- Worku, J. Effects of Powdery Mildew (Leveillula Taurica and Erysiphe Polygoni) on yield and yield components of fenugreek in the mid-altitudes of Bale, South Eastern Ethiopia. J. Plant. Sci. 2017, 5, 65–67. [Google Scholar] [CrossRef]
- Bradshaw, M.; Braun, U.; Götz, M.; Jurick, W. Phylogeny and taxonomy of powdery mildew caused by Erysiphe species on Lupinus hosts. Mycologia 2022, 114, 76–88. [Google Scholar] [CrossRef]
- Rubiales, D.; Sillero, J.C.; Emeran, A.A. Response of vetches (Vicia spp.) to specialized forms of Uromyces vicia-fabae and to Uromyces pisi. Crop Prot. 2013, 46, 38–43. [Google Scholar]
- Maxted, N. An ecogeographical study of Vicia subgenus Vicia. In Systematic and Ecogeographic Studies on Crop Genepools; International Plant Genetic Resources Institute: Rome, Italy, 1995; Volume 8. [Google Scholar]
- Leppik, E.E. Evolutionary specialization of rust fungi (Uredinales) on the Leguminosae. Ann. Bot. Fenn. 1972, 9, 135–148. [Google Scholar]
- Van der Merwe, M.; Walker, J.; Ericson, L.; Burdon, J.J. Coevolution with higher taxonomic host groups within the Puccinia/Uromyces rust lineage obscured by host jumps. Mycol. Res. 2008, 112, 1387–1408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Micrographs of powdery mildew isolated on Sulla coronaria. Pictures show typical lobed appressoria (A) conidial foot cell composed of 1 or 2 intermediate cell(s) before the forming conidia (B,C) and mature and germinating conidia (D). All pictures were taken under bright-field microscope at ×40 magnification after staining with 0.1% trypan blue solution. Bar represents 10 µm.
Figure 1.
Micrographs of powdery mildew isolated on Sulla coronaria. Pictures show typical lobed appressoria (A) conidial foot cell composed of 1 or 2 intermediate cell(s) before the forming conidia (B,C) and mature and germinating conidia (D). All pictures were taken under bright-field microscope at ×40 magnification after staining with 0.1% trypan blue solution. Bar represents 10 µm.

Figure 2.
Box plots showing variations in conidial (left box) and foot cell (right box) lengths among 4 samples of powdery mildew collected on S. coronaria. The boxes and middle lines represent the middle 50 percentiles and medians, respectively. The whiskers represent upper and lower limits and circle represent outliers.
Figure 2.
Box plots showing variations in conidial (left box) and foot cell (right box) lengths among 4 samples of powdery mildew collected on S. coronaria. The boxes and middle lines represent the middle 50 percentiles and medians, respectively. The whiskers represent upper and lower limits and circle represent outliers.

Figure 3.
Box plots showing variations in conidial and foot cell lengths of powdery mildew samples collected on sulla (S. coronaria), pea and common bean. The boxes and middle lines represent the middle 50 percentiles and medians, respectively. The whiskers represent upper and lower limits and circle represents outliers.
Figure 3.
Box plots showing variations in conidial and foot cell lengths of powdery mildew samples collected on sulla (S. coronaria), pea and common bean. The boxes and middle lines represent the middle 50 percentiles and medians, respectively. The whiskers represent upper and lower limits and circle represents outliers.

Figure 4.
Phylogenetic relationship of legume-infecting Erysiphe spp. The figure represents the phylogenetic reconstruction of legume-infecting powdery mildews based on the analysis of ITS sequences with the maximum likelihood or Bayesian approaches. Numbers on node represent the bootstrap value or the Bayesian posterior probability value (BPP) expressed as percentages. Only bootstrap or BPP values >50% are shown. For both trees, scale bars below the trees represent numbers of substitution per sites. Sequences included in the trees are listed in Table 1.
Figure 4.
Phylogenetic relationship of legume-infecting Erysiphe spp. The figure represents the phylogenetic reconstruction of legume-infecting powdery mildews based on the analysis of ITS sequences with the maximum likelihood or Bayesian approaches. Numbers on node represent the bootstrap value or the Bayesian posterior probability value (BPP) expressed as percentages. Only bootstrap or BPP values >50% are shown. For both trees, scale bars below the trees represent numbers of substitution per sites. Sequences included in the trees are listed in Table 1.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of powdery mildew samples recollected from Sulla coronaria and other related Erysiphe species used for phylogenetic inferences, host range estimation and/or morphological studies.
Table 1.
List of powdery mildew samples recollected from Sulla coronaria and other related Erysiphe species used for phylogenetic inferences, host range estimation and/or morphological studies.
| Host | Powdery Mildew | Sample | Origin | Genbank |
|---|---|---|---|---|
| Sulla coronaria | Co08 | Córdoba, Spain | ON729986 | |
| S. coronaria | Co09 | Córdoba, Spain | ON729988 | |
| S. coronaria | Co11 | Córdoba, Spain | ON729985 | |
| S. coronaria | Ca09 | Tarifa, Spain | ON729987 | |
| S. coronaria | Ub10 | Ubrique, Spain | ON729991 | |
| S. coronaria | Je11 | Jerez de la Frontera, Spain | ||
| S. coronaria | Tu11 | Hammemet, Tunisia | ON729989 | |
| S. coronaria | Ub17 | Ubrique, Spain | ON729990 | |
| S. coronaria | Po17 | Oerias, Portugal | ON729992 | |
| Pisum sativum | E. pisi | Ps11 | Córdoba, Spain | |
| Phaseolus vulgaris | E. diffusa | Pv11 | Oerias, Portugal | |
| Lathyrus sativus | E. trifoliorum ex. L. sativus | Córdoba, Spain | ||
| Medicago truncatula | E. trifoliorum ex. M. truncatula | Córdoba, Spain | ||
| Vicia articulata | E. trifoliorum ex. V. articulata | Córdoba, Spain | ||
| Albizia julibrissin | Erysiphe cf. trifoliorum | MUMH0133 | Japan | LC010085 |
| Alhagi sp. | E. bremeri | AK113 | Guilan, Iran | AB104463 |
| Astragalus glycyphyllus | E. astragali | MUMH2585 | Ukraine | LC010052 |
| Astragalus sp. | E. astragali | Guilan, Iran | AB104515 | |
| Baptisia australis | E. guarinonii | HAL2337 | Germany | MT524083 |
| Caragana arborescens | E. palczewskii | MUMH2581 | Ukraine | LC010048 |
| C. rosea | E. longissima | HMJAU91780 | China | MH371103 |
| C. rosea | E. longissima | HMJAU91781 | China | MH371105 |
| Coronilla varia | Pseudoidium sp. | MUMH2587 | Ukraine | LC010054 |
| Desmodium incanum | E. diffusa | MUMH3121 | Corrientes, Argentina | LC010060 |
| D. laxum | E. glycines | MUMH396 | Shiga, Japan | LC009948 |
| Euonymus maackii | E. euonymi | HMJAU91794 | China | MK182295 |
| E. japonica | E. euonymicola | MUMH133 | Japan | AB250228 |
| E. japonica | E. euonymicola | MUMH2470 | Argentina | AB250229 |
| Glycine max | E. diffusa | MUMH791 | Oita, Japan | AB078800 |
| G. max | E. glycines | MUMH1462 | Mie, Japan | AB078807 |
| Hardenbergia sp. | P. hardenbergia | VPRI19879 | Australia | LC010094 |
| Hypericum ascyron | E. hypericin | F29454 | Jinan, Korea | MF099408 |
| Labrunum alpinum | E. guarinonii | MUMH1425 | Switzerland | LC009983 |
| Lathyrus latifolium | E. pisi | Belden, USA | AF011306 | |
| L. pratensis | E. ludens | OE2016PMCS45 | Balerno, UK | KY661116 |
| Lens culinaris | E. trifolii | LGH06 | Pullman, USA | FJ378883 |
| Lupinus sp. | E. intermedia | OE2015PMCS297 | Chelmsford, UK | KY660904 |
| Maytenus boaria | E. oehrensii | MUMH2492 | Bariloche, Argentina | LC010022 |
| M. boaria | E. oehrensii | MUMH1936 | Bariloche, Argentina | LC010008 |
| Medicago littoralis | E. trifoliorum | MUMH7038 | Baku, Azerbaijan | LC270860 |
| Oenothera amoena | E. howeana | MUMH2572 | The Netherlands | LC010043 |
| O. biennis | E. howeana | UC1512301 | Redlands, USA | AF011301 |
| Oxalis corniculatus | E. russellii | MUMH0105 | Mie, Japan | LC009922 |
| Phaseolus vulgaris | Erysiphe sp. | EB2004 | Londrina, Brazil | AY739109 |
| Pisum sativum | E. pisi | P1 | India | KX455922 |
| Robinia pseudoacacia | E. palczewskii | ZKEP001 | China | KX578824 |
| Senna septemtrionalis | Erysiphe sp. | 3D | Buenavista, Mexico | JQ730709 |
| Tecoma capensis | Erysiphe sp. | DAG08-36 | Yuma, USA | GU117107 |
| T. capensis | Erysiphe sp. | 08-3618S | Yuma, USA | GU987124 |
| Trifolium arvense | E. trifoliorum | MUMH0701 | Budapest, Hungria | LC009955 |
| Vicia hirsute | E. baeumleri | MUMH0240 | Shiga, Japan | LC009933 |
| V. angustifolius | E. viciae-unijugae | MUMH0817 | Yamanashi, Japan | LC009962 |
| V. unijuga | E. viciae-unijugae | TPU-153 | Japan | LC010086 |
| Xanthoxalis sp. | E. russellii | MUMH2593 | Ukraine | LC010056 |
Table 2.
Morphological characterization of powdery mildew samples from S. coronaria and comparison with samples collected on nearby Pisum sativum and Phaseolus vulgaris plants and with related Erysiphe species.
Table 2.
Morphological characterization of powdery mildew samples from S. coronaria and comparison with samples collected on nearby Pisum sativum and Phaseolus vulgaris plants and with related Erysiphe species.
| Erysiphe sp. ex. Sulla coronaria | E. pisi Ps11 | E. diffusa Pv11 | E. palcewskii [23] | E. longissimi [26] | E. trifoliorum [24] | E. pisi [25] | E. diffusa [22] | |
|---|---|---|---|---|---|---|---|---|
| conidia | ||||||||
| Length (µm) | 22.5–50 | 27.5–47.5 | 25–50 | 20–37.5 | 19.2–48.2 | 25.5–39.5 | 30–44 | 28–36 |
| Width (µm) | 10–22.5 | 12.5–20 | 15–22.5 | 8.75–17.5 | 12.3–17.2 | 12–18 | 18–20 | 12.5–16 |
| Ratio L/W | 1–5 | 1.4–3.8 | 1.1–3.3 | 1.54–3.14 | 1.4–2.9 | 1.4–3.3 | 1.5–2.4 | 1.8–2.9 |
| Foot cell | ||||||||
| Length (µm) | 15–52.5 | 17.5–42.5 | 15–30 | 15–40 | 24.5–54.2 | 13–45 | 35–70 | 25–33 |
| Width (µm) | 5–12.5 | 5–12.5 | 7.5–10 | nd | 6–9.1 | 7–9.5 | 8–12.5 | 7.7–9 |
| Ratio L/W | 1.2–10.5 | 1.4–8.5 | 1.5–4 | nd | 2.69–9.03 | 1.4–6.4 | 2.8–8.75 | 2.8–4.3 |
Table 3.
Comparison of ITS sequence from S. coronaria-infecting powdery mildew samples. Values in the lower diagonals are % similarity as estimated by pairwise local comparison.
Table 3.
Comparison of ITS sequence from S. coronaria-infecting powdery mildew samples. Values in the lower diagonals are % similarity as estimated by pairwise local comparison.
| Co08 | Co09 | Co11 | Ca08 | Tu11 | Ub17 | Ub10 | Po17 | |
|---|---|---|---|---|---|---|---|---|
| Co08 | 100 | |||||||
| Co09 | 99.4 | 100 | ||||||
| Co11 | 99.2 | 99.7 | 100 | |||||
| Ca09 | 98.9 | 99.1 | 99.1 | 100 | ||||
| Tu11 | 98.9 | 99.2 | 99.2 | 98.8 | 100 | |||
| Ub17 | 99.2 | 99.7 | 99.7 | 99.2 | 99.2 | 100 | ||
| Ub10 | 99 | 99.7 | 99.6 | 99 | 99.1 | 99.7 | 100 | |
| Po17 | 99.4 | 99.9 | 99.9 | 99.3 | 99.4 | 99.9 | 99.7 | 100 |
Table 4.
Comparison of host range of the S. coronaria isolate with other isolates infecting closely related legume genera including E. pisi and E. trifoliorum isolates. *(- = DS < 1%; x = 1% < DS < 25%; xx = 26% < DS < 50%; xxx = DS > 51%; ns = not studied).
Table 4.
Comparison of host range of the S. coronaria isolate with other isolates infecting closely related legume genera including E. pisi and E. trifoliorum isolates. *(- = DS < 1%; x = 1% < DS < 25%; xx = 26% < DS < 50%; xxx = DS > 51%; ns = not studied).
| Response* to Powdery Mildew Isolate | |||||||
|---|---|---|---|---|---|---|---|
| Crop | Host specie | Number of accessions studied | Erysiphe sp. ex S. coronaria | E. pisi ex P. sativum | E. trifoliorum ex L. sativus | E. trifoliorum ex M. truncatula | E. trifoliorum ex V. articulata |
| Sulla | Sulla coranaria | 5 | xx/xxx | - | - | - | - |
| Pea and related sp. | Pisum sativum | 6 | - | xx/xxx | xx/xxx | -/x | x |
| Pisum fulvum | 2 | - | - | xxx | -/x | - | |
| Lentil | Lens culinaris | 5 | - | - | xx/xxx | x | xxx |
| Grasspea and related sp. | Lathyrus sativus | 5 | - | x/xx | xxx | - | - |
| Lathyrus cicera | 6 | - | x/xxx | x/xxx | - | - | |
| Lathyrus ochrus | 3 | - | -/x | xxx | - | xx | |
| Lathyrus clymenum | 2 | - | -/x | xxx | - | ns | |
| Vetches | Vicia articulata | 1 | - | - | xx | - | xxx |
| Vicia monantha | 2 | - | x | ns | - | xxx | |
| Vicia villosa | 1 | - | ns | - | - | xx | |
| Vicia sativa | 5 | - | - | -/x | - | -/x | |
| Vicia pannonica | 1 | - | - | x | - | - | |
| Vicia tetrasperma | 1 | - | - | - | - | - | |
| Faba bean | Vicia faba | 5 | - | - | - | - | - |
| Fenugreek | Trigonella foenun-graecum | 6 | - | - | -/xx | -/xx | - |
| Annual medics | Medicago truncatula | 1 | - | ns | x | xxx | ns |
| Medicago polymorpha | 1 | - | - | x | xxx | ns | |
| Alfalfa | Medicago sativa | 4 | - | - | - | - | ns |
| Chickpea and related sp. | Cicer arietinum | 5 | - | - | - | - | - |
| Cicer reticulatum | 2 | - | - | - | - | ns | |
| Common bean and related sp. | Phaseolus vulgaris | 9 | - | - | - | - | - |
| Phaselus filiformis | 2 | - | - | - | - | ns | |
| Phaseolus leptostachyus | 2 | - | - | - | - | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rubiales, D.; Moral, A.; Rispail, N. Sulla Powdery Mildew: Phylogeny and Host Range. Agronomy 2022, 12, 1852. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081852
AMA Style
Rubiales D, Moral A, Rispail N. Sulla Powdery Mildew: Phylogeny and Host Range. Agronomy. 2022; 12(8):1852. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081852
Chicago/Turabian StyleRubiales, Diego, Ana Moral, and Nicolás Rispail. 2022. "Sulla Powdery Mildew: Phylogeny and Host Range" Agronomy 12, no. 8: 1852. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081852
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.