
Effect of Brown Algae (Fucus vesiculosus L.) on Humus and Chemical Properties of Soils of Different Type and Postgermination Growth of Cucumber Seedlings

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Soils Description
2.2. Physicochemical Analysis of Soil
2.3. Germination of Cucumber Seeds in Various Research Media
2.4. Statistical Analysis
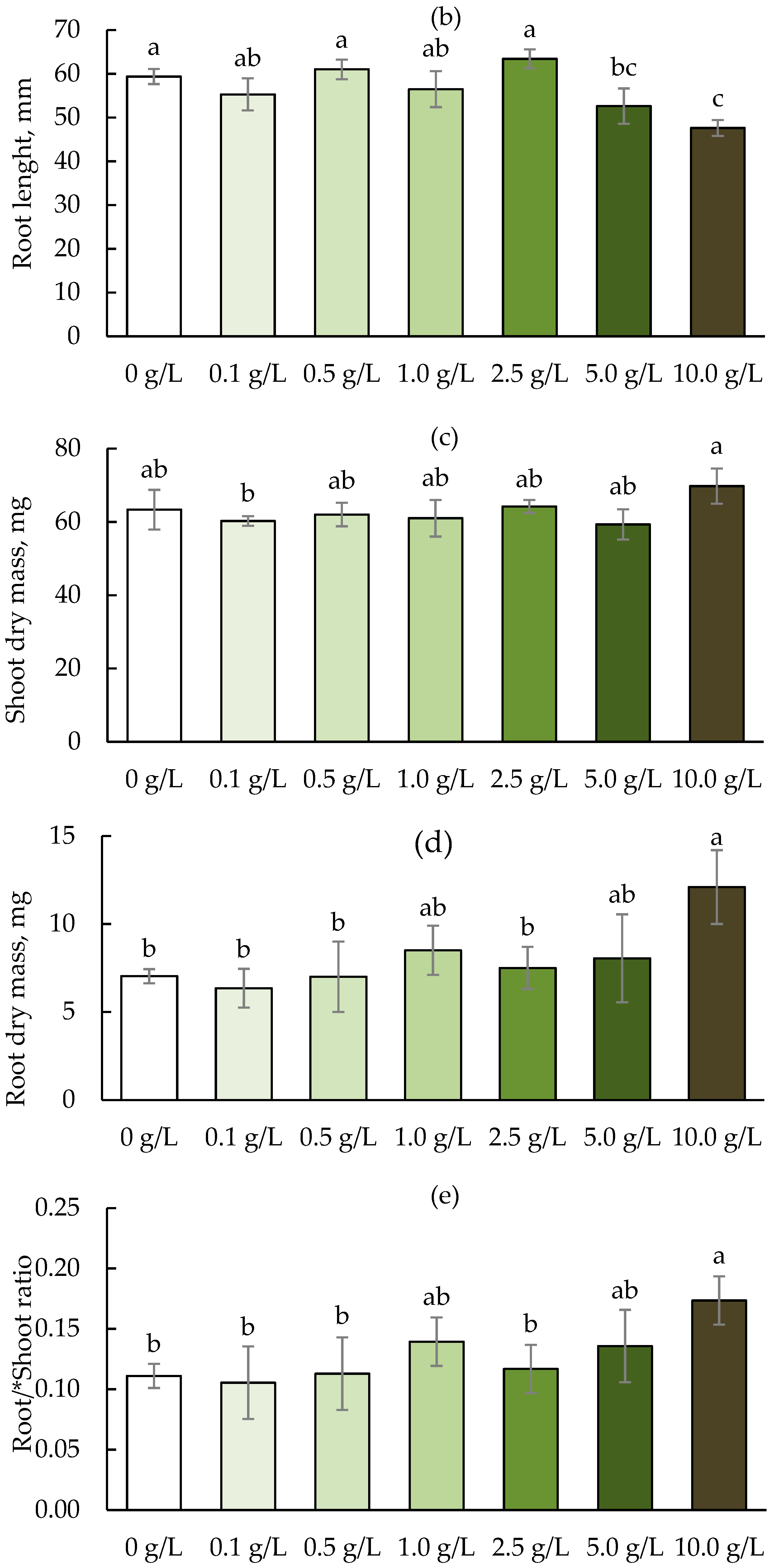
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krasilnikov, P.V. Stable carbon compounds in soils: Their origin and functions. Eurasian Soil Sci. 2015, 48, 997–1008. [Google Scholar] [CrossRef]
- Butphu, S.; Rasche, F.; Cadisch, G.; Kaewpradit, W. Eucalyptus biochar application enhances Ca uptake of upland rice, soil available P, exchangeable K, yield, and N use efficiency of sugarcane in a crop rotation system. J. Plant Nutr. Soil Sci. 2020, 183, 58–68. [Google Scholar] [CrossRef]
- Ikkonen, E.; Chazhengina, S.; Butilkina, M.; Sidorova, V. Physiological response of onion (Allium cepa L.) seedlings to shungite application under two soil water regimes. Acta Physiol. Plant. 2021, 43, 75–88. [Google Scholar] [CrossRef]
- Ikkonen, E.; Chazhengina, S.; Jurkevich, M. Photosynthetic Nutrient and Water Use Efficiency of Cucumissativus under Contrasting Soil Nutrient and Lignosulfonate Levels. Plants 2021, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, I.V.; Utkaeva, V.F.; Bondarev, A.G. Assessment of changes in the physical properties of plowed loamy soddypodzolic soils in the nonchernozemic zone of European Russia under the impact of anthropogenic loads. Eurasian Soil Sci. 2009, 42, 137–146. [Google Scholar] [CrossRef]
- Craigie, J.S. Cell walls. In Biology of the Red Algae; Cole, K.M., Sheath, R.G., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 221–257. [Google Scholar] [CrossRef]
- Janceva, S.; Lauberte, L.; Vevere, L.; Bikovens, O.; Andersone, A.; Telysheva, G. Brown Algae as a Potential Source for Obtaining of High Valuable Products. Key Eng. Mater. 2019, 800, 98–102. [Google Scholar] [CrossRef]
- Bikovens, O.; Ponomarenko, J.; Janceva, S.; Lauberts, M. Development of the approaches for complex utilization of brown algae (Fucusvesiculosus) biomass for the obtaining of value-added products. In Proceedings of the Rural Development Conference, Kaunas, Lithuania, 23–24 November 2017. [Google Scholar] [CrossRef]
- Norrie, J.; Keathley, J.P. Benefits of Ascophyllumnodosum marine-plant extract applications to ‘Thompson seedless’ grape production. ActaHortic. 2006, 727, 243–247. [Google Scholar] [CrossRef]
- Dmytryk, A.; Chojnacka, K. Algae as fertilizers, biostimulants, and regulators of plant growth. In Algae Biomass: Characteristics and Applications; Chojnacka, K., Schroeder, P.P.W.G., Michalak, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 115–122. [Google Scholar] [CrossRef]
- Sunarpi, H.; Nikmatullah, A.; Sunarwidhi, A.L.; Sapitri, I.; Ilhami, B.T.K.; Widyastuti, S.; Prasedya, E.S. Growth and yield of rice plants (Oryza sativa) grown in soil media containing several doses of inorganic fertilizers and sprayed with lombok brown algae extracts. IOP Conf. Ser. Earth Environ. Sci. 2020, 594, 012032. [Google Scholar] [CrossRef]
- Chatterjee, A.; Singh, S.; Agrawal, C.; Yadav, S.; Rail, R.; Rai, C. Role of algae as a biofertilizer. In Algal Green Chemistry; Rastogy, R.P., Madamwar, D., Panday, A., Eds.; Elsevier: Amsterdam, The Netherland, 2017; pp. 189–200. [Google Scholar] [CrossRef]
- Brahmbhatt, N.H.; Kalasariya, H.S. Effect of algae on seedling growth of “Queen of Forages”. Int. J. Eng. Res. Generic Sci. 2015, 3, 827–833. [Google Scholar]
- Saadaoui, I.; Sedky, R.; Rasheed, R.; Bounnit, T.; Almahmoud, A.; Elshekh, A.; Dalgamouni, T.; Al Jmal, K.; Das, P.; Al Jabr, H. Assessment of the algae-based biofertilizer influence on date palm (Phoenix dactylifera L.) cultivation. J. Appl. Phycol. 2019, 31, 457–463. [Google Scholar] [CrossRef]
- Eyras, M.C.; Defosse, G.E.; Dellatorre, F. Seaweed compost as an amendment for horticultural soils in Patagonia, Argentina. Compos. Sci. Util. 2008, 16, 119–124. [Google Scholar] [CrossRef]
- Moore, K.K. Using seaweed compost to grow bedding plants. BioCycle 2004, 45, 43–44. [Google Scholar]
- Chen, S.K.; Edwards, C.A.; Subler, S. The influence of two agricultural biostimulants on nitrogen transformations. microbial activity. and plant growth in soil microcosms. Soil Biol. Biochem. 2003, 35, 9–19. [Google Scholar] [CrossRef]
- Zaborowska, M.; Kucharski, J.; Wyszkowska, J. Brown algae and basalt meal in maintaining the activity of arylsulfatase of soil polluted with cadmium. Water Air Soil Pollut. 2017, 228, 267. [Google Scholar] [CrossRef]
- Berthon, J.-Y.; Michel, T.; Wauquier, A.; Joly, P.; Gerbore, J.; Filaire, E. Seaweed and microalgae as major actors of blue biotechnology to achieve plant stimulation and pest and pathogen biocontrol—A review of the latest advances and future prospects. J. Agric. Sci. 2021, 159, 523–534. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; El-Shershaby, N.A. Algae as Bio-fertilizers: Between current situation and future prospective. Saudi J. Biol. Sci. 2022, 29, 3083–3096. [Google Scholar] [CrossRef]
- Agroclimatic Resources of the Karelian ASSR; Gidrometeoizdat: Leningrad, Russia, 1974; p. 115. (In Russian)
- GOST 54650-2011 Soils; Determination of Mobile Phosphorus and Potassium Compounds by Kirsanov Method Modified by CINAO. Standartinform: Moscow, Russian, 2020. Available online: http://docs.cntd.ru/document/1200094361 (accessed on 20 December 2020). (In Russian)
- Van Reeuwijk, L.P. Procedures for Soil Analysis; Technical Paper 9; ISRIC: Wageningen, The Netherlands, 2002; p. 95. [Google Scholar]
- Zhang, J.; Zhang, Q.; Wang, J.; Shi, X.; Zhang, Z. Analysis of The Monosaccharide Composition of Fucoidan by Precolumn Derivation Hplc. Chin. J. Oceanol. Limnol. 2009, 27, 578–582. [Google Scholar] [CrossRef]
- Semenova, E.V.; Chebotok, V.V.; Borisovskaja, I.V. To the question of the use of alginates from brown algae in medicine and pharmacy. Mod. Probl. Sci. Educ. 2019, 6, 1. (In Russian) [Google Scholar]
- Podkoritova, A.V.; Roschina, A.N. Marine Brown Algae—A Promising Source of BAS for Medical, Pharmaceutical and Food Applications. Tr. Vniro 2021, 186, 156–172. (In Russian) [Google Scholar] [CrossRef]
- Fedorets, N.G.; Bakhmet, O.N. Features of the formation of soils and soil cover of the Karelian-Kola region. Proc. Karelian Sci. Cent. Russ. Acad. Sci. 2016, 12, 39–51. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Morozova, P.M.; Fedorets, N.G. Land Resources of Karelia and Their Protection; KarRC RAN: Petrozavodsk, Russia, 2004; p. 152. (In Russian) [Google Scholar]
- Sáez-Plaza, P.; Michałowski, T.; Navas, M.J.; Asuero, A.G.; Wybraniec, S. An overview of the Kjeldahl method of nitrogen determination. Part I. Early history, chemistry of the procedure, and titrimetric finish. Crit. Rev. Anal. Chem. 2013, 43, 178–223. [Google Scholar] [CrossRef]
- Usov, A.I.; Bilan, M.I. Fucoidans-Sulfated Polysaccharides of Brown Algae. Adv. Chem. 2009, 78, 846–862. (In Russian) [Google Scholar] [CrossRef]
- Mendoza-Aguirre, O.M.; De Neve, S.; Deroo, H.; Li, H.; Sleutel, S. Do interactions between application rate and native soil organic matter content determine the degradation of exogenous organic carbon? Soil Biol. Biochem. 2022, 164, 108473. [Google Scholar] [CrossRef]
- Hong, H.; Chen, S.; Fang, Q.; Algeo, T.J.; Zhao, L. Adsorption of organic matter on clay minerals in the Dajiuhu peat soil chronoseuence, South China. Appl. Clay Sci. 2019, 178, 105125. [Google Scholar] [CrossRef]
- Fu, W.; Fan, J.; Wang, S.; Wang, H.; Dai, Z.; Zhao, X.; Hao, M. Woody peat addition increases soil organic matter but its mineralization is affected by soil clay in the four degenerated erodible soils. Agric. Ecosyst. Environ. 2021, 318, 107495. [Google Scholar] [CrossRef]
- Vidal, A.; Lenhart, T.; Dignac, M.F.; Biron, P.; Höschen, C.; Barthod, J.; Vedere, C.; Vaury, V.; Bariac, T.; Rumpel, C. Promoting plant growth and carbon transfer to soil with organic amendments produced with mineral additives. Geoderma 2020, 374, 114454. [Google Scholar] [CrossRef]
- Wen, Y.; Zang, H.; Freeman, B.; Musarika, S.; Evans, S.D.; Chadwick, D.R.; Jones, D.L. Microbial utilization of low molecular weight organic carbon substrates in cultivated peats in response to warming and soil degradation. Soil Biol. Biochem. 2019, 139, 107629. [Google Scholar] [CrossRef]
- AminiTabrizi, R.; Dontsova, K.; Grachet, N.G.; Tfaily, M.M. Elevated temperatures drive abiotic and biotic degradation of organic matter in a peat bog under oxic conditions. Sci. Total Environ. 2022, 804, 150045. [Google Scholar] [CrossRef]
- Bhattacharyya, S.S.; Ros, G.Р.; Furtak, K.; Iqbal, H.M.N.; Parra-Saldívar, R. Soil carbon sequestration—An interplay between soil microbial community and soil organic matter dynamics. Sci. Total Environ. 2022, 815, 152928. [Google Scholar] [CrossRef]
- Gryta, A.; Frąc, M.; Oszust, K. Community shift in structure and functions across soil profile in response to organic waste and mineral fertilization strategies. Appl. Soil Ecol. 2019, 143, 55–60. [Google Scholar] [CrossRef]
- Zhuang, Y.; Zhu, J.; Shi, L.; Fu, Q.; Hu, H.; Huang, Q. Influence mechanisms of iron, aluminum and manganese oxides on the mineralization of organic matter in paddy soil. J. Environ. Manag. 2022, 301, 113916. [Google Scholar] [CrossRef] [PubMed]
- Saleem, H.; Ahmad, M.; Rashid, J.; Ahmad, M.; Mohammad, I.; Alwabel, M.I.; Amin, M. Carbon potentials of different biochars derived from municipal solid waste in a saline soil. Pedosphere 2022, 32, 283–293. [Google Scholar] [CrossRef]
- Orlov, D.S. Soil Chemistry, 1st ed.; CRC Press: Rotterdam, The Netherlands, 1992; p. 402. [Google Scholar]
- Zhao, X.; Zhao, C.; Stahr, K.; Kuzyakov, Y.; Wei, X. The effect of microorganisms on soil carbonate recrystallization and abiotic CO2 uptake of soil. Catena 2020, 192, 104592. [Google Scholar] [CrossRef]
- Cui, L.; Liu, Y.; Yan, J.; Hina, K.; Hussain, Q.; Qiu, T.; Zhu, J. Revitalizing coastal saline-alkali soil with biochar application for improved crop growth. Ecol. Eng. 2022, 179, 106594. [Google Scholar] [CrossRef]
- Richardson, J.B.; Johnston, M.R.; Herrick, B.M. Invasive earthworms Amynthastokioensis and Amynthasagrestis alter macronutrients (Ca, Mg, K, P) in field and laboratory forest soils. Pedobiologia 2022, 91–92, 150804. [Google Scholar] [CrossRef]




{kind=link}
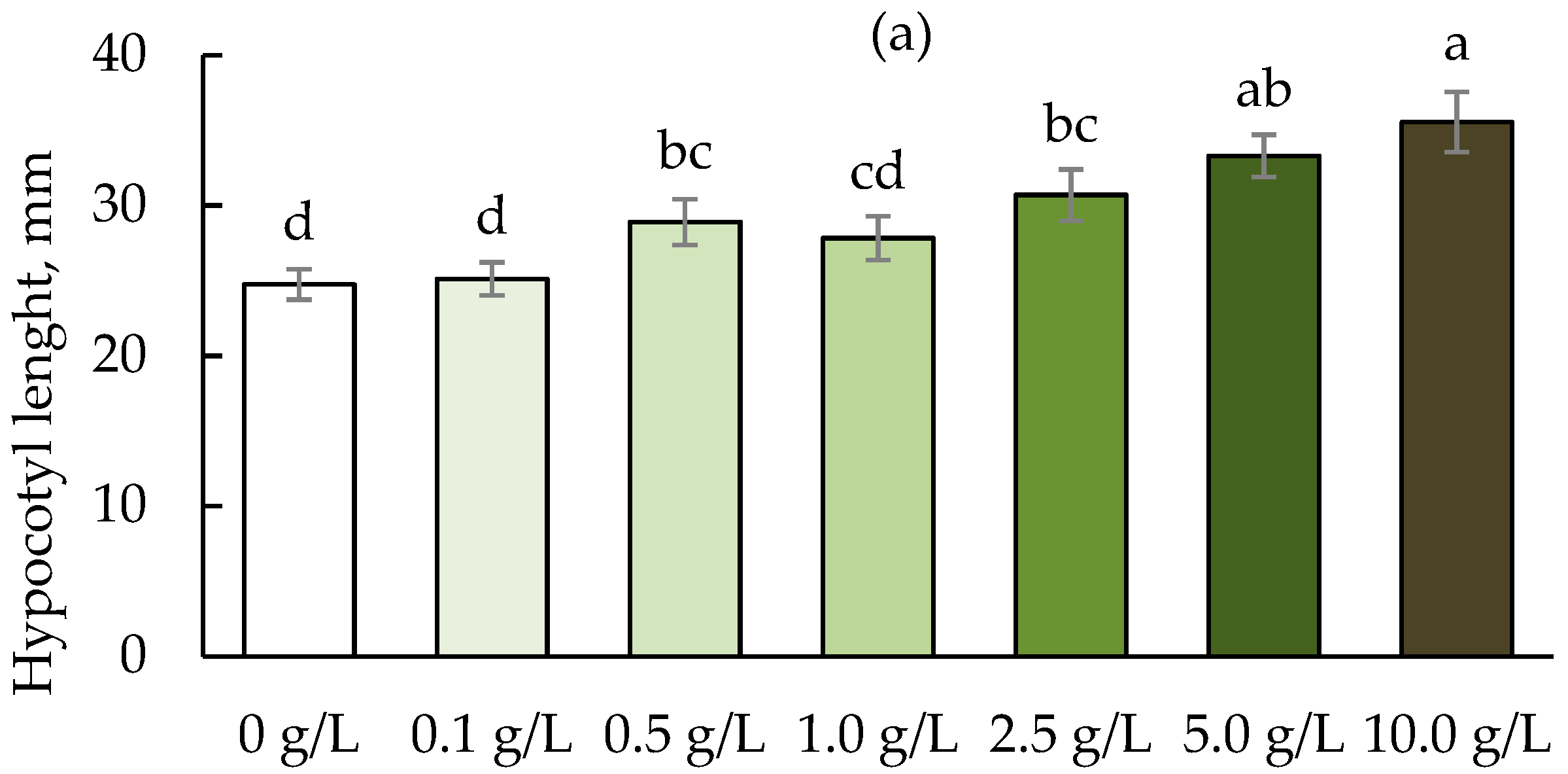{kind=link}
{kind=link}
| F. vesiculosus Material | Phenolic Compounds | Proline | Alginate | Mannitol | Total Sugar Content |
|---|---|---|---|---|---|
| n = 5 | |||||
| Waste | 2.10 ± 0.03 | 0.048 ± 0.004 | 7.4 ± 0.03 | 1.1 ± 0.1 | 104.1 ± 5.2 |
| F. vesiculosus Material | Type of Sugar | Sugar Content, % |
|---|---|---|
| n = 5 | ||
| Waste | Ribose | 4.0 ± 0.4 |
| Xylose | 12.9 ± 1.6 | |
| Hexose | 10.6 ± 0.8 | |
| Pentose | 26.4 ± 2.7 | |
| Glucose | 5.1 ± 0.5 | |
| Galactose | 23.4 ± 2.1 |
| Treatments | pHH2O | C Total | N Total | P2O5 | Ca2+ | Mg2+ | Na+ | К+ | Dry Residue |
|---|---|---|---|---|---|---|---|---|---|
| % | mg 100g−1 soil | mg kg−1 soil | % | ||||||
| n = 5 | |||||||||
| Retisol loamy sand | |||||||||
| 0 | 5.41 | 1.28 | 0.12 | 10.2 | 394.0 | 18.7 | 40.6 | 34.4 | 0.10 |
| 0.5 | 5.38 | 1.62 * | 0.13 | 9.7 | 371.4 | 22.8 | 65.1 * | 46.1 * | 0.10 |
| 2.0 | 6.38 * | 1.71 * | 0.12 | 14.3 | 566.4 | 51.6 * | 139.5 * | 65.3 * | 0.16 |
| 5.0 | 7.33 * | 2.24 * | 0.14 | 13.6 | 574.1 | 58.2 * | 223.0 * | 93.2 * | 0.19 |
| 10 | 8.01 * | 3.12 * | 0.19 | 13.8 | 899.8 * | 112.3 * | 437.4 * | 213.3 * | 0.33* |
| Retisol loam | |||||||||
| 0 | 5.77 | 3.96 | 0.22 | 162.0 | 2856 | 164.1 | 67.2 | 379.7 | 0.69 |
| 0.5 | 5.56 | 4.71 * | 0.23 | 172.0 | 3090 | 253.6 * | 104.9 * | 270.5 | 0.74 |
| 2.0 | 5.80 | 4.94 * | 0.21 | 172.5 | 2956 | 284.6 * | 177.3 * | 357.7 | 0.76 |
| 5.0 | 5.94 * | 5.44 * | 0.21 | 174.7 | 2975 | 290.3 * | 186.3 * | 462.6 * | 0.78 |
| 10 | 7.40 * | 5.50 * | 0.20 | 165.5 | 2511 | 316.2 * | 371.6 * | 472.4 * | 0.73 |
| Retisol clay | |||||||||
| 0 | 6.30 | 0.48 | 0.18 | 105.0 | 630 | 194.9 | 99.1 | 84.2 | 0.20 |
| 0.5 | 6.89 * | 0.66 | 0.23 | 111.5 | 671 | 191.8 | 96.7 | 78.4 | 0.21 |
| 2.0 | 7.47 * | 1.48 * | 0.21 | 107.5 | 742 * | 177.0 | 155.2 * | 79.7 | 0.23 |
| 5.0 | 8.11 * | 1.76 * | 0.21 | 128.3 | 1073 * | 271.3 * | 229.2 * | 47.9 | 0.32 |
| 10 | 8.62 * | 2.78 * | 0.28 | 122.6 | 1260 * | 249.5 * | 379.5 * | 92.7 * | 0.40* |
| Histosol | |||||||||
| 5.49 | 35.93 | 1.80 | 42.3 | 8914 | 635.8 | 81.2 | 53.2 | 1.94 | |
| 0.5 | 5.34 | 36.51 | 1.68 | 45.5 | 8849 | 626.1 | 82.7 | 48.4 | 1.92 |
| 2.0 | 5.39 | 35.78 | 1.73 | 47.3 | 9357 | 682.5 | 232.3 * | 67.8 | 2.07 |
| 5.0 | 5.54 | 36.31 | 1.65 | 65.3 * | 8404 | 682.3 | 283.2 * | 108.9 * | 1.90 |
| 10 | 5.71 * | 36.93 | 1.80 | 60.1 * | 9316 | 832.8 * | 633.7 * | 180.3 * | 2.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yurkevich, M.; Suleymanov, R.; Ikkonen, E.; Dorogaya, E.; Bakhmet, O. Effect of Brown Algae (Fucus vesiculosus L.) on Humus and Chemical Properties of Soils of Different Type and Postgermination Growth of Cucumber Seedlings. Agronomy 2022, 12, 1991. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12091991
Yurkevich M, Suleymanov R, Ikkonen E, Dorogaya E, Bakhmet O. Effect of Brown Algae (Fucus vesiculosus L.) on Humus and Chemical Properties of Soils of Different Type and Postgermination Growth of Cucumber Seedlings. Agronomy. 2022; 12(9):1991. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12091991
Chicago/Turabian StyleYurkevich, Marija, Ruslan Suleymanov, Elena Ikkonen, Ekaterina Dorogaya, and Olga Bakhmet. 2022. "Effect of Brown Algae (Fucus vesiculosus L.) on Humus and Chemical Properties of Soils of Different Type and Postgermination Growth of Cucumber Seedlings" Agronomy 12, no. 9: 1991. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12091991