
Screening Soybean for Adaptation to Relay Intercropping Systems: Associations between Reproductive Organ Abscission and Yield
Abstract
:1. Introduction
2. Materials and Methods
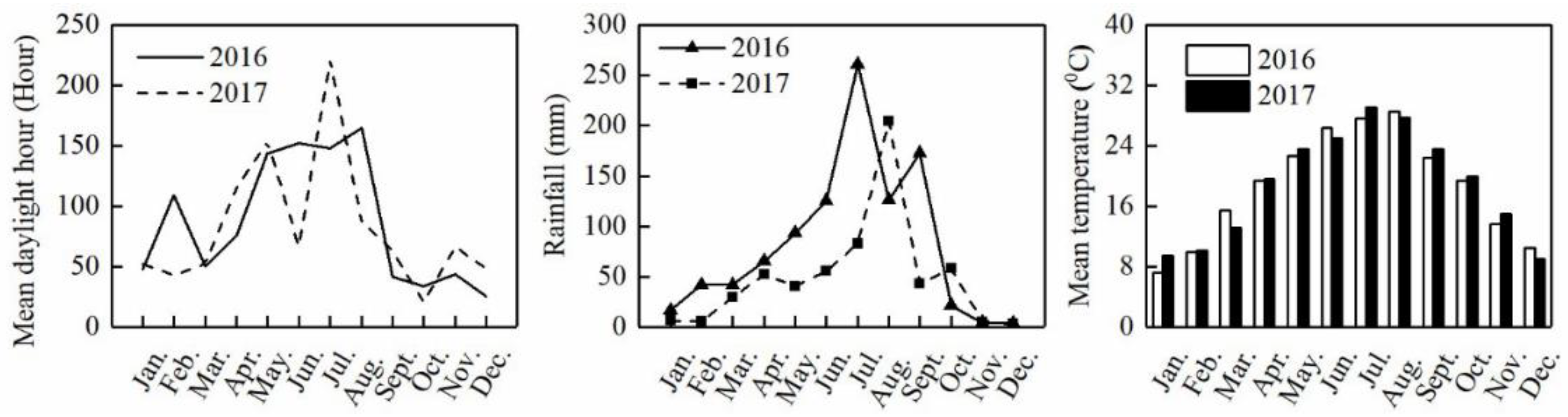
2.1. Experiment Sites
2.2. Experiment Design and Field Management
2.3. Plant Sampling and Measurements
2.4. Calculation and Statistical Analysis
3. Results
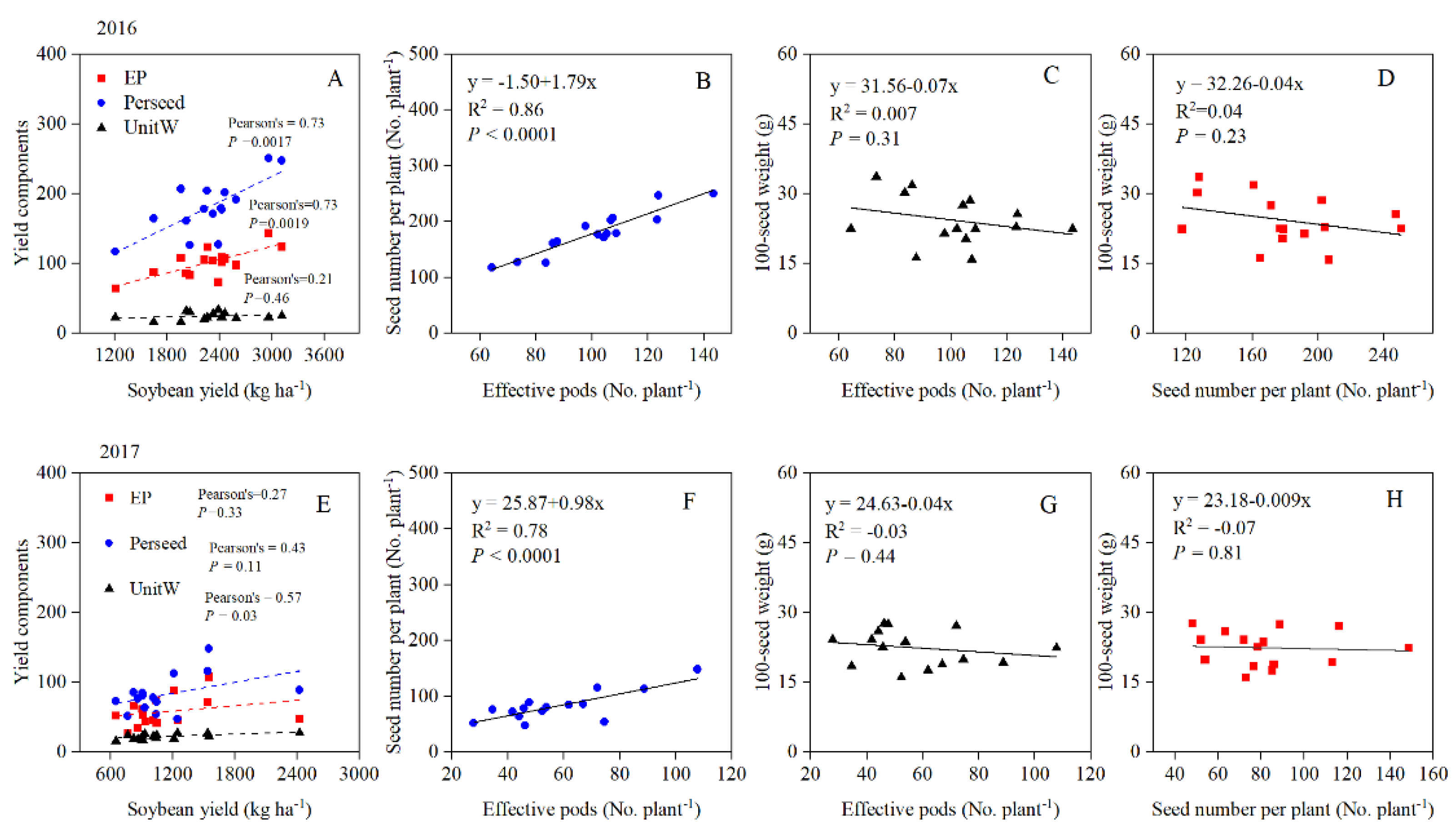
3.1. Yield and Yield Components
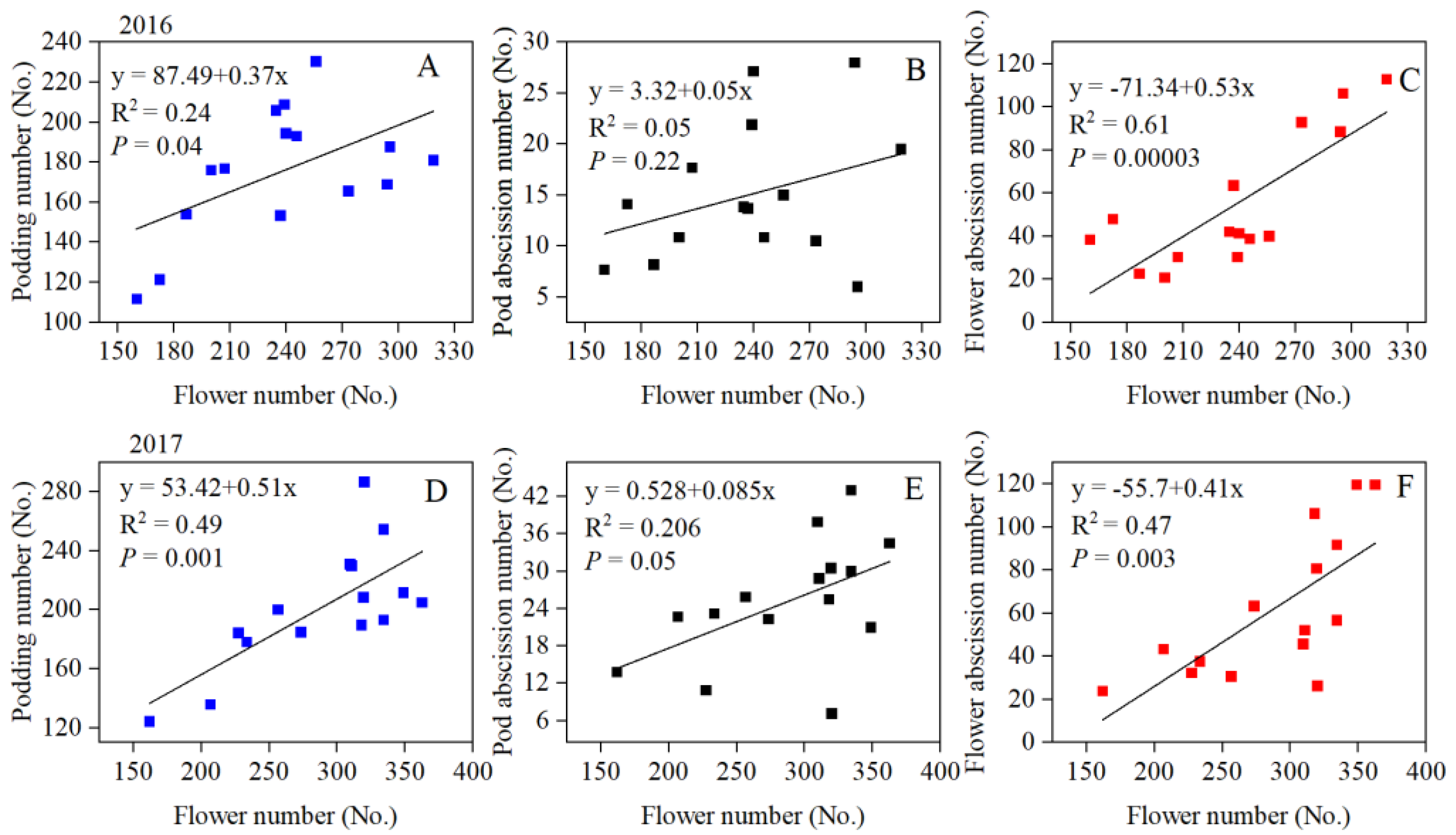
3.2. Flower and Pod Abscission Characters
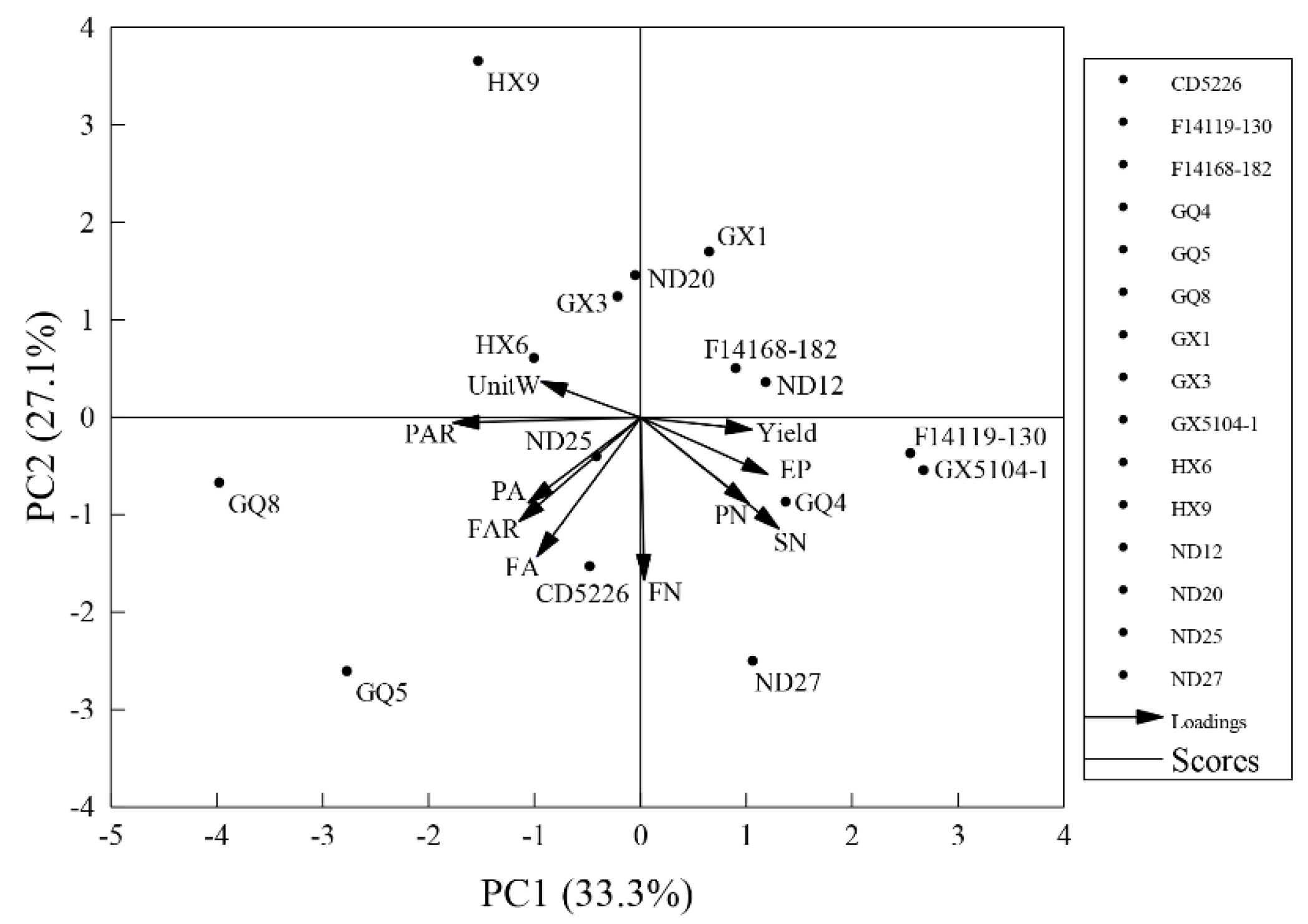
3.3. Principal Component Analysis (PCA) on Agronomic Traits
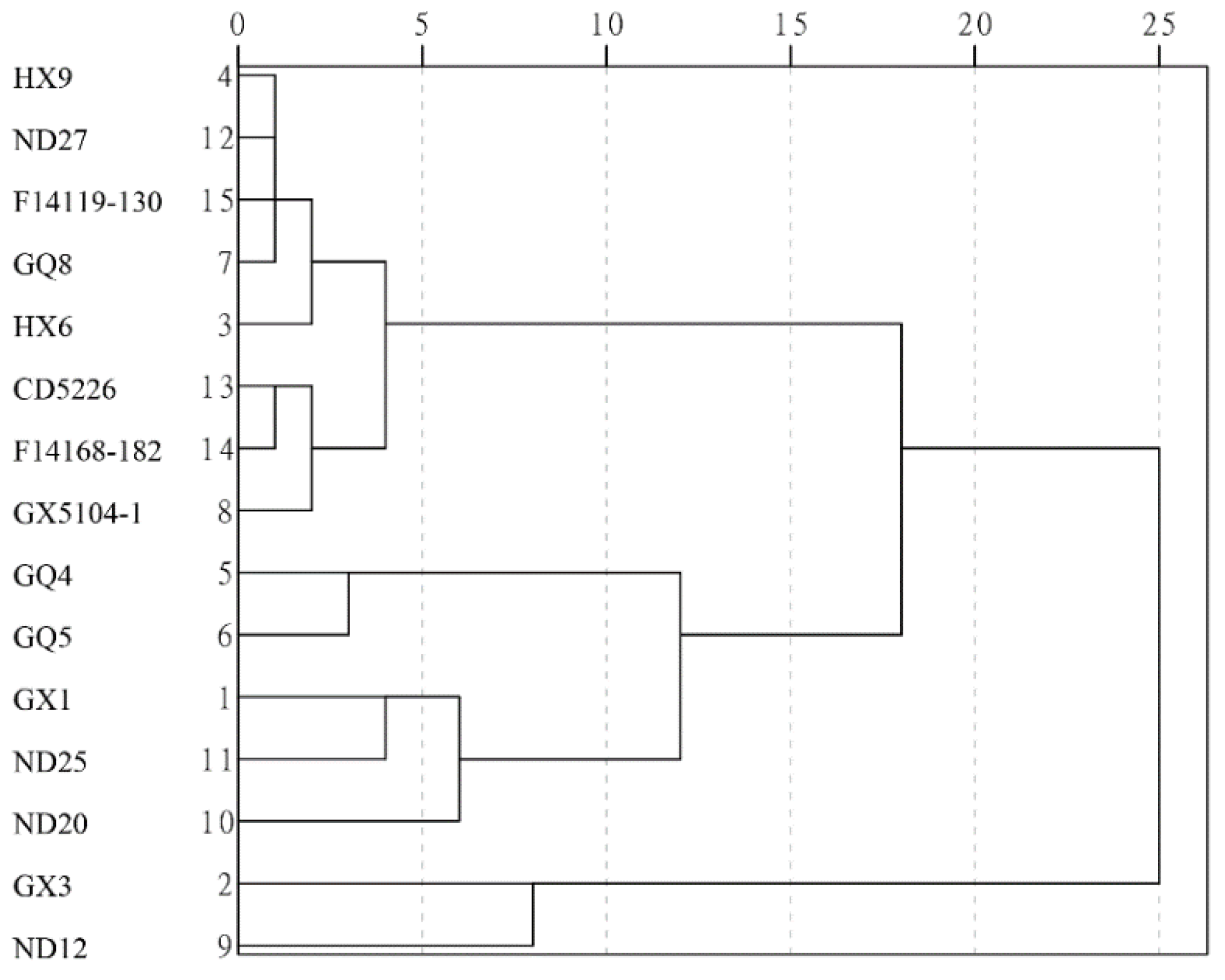
3.4. Cluster Analysis (CA) of Soybean Cultivars Based on PCA
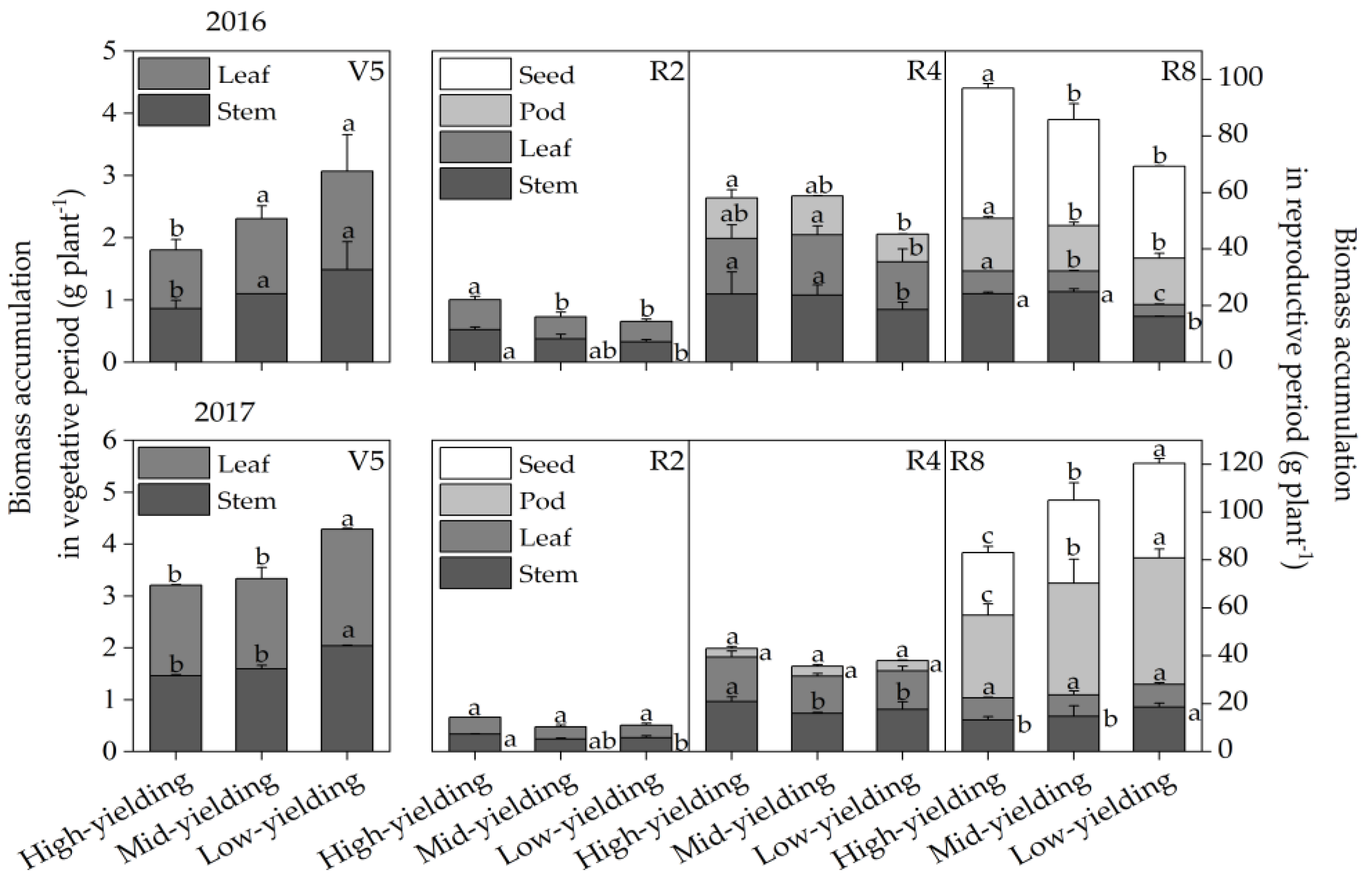
3.5. Biomass Accumulation of High-, Mid-, and Low-Yielding Cultivars
3.6. Crop Growth Parameters
4. Discussion
4.1. Reasonable Vegetative and Reproductive Growth of the High-Yielding Group Will Decrease Flower and Pod Abscission under Relay Strip Intercropping Systems
4.2. Enlightenment of Relay Intercropping System Based on Soybean High-Yielding Varieties to Improve Soybean Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, W.; Qi, Y.; Zhang, Z. A long-term forecast analysis on worldwide land uses. Environ. Monit. Assess. 2006, 119, 609–620. [Google Scholar] [CrossRef]
- Du, J.; Han, T.; Gai, J.; Yong, T.; Sun, X.; Wang, X.; Yang, F.; Liu, J.; Shu, K.; Liu, W.; et al. Maize-soybean strip intercropping: Achieved a balance between high productivity and sustainability. J. Integr. Agric. 2018, 17, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Rahman, T.; Song, C.; Yang, F.; Su, B.; Cui, L.; Bu, W.; Yang, W. Relationships among light distribution, radiation use efficiency and land equivalent ratio in maize-soybean strip intercropping. Field Crops Res. 2018, 224, 91–101. [Google Scholar] [CrossRef]
- Moore, V.M.; Schlautman, B.; Fei, S.-z.; Roberts, L.M.; Wolfe, M.; Ryan, M.R.; Wells, S.; Lorenz, A.J. Plant Breeding for Intercropping in Temperate Field Crop Systems: A Review. Front. Plant Sci. 2022, 13, 843065. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, F.; Gong, W.; Ahmed, S.; Fan, Y.; Wu, X.; Yong, T.; Liu, W.; Shu, K.; Liu, J.; et al. Shade adaptive response and yield analysis of different soybean genotypes in relay intercropping systems. J. Integr. Agric. 2017, 16, 1331–1340. [Google Scholar] [CrossRef]
- Nandula, V.K. Herbicide Resistance Traits in Maize and Soybean: Current Status and Future Outlook. Plants 2019, 8, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubiales, D.; Fondevilla, S.; Chen, W.; Gentzbittel, L.; Higgins, T.J.V.; Castillejo, M.A.; Singh, K.B.; Rispail, N. Achievements and Challenges in Legume Breeding for Pest and Disease Resistance. Crit. Rev. Plant Sci. 2015, 34, 195–236. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Francis, C.; Wu, P.; Chen, X.; Zhao, X. Maize-Soybean Intercropping Interactions Above and Below Ground. Crop Sci. 2014, 54, 914–922. [Google Scholar] [CrossRef]
- Liu, X.; Rahman, T.; Yang, F.; Song, C.; Yong, T.; Liu, J.; Zhang, C.; Yang, W. PAR Interception and Utilization in Different Maize and Soybean Intercropping Patterns. PLoS ONE 2017, 12, e0169218. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Duan, A.; Qiu, X.; Sun, J.; Zhang, J.; Liu, H.; Wang, H. Distribution and Use Efficiency of Photosynthetically Active Radiation in Strip Intercropping of Maize and Soybean. Agron. J. 2010, 102, 1149–1157. [Google Scholar] [CrossRef]
- Bing, L.; Ning, Q.D. Effects of shading on spatial distribution of flower and flower abscission in field-grown three soybeans in Northern China. Emir. J. Food Agric. 2015, 27, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Doku, E.V. Flower and pod abscission in soybean Glycine max (L.) Merril. Ghana J. Agric. Sci. 1976, 9, 119–124. [Google Scholar]
- Jia, H.; Jiang, B.; Wu, C.; Lu, W.; Hou, W.; Sun, S.; Yan, H.; Han, T. Maturity group classification and maturity locus genotyping of early-maturing soybean varieties from high-latitude cold regions. PLoS ONE 2014, 9, e94139. [Google Scholar] [CrossRef]
- Gai, J.Y.; Wang, Y.S.; Zhang, M.C.; Wang, J.A.; Chang, R.Z. Studies on the Classification of Maturity Groups of Soybeans in China. Acta Agron. Sin. 2001, 27, 286–292. [Google Scholar]
- Oberholster, S.D.; Peterson, C.M.; Dute, R.R. Pedicel abscission of soybean: Cytological and ultrastructural changes induced by auxin and ethephon. Botany 1991, 69, 2177–2186. [Google Scholar] [CrossRef]
- Jiang, H.; Egli, D.B. Shade Induced Changes in Flower and Pod Number and Flower and Fruit Abscission in Soybean. Agron. J. 1993, 85, 221–225. [Google Scholar] [CrossRef]
- Ascough, G.D.; Nogemane, N.; Mtshali, N.P.; van Staden, J.; Bornman, C.H. Flower abscission: Environmental control, internal regulation and physiological responses of plants. S. Afr. J. Bot. 2005, 71, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Burn, W.; Heindl, J.; Betts, K. The physiology of reproductive abscission in soybeans. In Proceedings of the World Soybean Research Conference III: Proceedings; CRC Press: Boca Raton, FL, USA, 2022; pp. 866–874. [Google Scholar]
- Quijano, A.; Morandi, E.N. Post-flowering leaflet removals increase pod initiation in soybean canopies. Field Crops Res. 2011, 120, 151–160. [Google Scholar] [CrossRef]
- Nagasuga, K. Soybean Seed Production and Canopy Photosynthesis in Cultivation; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Egli, D.B. Soybean Reproductive Sink Size and Short-term Reductions in Photosynthesis during Flowering and Pod Set. Crop Sci. 2010, 50, 1971–1977. [Google Scholar] [CrossRef]
- Monzon, J.P.; Cafaro La Menza, N.; Cerrudo, A.; Canepa, M.; Rattalino Edreira, J.I.; Specht, J.; Andrade, F.H.; Grassini, P. Critical period for seed number determination in soybean as determined by crop growth rate, duration, and dry matter accumulation. Field Crops Res. 2021, 261, 108016. [Google Scholar] [CrossRef]
- Kokubun, M. Physiological Mechanisms Regulating Flower Abortion in Soybean. IntechOpen, 2011. Available online: https://www.intechopen.com/chapters/15727 (accessed on 29 August 2022).
- Kakiuchi, J.; Kobata, T. Shading and Thinning Effects on Seed and Shoot Dry Matter Increase in Determinate Soybean during the Seed-Filling Period. Agron. J. 2004, 96, 398–405. [Google Scholar] [CrossRef]
- Sudhakar, P.; Latha, P.; Reddy, P.V. Phenotyping Crop Plants for Physiological and Biochemical Traits; Nikki Levy: London, UK, 2016. [Google Scholar]
- Han, S.-S.; Park, H.-J.; Shin, T.; Ko, J.; Choi, W.-J.; Lee, Y.-H.; Bae, H.-S.; Ahn, S.-H.; Youn, J.-T.; Kim, H.-Y. Effects of Tillage System, Sowing Date, and Weather Course on Yield of Double-Crop Soybeans Cultivated in Drained Paddy Fields. Agronomy 2022, 12, 1901. [Google Scholar] [CrossRef]
- Singer, J.W.; Meek, D.W.; Sauer, T.J.; Prueger, J.H.; Hatfield, J.L. Variability of light interception and radiation use efficiency in maize and soybean. Field Crops Res. 2011, 121, 147–152. [Google Scholar] [CrossRef]
- Yong, T.; Chen, P.; Dong, Q.; Du, Q.; Yang, F.; Wang, X.; Liu, W.; Yang, W. Optimized nitrogen application methods to improve nitrogen use efficiency and nodule nitrogen fixation in a maize-soybean relay intercropping system. J. Integr. Agric. 2018, 17, 664–676. [Google Scholar] [CrossRef]
- Pietsch, D.; Mabit, L. Terrace soils in the Yemen Highlands: Using physical, chemical and radiometric data to assess their suitability for agriculture and their vulnerability to degradation. Geoderma 2012, 185, 48–60. [Google Scholar] [CrossRef]
- Okalebo, J.R.; Gathua, K.W.; Woomer, P.L. Laboratory methods of soil and plant analysis: A working manual second edition. Sacred Afr. Nairobi 2002, 21, 25–26. [Google Scholar]
- Olsen, S.; Sommers, L.; Page, A. Methods of soil analysis. Part 1982, 2, 403–430. [Google Scholar]
- Wang, H.; Cheng, W.; Li, T.; Zhou, J.-m.; Chen, X. Can Nonexchangeable Potassium be Differentiated from Structural Potassium in Soils. Pedosphere 2016, 26, 206–215. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Fehr, W.R.; Caviness, C.E.; Burmood, D.T.; Pennington, J.S. Stage of Development Descriptions for Soybeans, Glycine Max (L.) Merrill. Crop Sci. 1971, 11, 929–931. [Google Scholar] [CrossRef]
- Wang, H.; Sun, X.; Yue, Y. Association mapping of flower and pod abscission with SSR markers in Northeast spring sowing soybeans. Soil Crop 2014, 3, 32–40. [Google Scholar]
- Ghanbari, S.; Nooshkam, A.; Fakheri, B.A.; Mahdinezhad, N. Assessment of Yield and Yield Component of Soybean Genotypes (Glycine Max L.) in North of Khuzestan. J. Crop Sci. Biotech. 2018, 21, 435–441. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, J.; Yang, X.; Ma, R.; Liu, C.; Liu, Q. RNA-seq Analysis Reveals Ethylene-Mediated Reproductive Organ Development and Abscission in Soybean (Glycine max L. Merr.). Plant Mol. Biol. Report. 2012, 31, 607–619. [Google Scholar] [CrossRef]
- Heitholt, J.J.; Egli, D.B.; Leggett, J.E.; Mackown, C.T. Role of Assimilate and Carbon-14 Photosynthate Partitioning in Soybean Reproductive Abortion. Crop Sci. 1986, 26, 999. [Google Scholar] [CrossRef]
- Pigeaire, A.; Seymour, M.; Delane, R.; Atkins, C. Partitioning of dry matter into primary branches and pod initiation on the main inflorescence of Lupinus angustifolius. Aust. J. Agric. Res.—AUST J AGR RES 1992, 43, 685–696. [Google Scholar] [CrossRef]
- Liang, F.; Ren, H.; Chen, C. A Study on Flower-Pod shedding And its Relation With Development of ovary And ovule In Summer Soybean (in Chinese). J. Henan Vocat. -Tech. Teach. Coll. 1991, 19, 1–6. [Google Scholar]
- Chamberlin, M.A.; Horner, H.T.; Palmer, R.G. Nutrition of ovule, embryo sac, and young embryo in soybean: An anatomical and autoradiographic study. Can. J. Bot. 1993, 71, 1153–1168. [Google Scholar] [CrossRef]
- Liu, B.; Liu, X.B.; Wang, C.; Li, Y.S.; Jin, J.; Herbert, S.J. Soybean yield and yield component distribution across the main axis inresponse to light enrichment and shading under different densities. Plant Soil Environ. 2010, 56, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Bing, L.; Yansheng, L.; Xiaobing, L.; Cheng, W.; Jian, J.; Herbert, S.J. Lower total soluble sugars in vegetative parts of soybean plants are responsible for reduced pod number under shading conditions. Aust. J. Crop Sci. 2011, 5, 1852–1857. [Google Scholar]
- Yang, F.; Liao, D.; Wu, X.; Gao, R.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; Liu, J.; et al. Effect of aboveground and belowground interactions on the intercrop yields in maize-soybean relay intercropping systems. Field Crops Res. 2017, 203, 16–23. [Google Scholar] [CrossRef]
- Greentracewicz, E.; Page, E.R.; Swanton, C.J. Shade Avoidance in Soybean Reduces Branching and Increases Plant-to-Plant Variability in Biomass and Yield Per Plant. Weed Sci. 2011, 59, 43–49. [Google Scholar] [CrossRef]
- Gommers, C.M.; Visser, E.J.; St Onge, K.R.; Voesenek, L.A.; Pierik, R.J.T.i.P.S. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Gelderen, K.V.; Kang, C.; Paalman, R.; Keuskamp, D.H.; Hayes, S.; Pierik, R. Far-red Light Detection in the Shoot Regulates Lateral Root Development through the HY5 Transcription Factor. Plant Cell 2018, 30, 101–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Years | Groups | Leaf Area (m−2) | Biomass (g plant−1) | Seed Weight (g plant−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| V5 | R2 | R4 | V5 | R2 | R4 | R8 | R8 | ||
| 2016 | High-yielding | 0.139 b | 0.667 a | 1.456 a | 1.80 b | 22.10 a | 58.03 a | 96.82 a | 45.91 a |
| Mid-yielding | 0.141 ab | 0.44 a | 1.282 ab | 2.31 ab | 16.05 b | 58.79 a | 85.81 a | 37.50 ab | |
| Low-yielding | 0.168 a | 0.500 a | 1.077 b | 3.07 a | 14.46 b | 45.32 a | 69.21 b | 32.39 b | |
| 2017 | High-yielding | 0.147 c | 0.299 a | 0.916 a | 3.21 b | 14.25 a | 43.05 a | 83.05 b | 26.07 b |
| Mid-yielding | 0.161 b | 0.329 a | 0.887 a | 3.34 b | 10.25 b | 35.63 b | 105.0 a | 34.75 a | |
| Low-yielding | 0.194 a | 0.353 a | 0.979 a | 4.29 a | 11.00 ab | 37.97 b | 120.4 a | 39.49 a | |
| ANOVA | |||||||||
| Groups (G) | * | ns | ns | * | * | ns | ns | ns | |
| Years (Y) | ns | * | ns | ** | ** | * | ** | ns | |
| G×Y | ns | ns | ns | ns | ns | ns | ns | ** | |
| Years | Groups | NAR (g m−2 day−1) | RGR (g g−1 day−1) | CGR (g m−2 day−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| V5-R2 | R2-R4 | V5-R2 | R2-R4 | R4-R8 | V5-R2 | R2-R4 | R4-R8 | ||
| 2016 | High-yielding | 1.232 a | 1.750 a | 0.0499 a | 0.0465 a | 0.0113 a | 0.812 a | 3.593 a | 1.552 a |
| Mid-yielding | 1.041 ab | 2.798 a | 0.0381 ab | 0.0662 a | 0.0077 a | 0.550 b | 4.273 a | 1.081 a | |
| Low-yielding | 0.838 b | 2.215 a | 0.0331 b | 0.0575 a | 0.0094 a | 0.45 b | 3.086 a | 0.956 a | |
| 2017 | High-yielding | 2.134 a | 3.988 a | 0.0340 a | 0.0552 a | 0.0181 a | 0.442 a | 2.880 a | 2.300 a |
| Mid-yielding | 0.867 b | 3.330 a | 0.0231 b | 0.0615 a | 0.0207 a | 0.277 b | 2.538 a | 2.776 a | |
| Low-yielding | 0.707 b | 2.860 a | 0.0216 b | 0.0621 a | 0.0229 a | 0.269 b | 2.697 a | 3.297 a | |
| Groups (G) | * | ns | * | ns | ns | * | ns | ns | |
| Years (Y) | ns | ns | ns | * | ns | ns | ns | ns | |
| G×Y | ns | ns | ns | ns | ns | ns | ns | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, Q.; Chen, P.; Zheng, B.; Hu, Y.; Yang, W.; Yong, T. Screening Soybean for Adaptation to Relay Intercropping Systems: Associations between Reproductive Organ Abscission and Yield. Agronomy 2022, 12, 2379. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102379
Du Q, Chen P, Zheng B, Hu Y, Yang W, Yong T. Screening Soybean for Adaptation to Relay Intercropping Systems: Associations between Reproductive Organ Abscission and Yield. Agronomy. 2022; 12(10):2379. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102379
Chicago/Turabian StyleDu, Qing, Ping Chen, Benchuan Zheng, Yongchun Hu, Wenyu Yang, and Taiwen Yong. 2022. "Screening Soybean for Adaptation to Relay Intercropping Systems: Associations between Reproductive Organ Abscission and Yield" Agronomy 12, no. 10: 2379. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102379