
Yield Potential of Machine-Transplanted Rice and Correlation of Crop-Growing Rate during Grain-Filling Stage
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Sites
2.2. Experimental Cultivars
2.3. Experimental Design
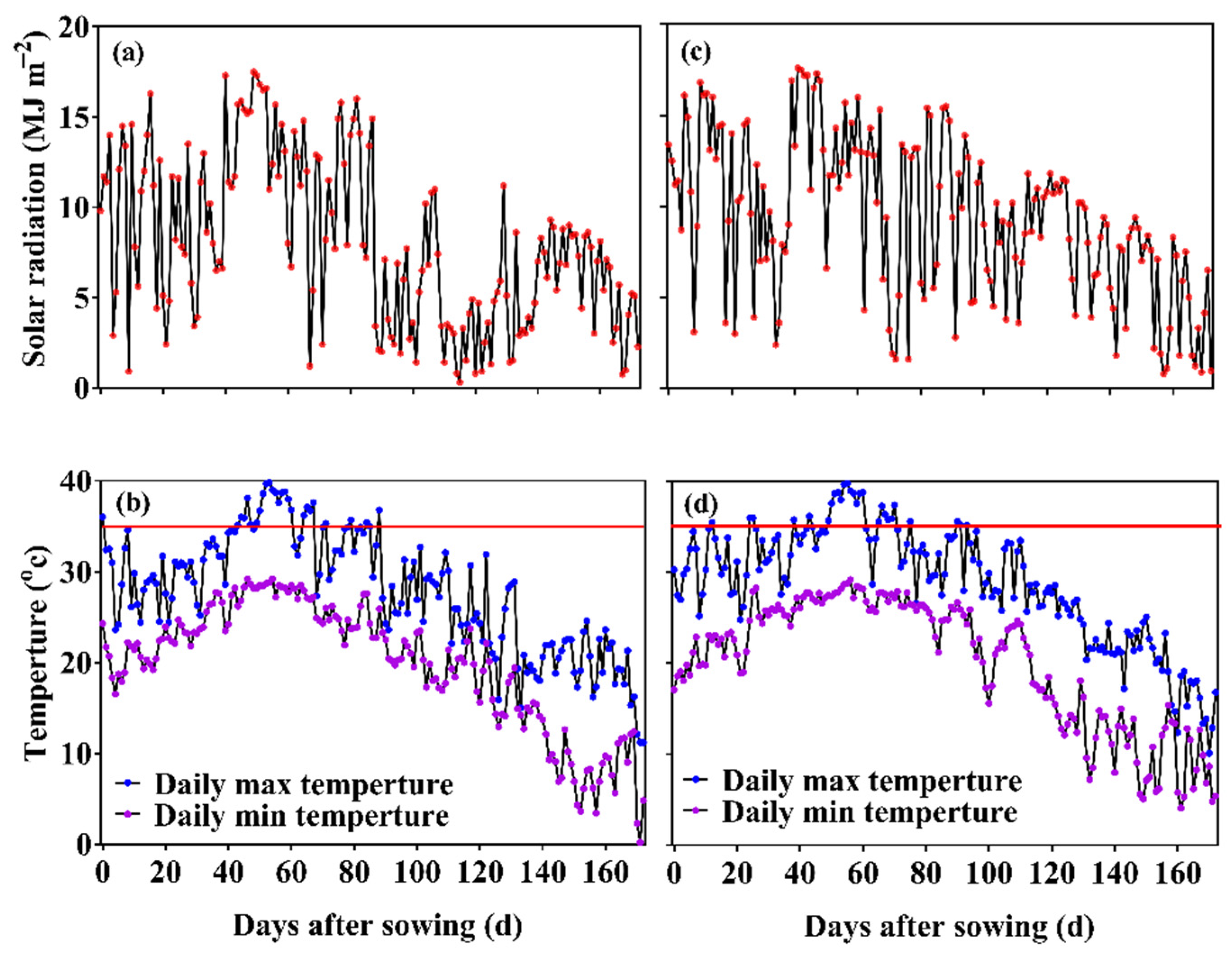
2.4. Observations and Measurements
2.5. Statistics Analyses
3. Results
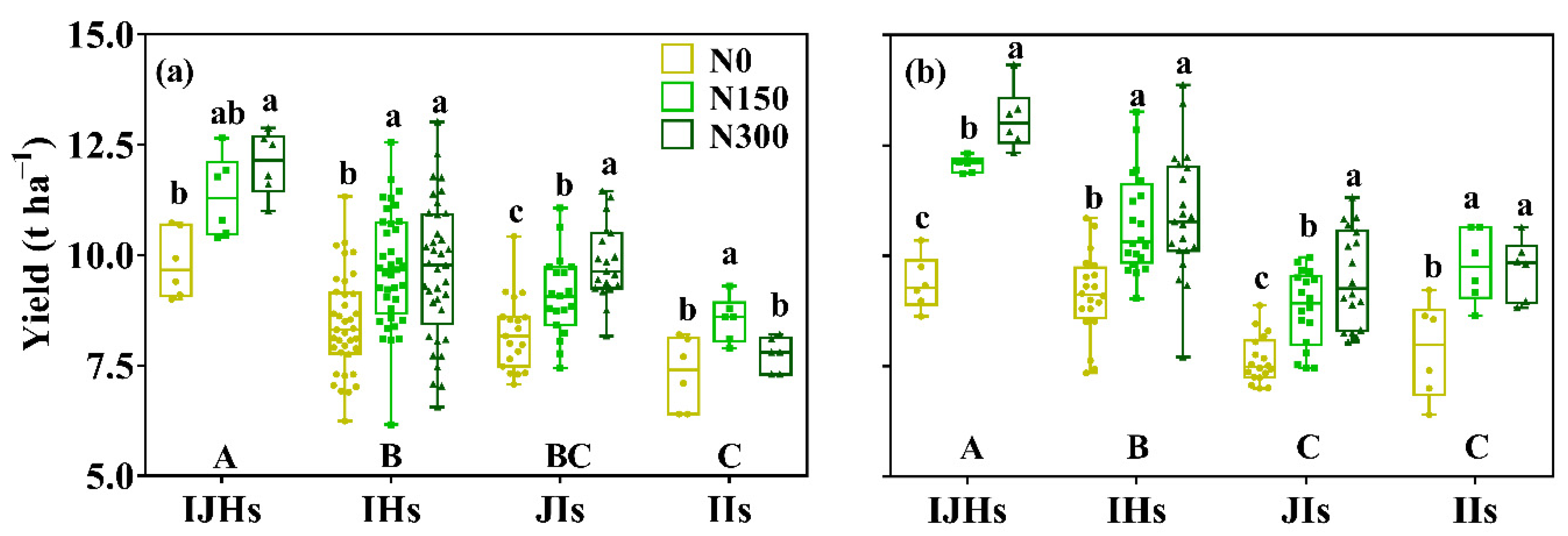
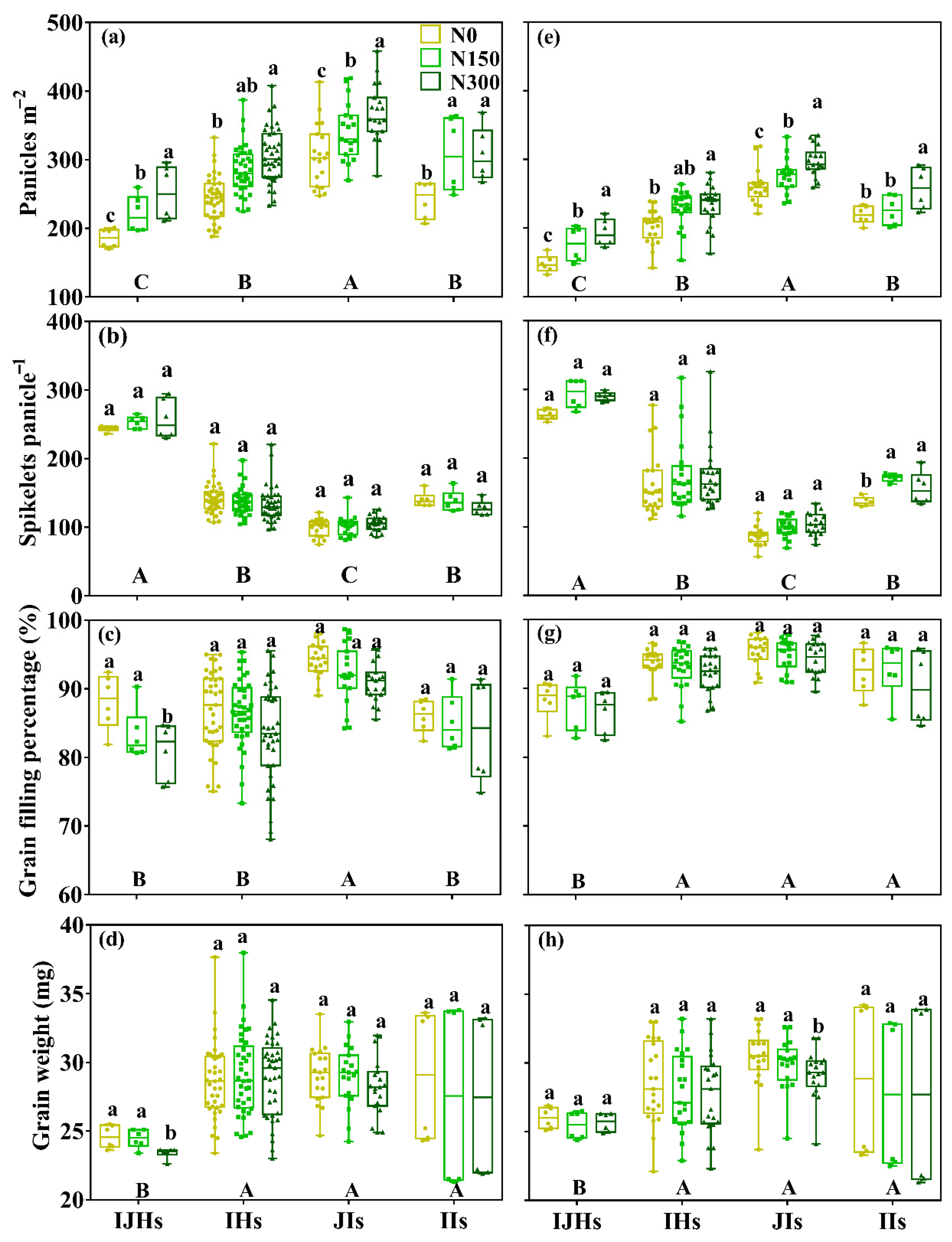
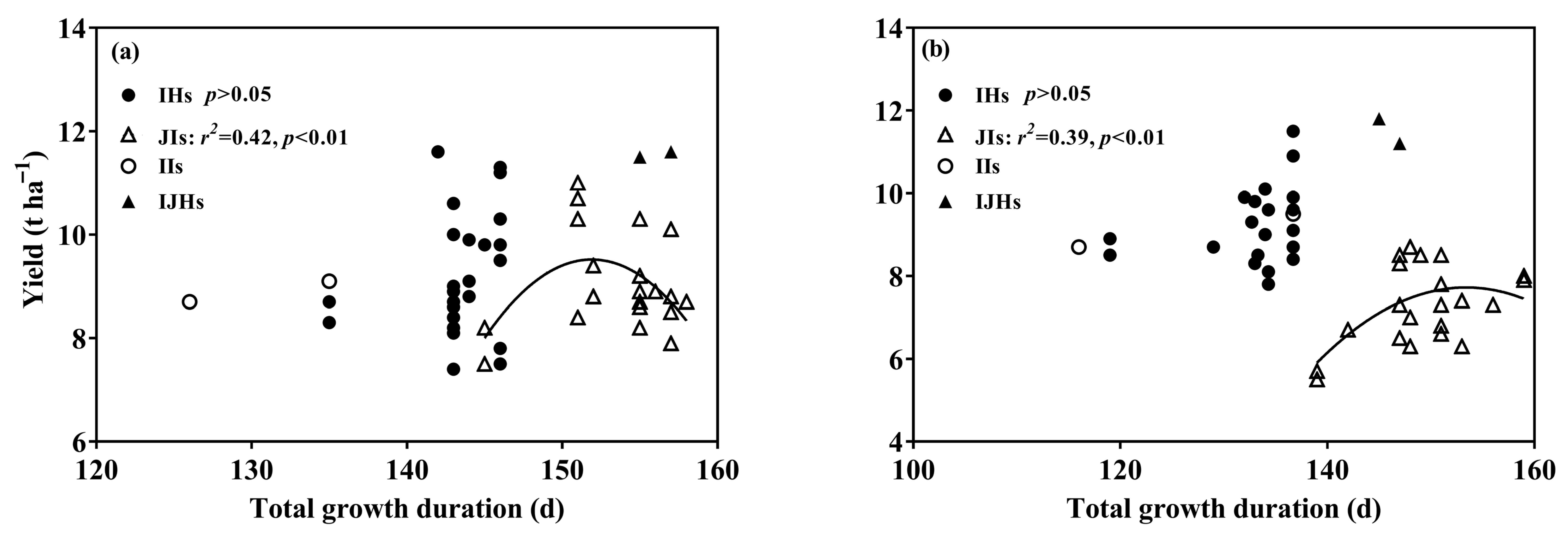
3.1. Yield and Its Components
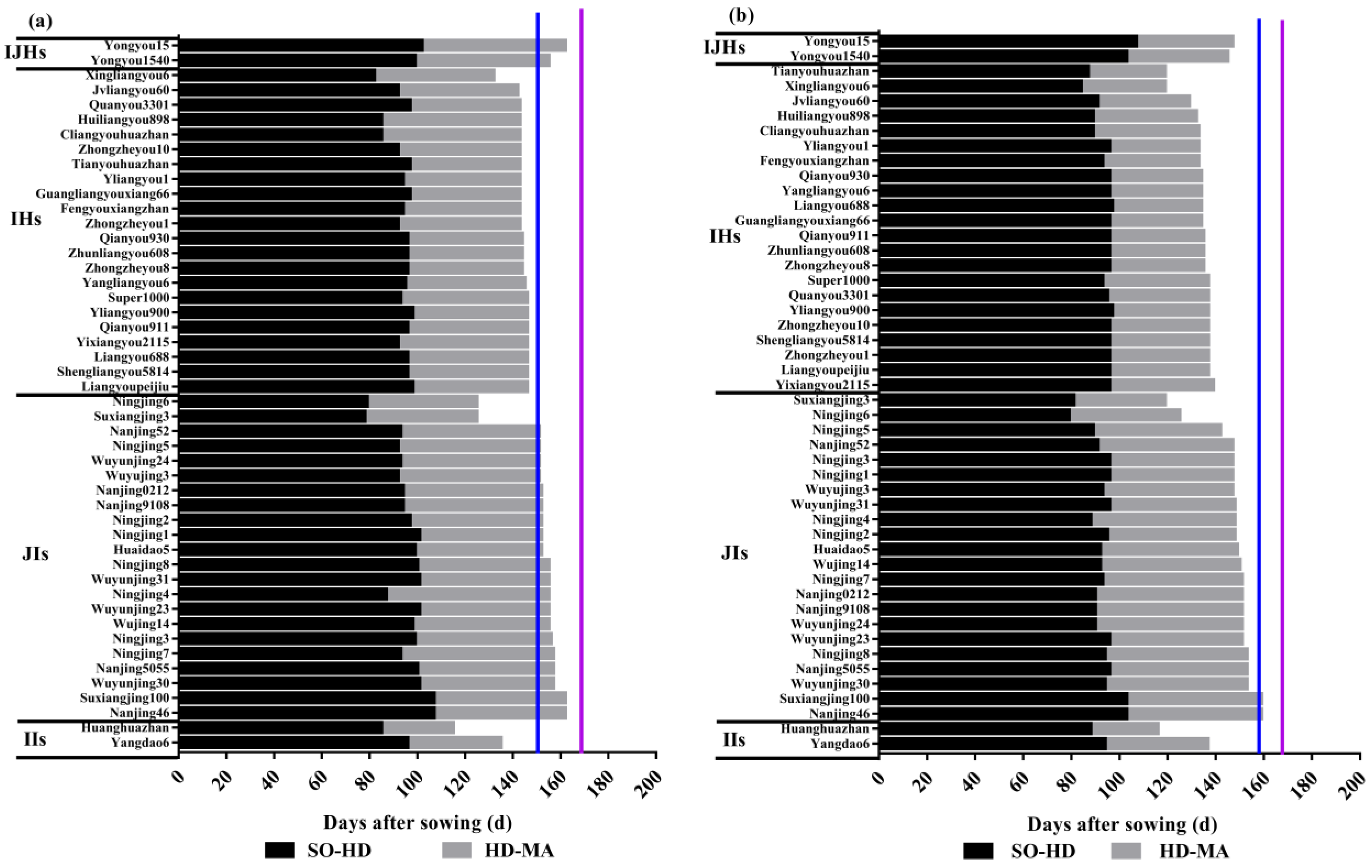
3.2. Growth Duration
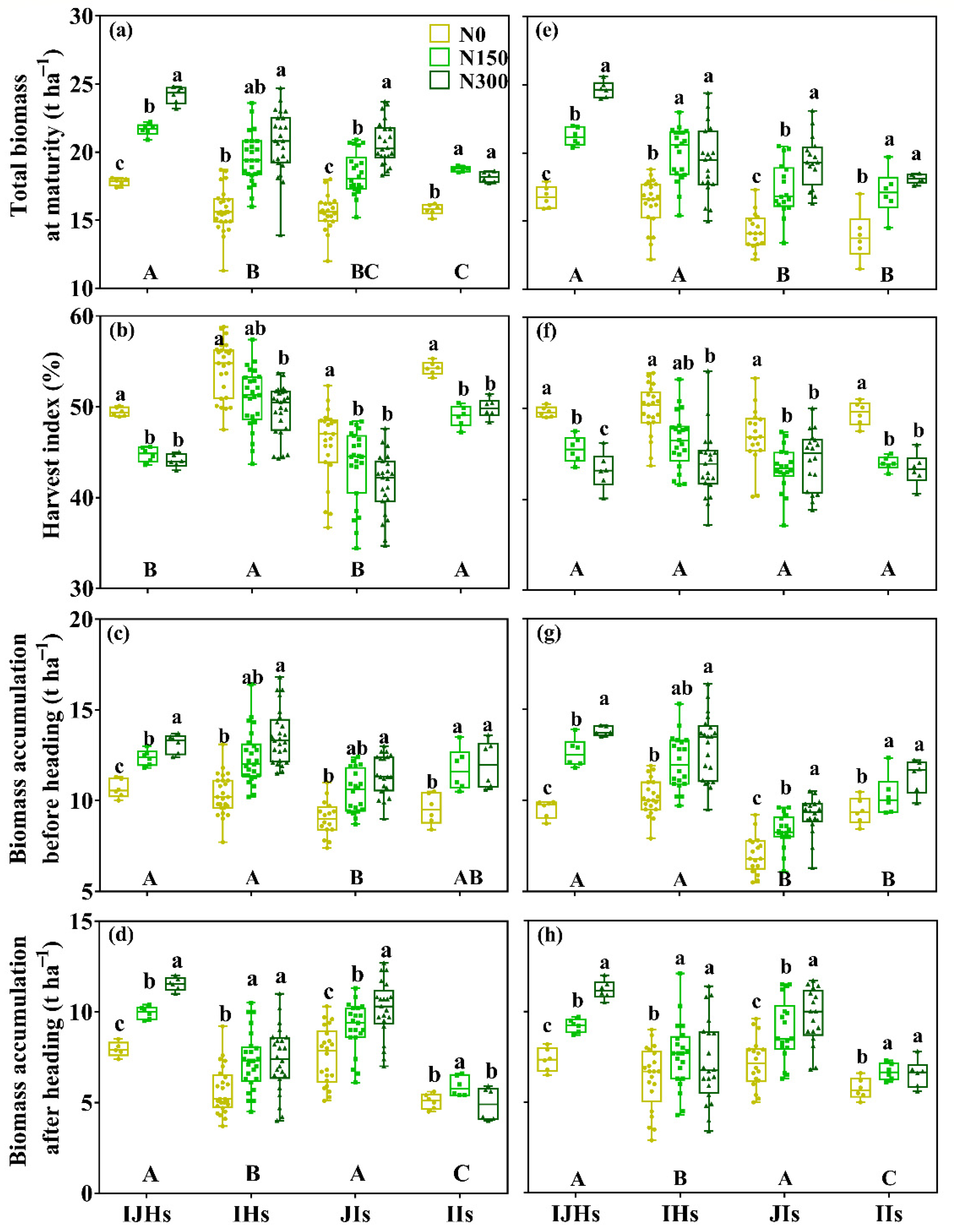
3.3. Biomass Accumulation
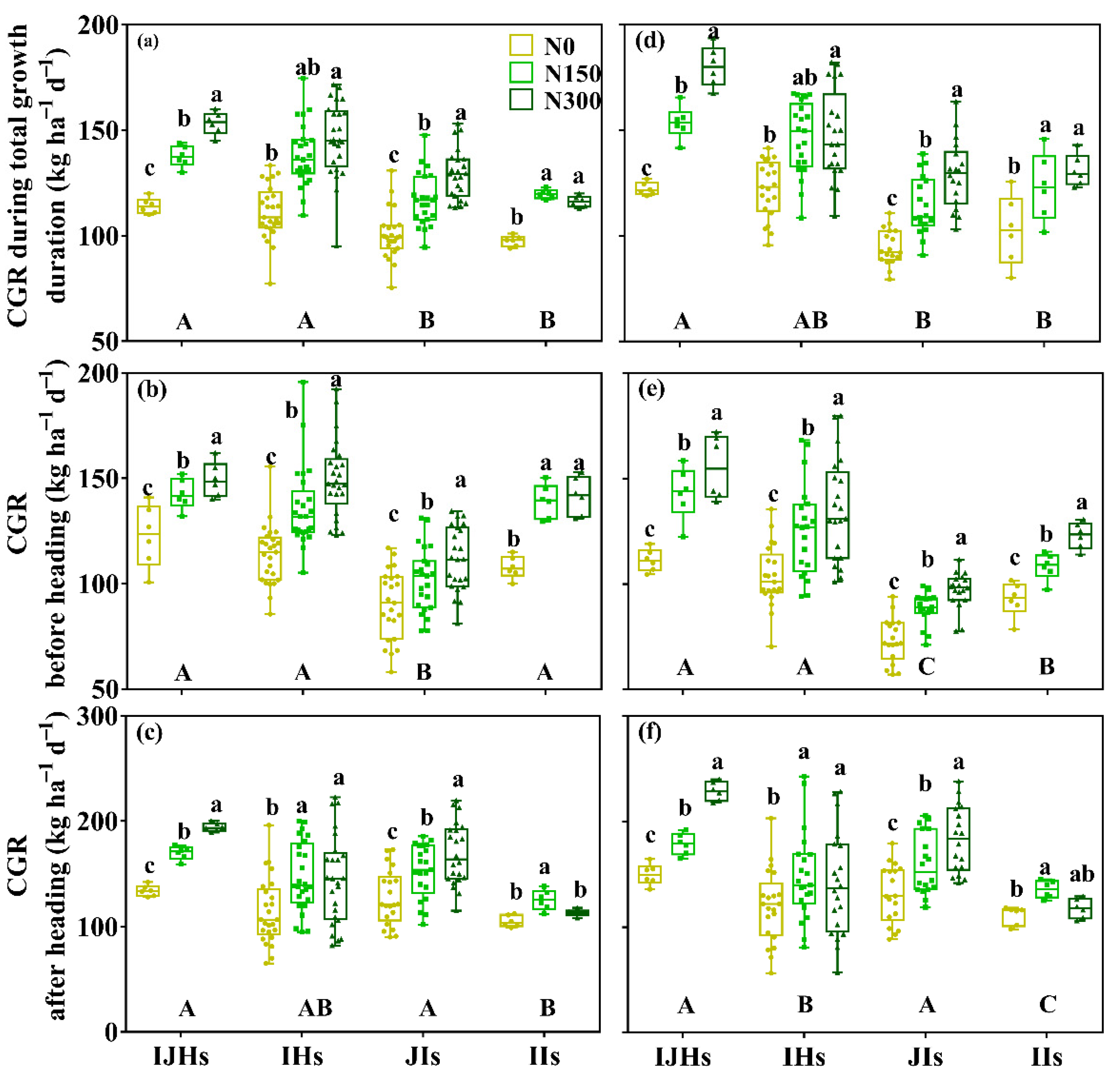
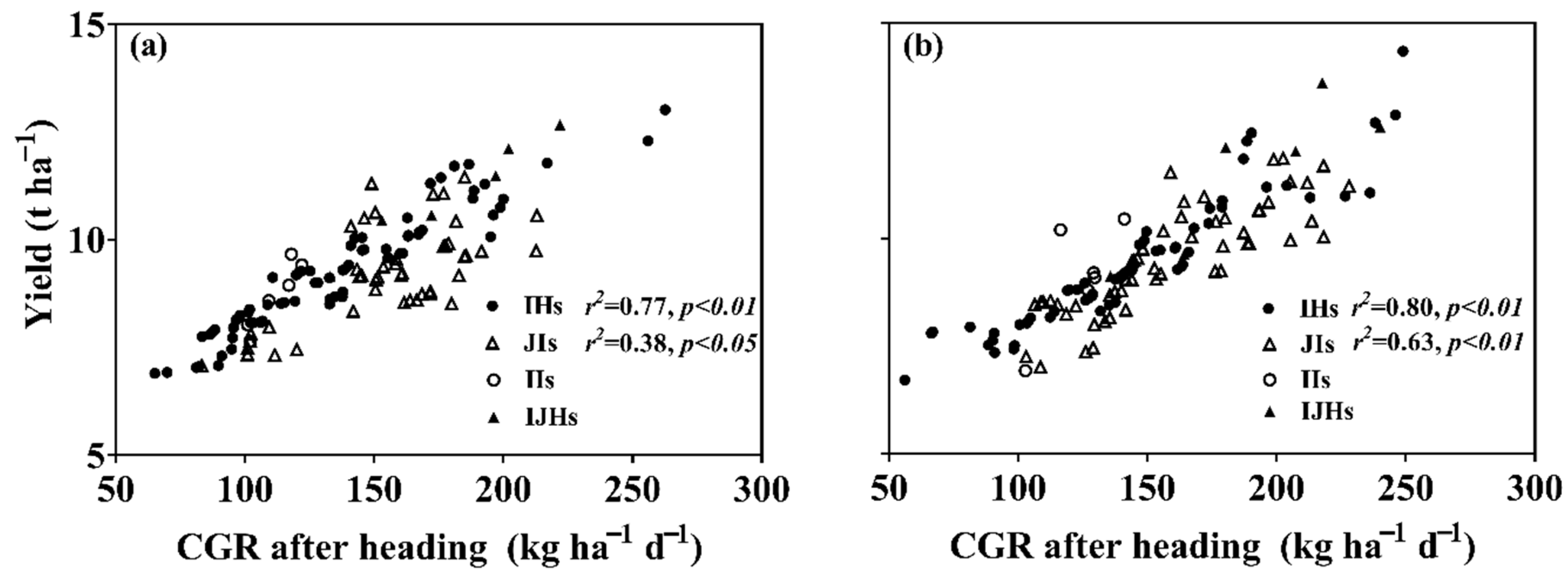
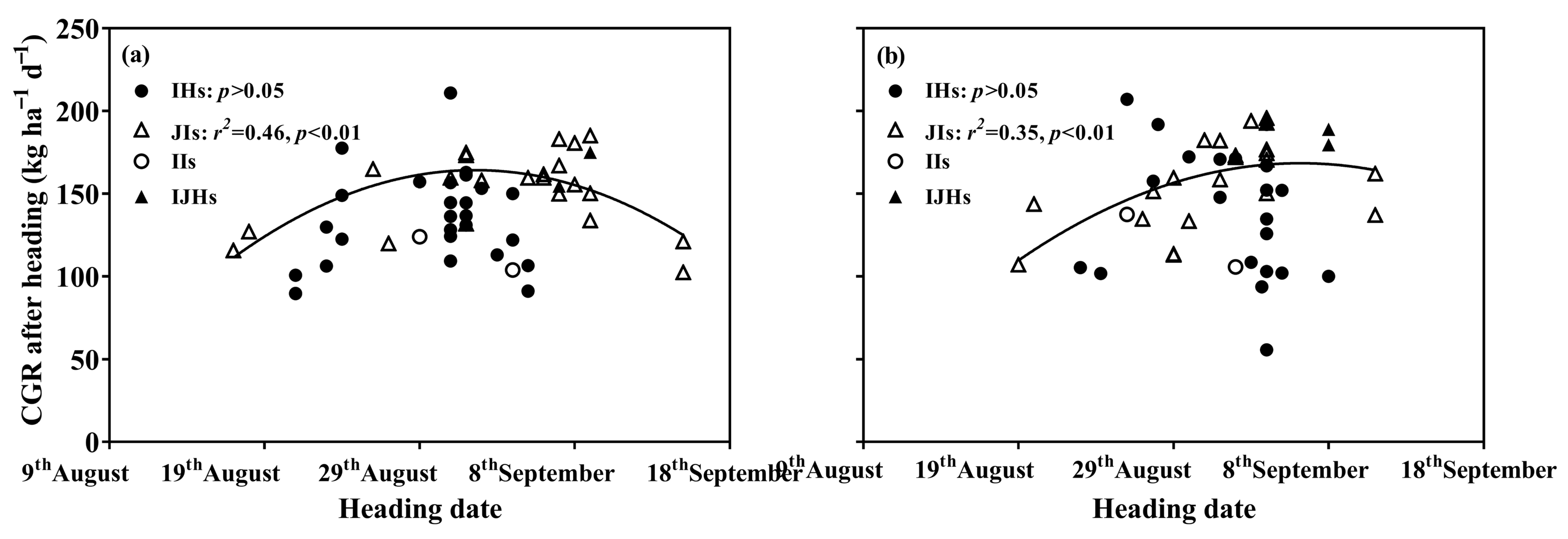
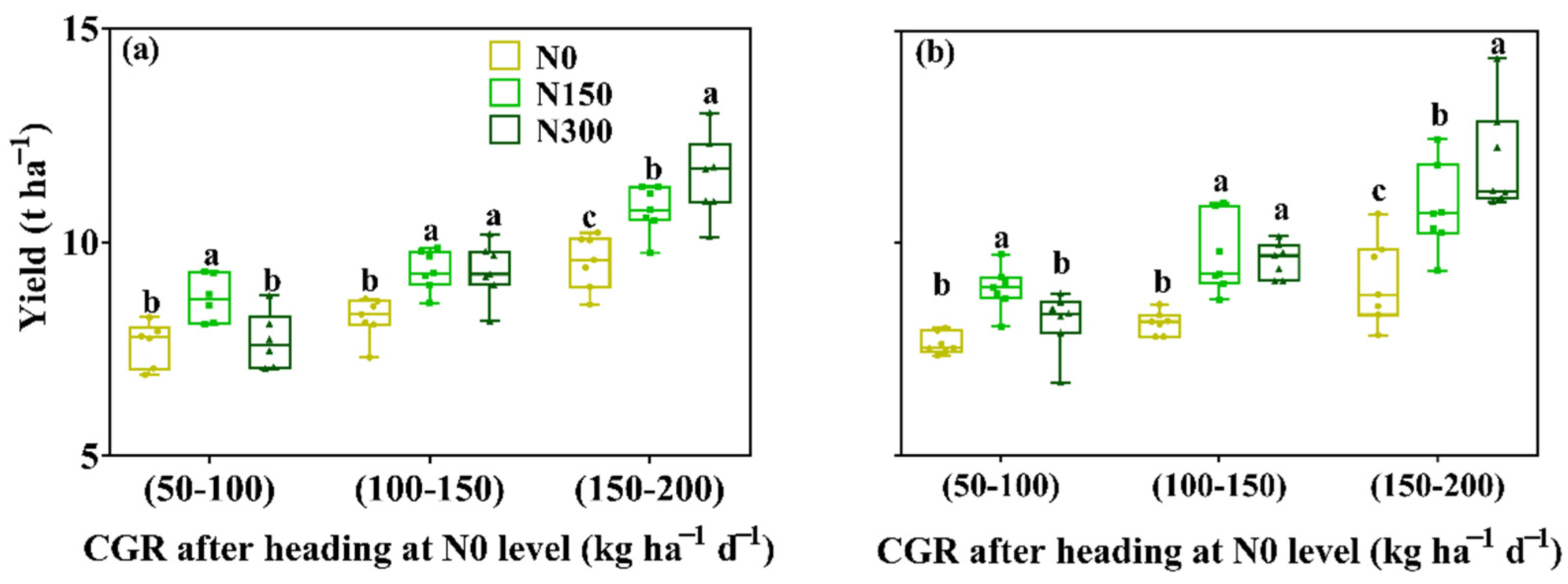
3.4. Crop-Growing Rate (CGR)
4. Discussion
4.1. Yield Differences among Groups of Rice
4.2. Yield Gap in IHs or JIs
4.3. Difference in Nitrogen Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Huang, M.; Zou, Y. Integrating mechanization with agronomy and breeding to ensure food security in China. Field Crops Res. 2018, 224, 22–27. [Google Scholar] [CrossRef]
- Ju, C.; Buresh, R.J.; Wang, Z.; Zhang, H.; Liu, L.; Yang, J.; Zhang, J. Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application. Field Crops Res. 2015, 175, 47–55. [Google Scholar] [CrossRef]
- Liu, X.; Xu, S.; Zhang, J.; Ding, Y.; Li, G.; Wang, S.; Liu, Z.; Tang, S.; Ding, C.; Chen, L. Effect of continuous reduction of nitrogen application to a rice-wheat rotation system in the middle-lower Yangtze River region (2013–2015). Field Crops Res. 2016, 196, 348–356. [Google Scholar] [CrossRef]
- Bai, H.; Tao, F. Sustainable intensification options to improve yield potential and eco-efficiency for rice-wheat rotation system in China. Field Crops Res. 2017, 211, 89–105. [Google Scholar] [CrossRef]
- Chen, S.; Liu, S.; Zheng, X.; Yin, M.; Chu, G.; Xu, C.; Yan, J.; Chen, L.; Wang, D.; Zhang, X. Effect of various crop rotations on rice yield and nitrogen use efficiency in paddy–upland systems in southeastern China. Crop J. 2018, 6, 576–588. [Google Scholar] [CrossRef]
- Yang, J.C.; Du, Y.; Liu, H. Cultivation approaches and techniques for annual super-high-yielding of rice and wheat in the lower reaches of Yangtze River. Sci. Agric. Sin. 2008, 41, 1611–1621. (In Chinese) [Google Scholar]
- Gong, J.; Zhang, H.; Li, J.; Huo, Z.; Xu, K.; Li, D.; Liu, G. Progress in research on patterns and systematic theories of super-high-yielding cultivation in rice. Chin. J. Rice Sci. 2010, 24, 417–424. (In Chinese) [Google Scholar]
- Li, H.; Liu, L.; Wang, Z.; Yang, J.; Zhang, J. Agronomic and physiological performance of high-yielding wheat and rice in the lower reaches of Yangtze River of China. Field Crops Res. 2012, 133, 119–129. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, J. Research status and development discussion on high-yielding agronomy of mechanized planting rice in China. Sci. Agric. Sin. 2014, 47, 1273–1289. (In Chinese) [Google Scholar]
- Zhu, D.F.; Zhang, Y.P.; Chen, H.Z.; Xiang, J.; Zhang, Y.K. Innovation and practice of high-yield rice cultivation technology in China. Sci. Agric. Sin. 2015, 48, 3404–3414. (In Chinese) [Google Scholar]
- Xing, Z.; Hu, Y.; Qian, H.; Cao, W.; Guo, B.; Wei, H.; Xu, K.; Huo, Z.; Zhou, G.; Dai, Q.; et al. Comparison of yield traits in rice among three mechanized planting methods in a rice-wheat rotation system. J. Integr. Agric. 2017, 16, 1451–1466. [Google Scholar] [CrossRef]
- Hu, Y.J.; Wu, P.; Zhang, H.C.; Dai, Q.G.; Huo, Z.Y.; Xu, K.; Gao, H.; Wei, H.Y.; Guo, B.W.; Cui, P. Comparison of agronomic performance between inter-sub-specific hybrid and inbred japonica rice under different mechanical transplanting methods. J. Integr. Agric. 2018, 17, 806–816. [Google Scholar] [CrossRef]
- Chen, S.; Ge, Q.; Chu, G.; Xu, C.; Yan, J.; Zhang, X.; Wang, D. Seasonal differences in the rice grain yield and nitrogen use efficiency response to seedling establishment methods in the Middle and Lower reaches of the Yangtze River in China. Field Crops Res. 2017, 205, 157–169. [Google Scholar] [CrossRef]
- Xing, Z.; Wu, P.; Zhu, M.; Qian, H.; Hu, Y.; Guo, B.; Wei, H.; Xu, K.; Huo, Z.; Dai, Q.; et al. Temperature and solar radiation utilization of rice for yield formation with different mechanized planting methods in the lower reaches of the Yangtze River, China. J. Integr. Agric. 2017, 16, 1923–1935. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Cao, F.; Yin, X.; Huang, M.; Zou, Y. Yield performance of early-season rice cultivars grown in the late season of double-season crop production under machine-transplanted conditions. PLoS ONE 2019, 14, e213075. [Google Scholar] [CrossRef]
- Zhang, Y.; Tang, Q.; Zou, Y.; Li, D.; Qin, J.; Yang, S.; Chen, L.; Xia, B.; Peng, S. Yield potential and radiation use efficiency of “super” hybrid rice grown under subtropical conditions. Field Crops Res. 2009, 114, 91–98. [Google Scholar] [CrossRef]
- Yuan, L.P. Progress in super-hybrid rice breeding. Crop J. 2017, 5, 100–102. [Google Scholar] [CrossRef]
- Huang, M.; Tang, Q.Y.; Ao, H.J.; Zou, Y. Yield potential and stability in super hybrid rice and its production strategies. J. Integr. Agric. 2017, 16, 1009–1017. [Google Scholar] [CrossRef]
- Shi, X.; Hu, R. Rice variety improvement and the contribution of foreign germplasms in China. J. Integr. Agric. 2017, 16, 2337–2345. [Google Scholar] [CrossRef]
- Tang, L.; Xu, Z.; Chen, W. Advances and prospects of super rice breeding in China. J. Integr. Agric. 2017, 16, 984–991. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, Y.; Zhu, Z.; Chen, T.; Zhao, Q.; Zhong, W.; Yang, J.; Yao, S.; Zhou, L.; Zhao, L.; et al. Research progress on the breeding of japonica super rice varieties in Jiangsu Province, China. J. Integr. Agric. 2017, 16, 992–999. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, H.; Blumwald, E.; Li, H.; Cheng, J.; Dai, Q.; Huo, Z.; Xu, K.; Guo, B. Different characteristics of high yield formation between inbred japonica super rice and inter-sub-specific hybrid super rice. Field Crops Res. 2016, 198, 179–187. [Google Scholar] [CrossRef]
- Wei, H.; Meng, T.; Li, X.; Dai, Q.; Zhang, H.; Yin, X. Sink-source relationship during rice grain filling is associated with grain nitrogen concentration. Field Crops Res. 2018, 215, 23–38. [Google Scholar] [CrossRef]
- Wei, H.; Meng, T.; Li, C.; Xu, K.; Huo, Z.; Wei, H.; Guo, B.; Zhang, H.; Dai, Q. Comparisons of grain yield and nutrient accumulation and translocation in high-yielding japonica/indica hybrids, indica hybrids, and japonica conventional varieties. Field Crops Res. 2017, 204, 101–109. [Google Scholar] [CrossRef]
- Jiang, Y.H.; Xu, K.; Zhao, K.; Sun, J.J.; Wei, H.H.; Xu, J.W.; Wei, H.Y.; Guo, B.W.; Huo, Z.Y.; Dai, Q.G.; et al. Canopy structure and photosynthetic characteristics of Yongyou series of indica-japonica hybrid rice under high-yielding cultivation condition. Acta Agron. Sin. 2015, 41, 286–296. (In Chinese) [Google Scholar] [CrossRef]
- Jiang, P.; Xie, X.; Huang, M.; Zhou, X.; Zhang, R.; Chen, J.; Wu, D.; Xia, B.; Xiong, H.; Xu, F.; et al. Potential Yield Increase of Hybrid Rice at Five Locations in Southern China. Rice 2016, 9, 11. [Google Scholar] [CrossRef]
- Koutroubas, S.D.; Ntanos, D.A. Genotypic differences for grain yield and nitrogen utilization in Indica and Japonica rice under Mediterranean conditions. Field Crops Res. 2003, 83, 251–260. [Google Scholar] [CrossRef]
- Gong, J.; Xing, Z.; Hu, Y.; Zhang, H.; Huo, Z.; Xu, K.; Gao, H. Difference of characteristics of photosynthesis, matter production and translocation between indica and japonica super rice. Acta Agron. Sin. 2014, 40, 497–510. (In Chinese) [Google Scholar] [CrossRef]
- Okamura, M.; Arai-Sanoh, Y.; Yoshida, H.; Mukouyama, T.; Adachi, S.; Yabe, S.; Nakagawa, H.; Tsutsumi, K.; Taniguchi, Y.; Kobayashi, N.; et al. Characterization of high-yielding rice cultivars with different grain-filling properties to clarify limiting factors for improving grain yield. Field Crops Res. 2018, 219, 139–147. [Google Scholar] [CrossRef]
- Liu, Q.; Hu, J.F.; Zhou, W.; Yang, Z.P.; Chen, Y.; Ren, W.J. Dry matter production and yield characteristics of machine-transplanted rice varieties falling into different types in sichuan basin. Chin. J. Rice Sci. 2019, 33, 35–46. (In Chinese) [Google Scholar]
- Li, M.; Zhang, H.; Yang, X.; Ge, M.; Ma, Q.; Wei, H.; Dai, Q.; Huo, Z.; Xu, K.; Luo, D. Accumulation and utilization of nitrogen, phosphorus and potassium of irrigated rice cultivars with high productivities and high N use efficiencies. Field Crops Res. 2014, 161, 55–63. [Google Scholar] [CrossRef]
- Liu, L.J.; Wang, K.J.; Bian, J.L.; Xiong, Y.W.; Chen, L.; Wang, Z.Q.; Yang, J.C. Differences in yield response to nitrogen fertilizer among rice cultivars and their relationship with root morphology and physiology. Acta Agron. Sin. 2014, 40, 1999–2007. (In Chinese) [Google Scholar] [CrossRef]
- Jiang, P.; Xie, X.; Huang, M.; Zhou, X.; Zhang, R.; Chen, J.; Wu, D.; Xia, B.; Xu, F.; Xiong, H.; et al. Comparisons of yield performance and nitrogen response between hybrid and inbred rice under different ecological conditions in southern China. J. Integr. Agric. 2015, 14, 1283–1294. [Google Scholar] [CrossRef]
- Huang, L.; Sun, F.; Yuan, S.; Peng, S.B.; Wang, F. Different mechanisms underlying the yield advantage of ordinary hybrid and super hybrid rice over inbred rice under low and moderate N input conditions. Field Crops Res. 2018, 216, 150–157. [Google Scholar] [CrossRef]
- Yin, C.Y.; Wei, H.Y.; Zhang, Q.; Dai, Q.G.; Huo, Z.Y.; Ke, X. Differences and correlations in grain yield, n uptake and utilization between medium-maturing indica and japonica rice under different n fertilizer levels. Acta Agron. Sin. 2009, 35, 348–355. (In Chinese) [Google Scholar] [CrossRef]
- Meng, T.Y.; Xu, J.W.; Shao, Z.B.; Ge, M.J.; Zhang, H.C.; Wei, H.Y.; Jing, P.P. Advantages and their formation characteristics of the highest population productivity of nitrogen fertilization in japonica/indica hybrid rice of Yongyou series. Acta Agron. Sin. 2015, 41, 1711–1725. (In Chinese) [Google Scholar] [CrossRef]
- Tabien, R.E.; Samonte, S.O.P.; McClung, A.M. Forty-eight years of rice improvement in Texas since the release of cultivar Bluebonnet in 1944. Crop Sci. 2008, 48, 2097–2106. [Google Scholar] [CrossRef]
- Yang, J.; Peng, S.; Zhang, Z.; Wang, Z.; Visperas, R.M.; Zhu, Q. Grain and dry matter yields and partitioning of assimilates in japonica/indica hybrid rice. Crop Sci. 2002, 42, 766–772. [Google Scholar] [CrossRef]
- Guo, B.; Hua, J.; Zhou, N.; Zhang, H.; Chen, B.; Shu, P.; Huo, Z.; Zhou, P.; Cheng, F.; Huang, D.; et al. Difference in yield and population characteristics of different types of late rice cultivars in double-cropping rice area. Acta Agron. Sin. 2015, 41, 1220–1236. (In Chinese) [Google Scholar] [CrossRef]
- Hua, J.; Zhou, N.; Zhang, J.; Zhang, H.; Huo, Z.; Zhou, P.; Cheng, F.; Li, G.; Huang, D.; Chen, Z.; et al. Selection of late rice cultivars of japonica rice switched from indica rice in double cropping rice area. Sci. Agric. Sin. 2014, 47, 4582–4594. (In Chinese) [Google Scholar]
- Bueno, C.S.; Lafarge, T. Higher crop performance of rice hybrids than of elite inbreds in the tropics: 1. Hybrids accumulate more biomass during each phenological phase. Field Crops Res. 2009, 112, 229–237. [Google Scholar] [CrossRef]
- Peng, S.; Huang, J.; Sheehy, J.E.; Laza, R.C.; Visperas, R.M.; Zhong, X.; Centeno, G.S.; Khush, G.S.; Cassman, K.G. Rice yield decline with higher night temperature from global warming. Proc. Natl. Acad. Sci. USA 2004, 101, 9971–9997. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Laza, M.R.C.; Cassman, K.G.; Huang, J.; Nie, L.; Ling, X.; Centeno, G.S.; Cui, K.; Wang, F.; Li, Y.; et al. Temperature explains the yield difffference of doubleseason rice between tropical and subtropical environments. Field Crops Res. 2016, 198, 303–311. [Google Scholar] [CrossRef]
- Chang, S.; Chang, T.; Song, Q.; Zhu, X.; Deng, Q. Photosynthetic and agronomic traits of an elite hybrid rice Y-Liang-You 900 with a record-high yield. Field Crops Res. 2016, 187, 49–57. [Google Scholar] [CrossRef]
- Li, G.; Yang, C. Design and Practice of Precise Quantitative Cultivation Techniques for Super-High Yield of Rice; China Agriculture Press: Beijing, China, 2019. [Google Scholar]
- Takai, T.; Matsuura, S.; Nishio, T.; Ohsumi, A.; Shiraiwa, T.; Horie, T. Rice yield potential is closely related to crop growth rate during late reproductive period. Field Crops Res. 2006, 96, 328–335. [Google Scholar] [CrossRef]
- Huang, L.; Yang, D.; Li, X.; Peng, S.B.; Wang, F. Coordination of high grain yield and high nitrogen use efficiency through large sink size and high post-heading source capacity in rice. Field Crops Res. 2019, 233, 49–58. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Bai, Q. Analysis of high wheat yields in northwest China. Agric. Syst. 1997, 53, 373–385. [Google Scholar] [CrossRef]
- Tashiro, T.; Wardlaw, I. The effect of high temperature on the accumulation of dry matter: Carbon and nitrogen in the kernel of rice. Funct. Plant Biol. 1991, 18, 259–265. [Google Scholar] [CrossRef]
- Deng, N.; Ling, X.; Sun, Y.; Zhang, C.; Fahad, S.; Peng, S.; Cui, K.; Nie, L.; Huang, J. Influence of temperature and solar radiation on grain yield and quality in irrigated rice system. Eur. J. Agron. 2015, 64, 37–46. [Google Scholar] [CrossRef]
- Zhou, Q.; Yuan, R.; Zhu, K.; Wang, Z.; Yang, J. Characteristics of grain yield and nitrogen absorption and utilization of indica/japonica hybrid rice Yongyou 2640 under different nitrogen application rates. Acta Agron. Sin. 2022, 48, 2285–2299. (In Chinese) [Google Scholar]
- Huang, M.; Jiang, P.; Shan, S.; Gao, W.; Ma, G.; Zou, Y.; Uphoff, N.; Yuan, L. Higher yields of hybrid rice do not depend on nitrogen fertilization under moderate to high soil fertility conditions. Rice 2017, 10, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Sun, Z.; Chen, Q.; Damaris, R.; Lu, B.; Hu, Z. Nitrogen fertilizer induced alterations in the root proteome of two rice cultivars. Int. J. Mol. Sci. 2019, 20, 3674. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Group | Year of | Planting Area | Cultivar | Group | Year of | Planting Area |
|---|---|---|---|---|---|---|---|
| Official Release | (×104 ha) | Official Release | (×104 ha) | ||||
| Liangyoupeijiu | IHs | 1999 | 604 | Wuyunjing7 | JIs | 1995 | 363 |
| Zhongzheyou1 | IHs | 1999 | 155 | Wujing14 | JIs | 1999 | 126 |
| Fengyouxiangzhan | IHs | 2001 | 49 | Ningjing1 | JIs | 2001 | 106 |
| Zhongzheyou8 | IHs | 2001 | 70 | Ningjing2 | JIs | 2001 | 13 |
| Xingliangyou6 | IHs | 2003 | 217 | Ningjing3 | JIs | 2001 | 34 |
| Guangliangyouxiang66 | IHs | 2005 | 50 | Nanjing46 | JIs | 2004 | 14 |
| Yliangyou1 | IHs | 2005 | 249 | Nanjing5055 | JIs | 2005 | 72 |
| Zhunliangyou608 | IHs | 2006 | 25 | Wuyunjing23 | JIs | 2005 | 99 |
| Shengliangyou5814 | IHs | 2007 | 157 | Wuyunjing24 | JIs | 2006 | 75 |
| Tianyouhuazhan | IHs | 2007 | 150 | Suxiangjing3 | JIs | 2006 | |
| Liangyou688 | IHs | 2008 | 41 | Ningjing4 | JIs | 2007 | 143 |
| Zhongzheyou10 | IHs | 2008 | 8 | Ningjing5 | JIs | 2007 | 5 |
| Yixiangyou2115 | IHs | 2009 | 53 | Ningjing6 | JIs | 2009 | 2 |
| Yangliangyou6 | IHs | 2009 | 287 | Nanjing52 | JIs | 2009 | 5 |
| Qianyou930 | IHs | 2009 | Nanjing9108 | JIs | 2009 | 111 | |
| Qianyou911 | IHs | 2010 | Nanjing0212 | JIs | 2010 | 3 | |
| Cliangyouhuazhan | IHs | 2010 | 63 | Wuyunjing31 | JIs | 2010 | 10 |
| Huiliangyou898 | IHs | 2011 | 10 | Suxiangjing100 | JIs | 2011 | |
| Jvliangyou60 | IHs | 2012 | Ningjing7 | JIs | 2011 | ||
| Yliangyou900 | IHs | 2013 | 25 | Ningjing8 | JIs | 2012 | |
| Quanyou3301 | IHs | 2013 | Yangdao6 | IIs | 1993 | 44 | |
| Chaoyou1000 | IHs | 2014 | Huanghuazhan | IIs | 2002 | 257 | |
| Wuyujing3 | JIs | 1990 | 608 | Yongyou1540 | IJHs | 2010 | 8 |
| Huaidao5 | JIs | 1994 | 175 | Yongyou538 | IJHs | 2013 | 29 |
| Traits | Group | Nitrogen | Year | G × N | G × Y | N × Y | G × N × Y |
|---|---|---|---|---|---|---|---|
| Yield | ** | ** | * | * | ns | ns | ns |
| Panicles m−2 | ** | ** | ** | ns | ns | ns | ns |
| Spikelets panicle−1 | ** | ns | ** | ns | ns | ns | ns |
| Grain filling percentage | ** | ** | ** | ns | * | ** | ns |
| Grain weight | ** | ** | ** | * | ns | * | ns |
| Total biomass at maturity | ** | ** | ** | ns | ns | ns | ns |
| Harvest index | ** | ** | ns | ns | * | ns | ns |
| Biomass accumulation before heading | ** | ** | ns | ns | ns | ns | ns |
| Biomass accumulation after heading | ** | ** | ** | ** | ns | ns | ns |
| CGR during the total growth duration | ** | ** | * | ns | ns | ns | ns |
| CGR before heading | ** | ** | ns | ns | ns | ns | ns |
| CGR after heading | * | ** | * | ** | ns | ns | ns |
| Traits | IJHs | IHs | JIs | IIs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | N | C × N | C | N | C × N | C | N | C × N | C | N | C × N | |
| Yield | ns | ** | ns | ** | ** | ** | ** | ** | ns | ** | * | ns |
| Panicles m−2 | ns | ** | ns | ** | ** | ns | ** | ** | ns | * | ** | ns |
| Spikelets panicle−1 | ns | ns | ns | ** | ** | ns | ** | ns | ns | ns | ns | ns |
| Grain filling percentage | ns | ** | ns | ** | ** | ** | ns | ** | ns | * | ns | ns |
| Grain weight | ns | ** | ns | ** | ** | ns | ** | ** | * | ** | ** | ns |
| Total biomass at maturity | ns | ** | ns | ** | ** | * | ** | ** | ns | ns | ** | ns |
| Harvest index | ns | ** | ns | ** | ** | ns | * | ** | ns | ns | ** | ns |
| Biomass accumulation before heading | ns | ** | ns | ** | ** | ns | * | ** | ns | ns | ** | ns |
| Biomass accumulation after heading | ns | ** | ns | ** | ** | ** | ** | ** | ns | ns | * | ns |
| CGR during the total growth duration | ns | ** | ns | ** | ** | ns | * | ** | ns | ns | ** | ns |
| CGR before heading | ns | ** | ns | ** | ** | ns | ns | ** | ns | ns | ** | ns |
| CGR after heading | ns | ** | ns | ** | ** | ** | ** | ** | ns | ns | * | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, C.; Zhu, X.; Xu, C.; Cambula, E.; Lu, B.; Luo, X.; Wu, Q.; Zhong, Q.; Xu, X.; Liu, Z.; et al. Yield Potential of Machine-Transplanted Rice and Correlation of Crop-Growing Rate during Grain-Filling Stage. Agronomy 2022, 12, 2299. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102299
Ding C, Zhu X, Xu C, Cambula E, Lu B, Luo X, Wu Q, Zhong Q, Xu X, Liu Z, et al. Yield Potential of Machine-Transplanted Rice and Correlation of Crop-Growing Rate during Grain-Filling Stage. Agronomy. 2022; 12(10):2299. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102299
Chicago/Turabian StyleDing, Chao, Xuhui Zhu, Congshan Xu, Elidio Cambula, Bo Lu, Xikun Luo, Qiong Wu, Qiuyi Zhong, Xia Xu, Zhenghui Liu, and et al. 2022. "Yield Potential of Machine-Transplanted Rice and Correlation of Crop-Growing Rate during Grain-Filling Stage" Agronomy 12, no. 10: 2299. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102299