
Efficient Micropropagation of Genetically Stable Panax ginseng Meyer by Somatic Embryogenesis
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
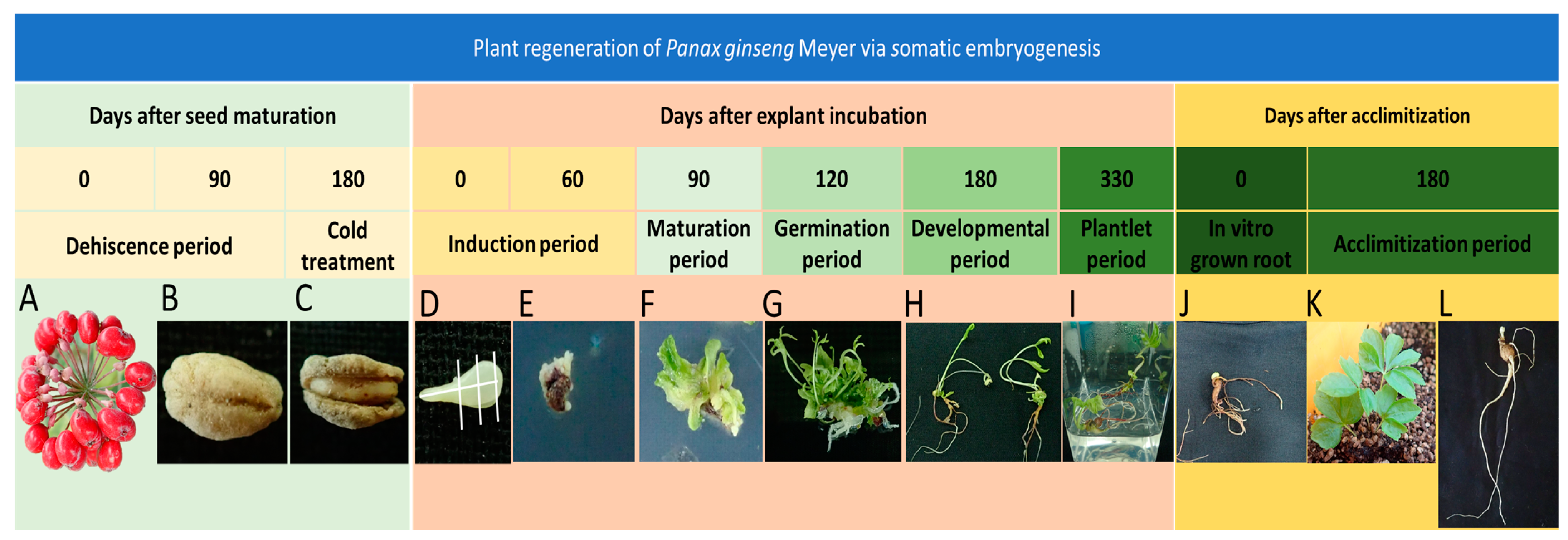
2.1. Plant Materials and Seed Sterilization
2.2. In Vitro Regeneration Protocol and Culture Conditions
2.3. Somatic Embryogenesis
2.4. Maturation and Germination of Somatic Embryos
2.5. Plant Regeneration and Plantlet Conversion
2.6. Acclimatization
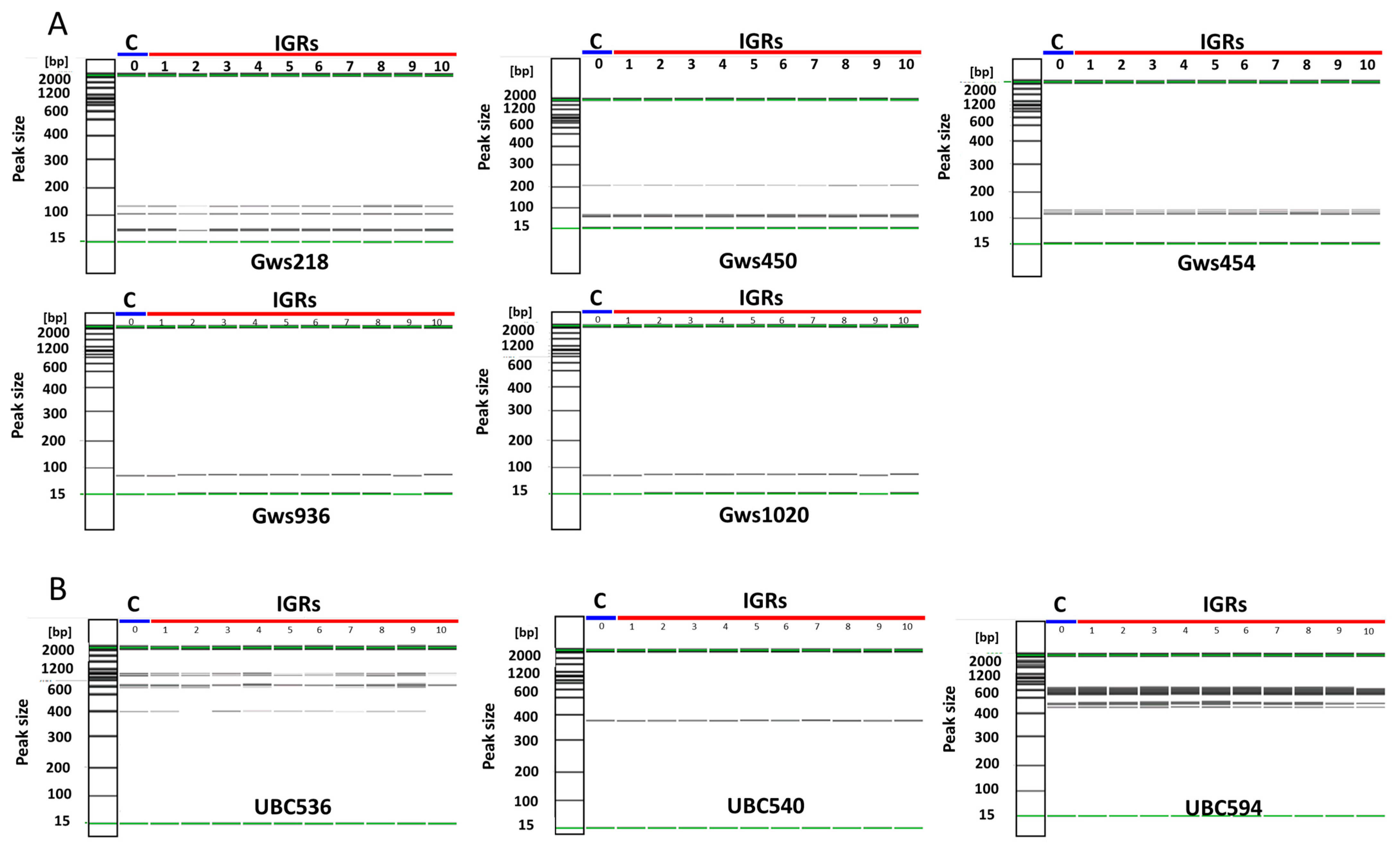
2.7. Determination of Genetic Homogeneity
2.8. Ginsenoside Analysis
2.9. Statistical Analysis
3. Results
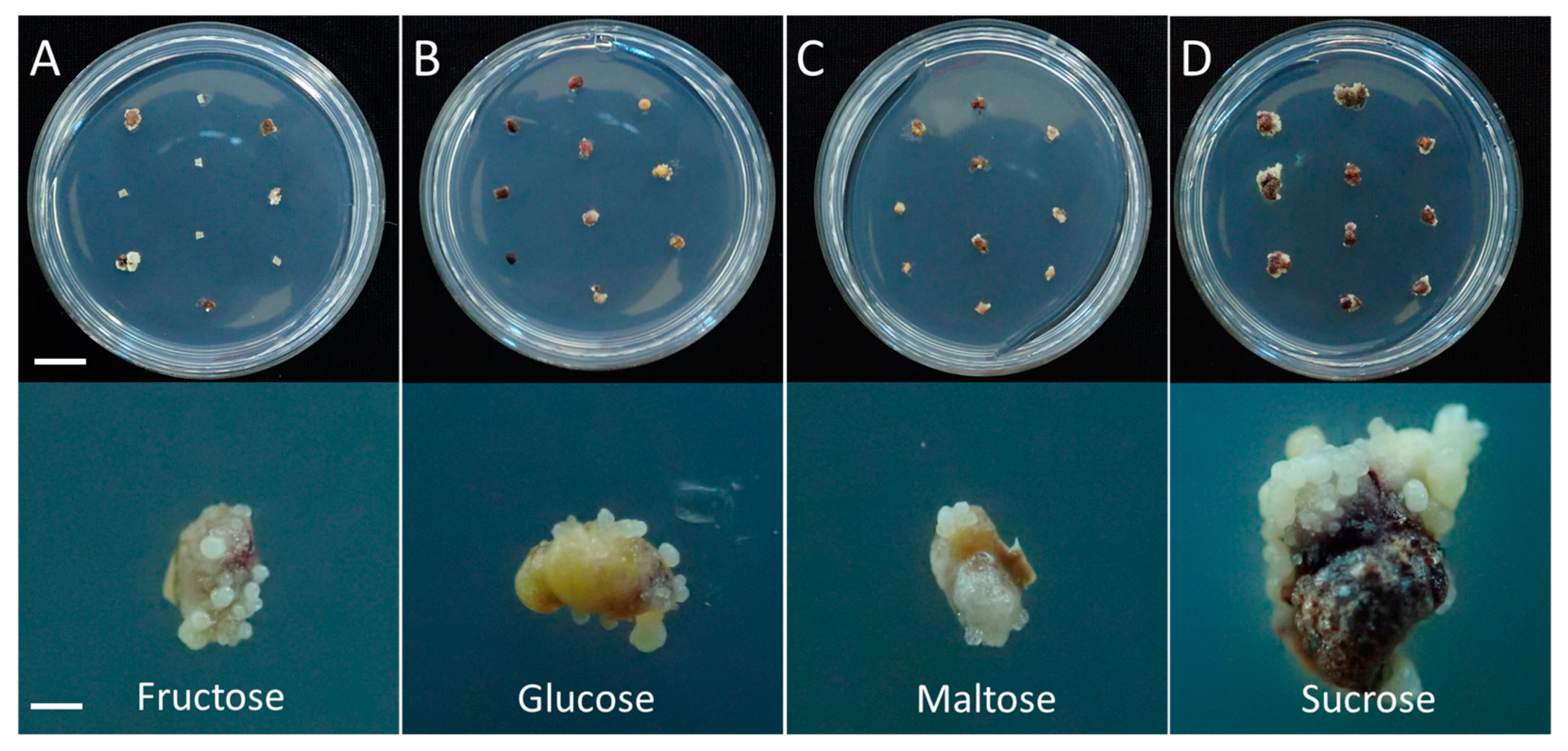
3.1. Effect of Carbon Source on Somatic Embryogenesis
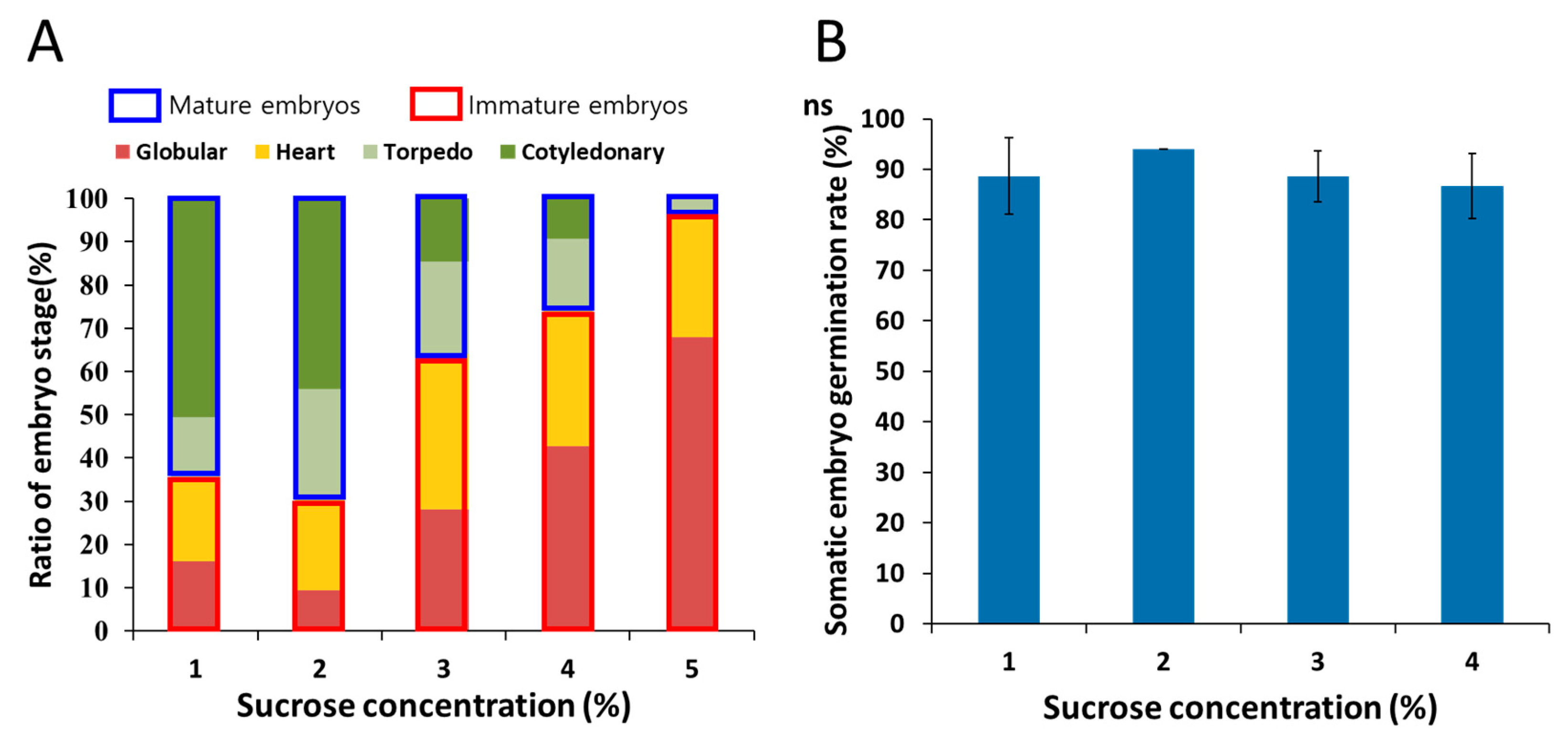
3.2. Effect of Sucrose Concentration on Somatic Embryo Maturation and Germination
3.3. Effect of Sucrose Concentration on Plant Regeneration
3.4. Acclimatization and Determination of Genetic Homogeneity
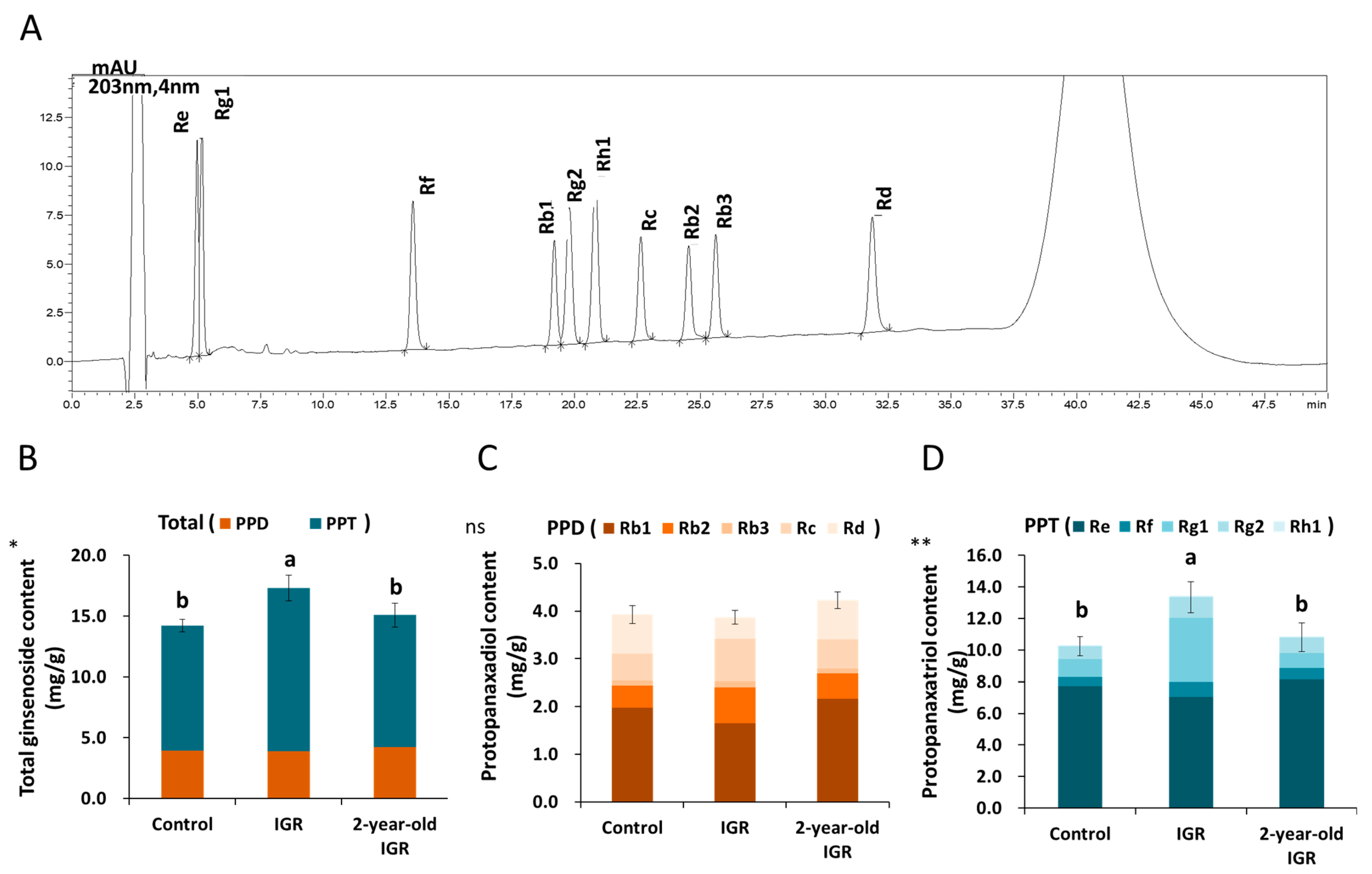
3.5. Ginsenoside Content Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nguyen, V.B.; Giang, V.N.L.; Waminal, N.E.; Park, H.S.; Kim, N.H.; Jang, W.; Lee, J.; Yang, T.J. Comprehensive comparative analysis of chloroplast genomes from seven Panax species and development of an authentication system based on species-unique single nucleotide polymorphism markers. J. Ginseng Res. 2020, 44, 135–144. [Google Scholar] [CrossRef]
- Glagoleva, E.S.; Konstantinova, S.E.; Kochkin, D.V.; Ossipov, V.; Titova, M.V.; Popova, E.V.; Nosov, A.M.; Paek, K.Y. Predominance of oleanane-type ginsenoside R0 and malonyl esters of protopanaxadiol-type ginsenosides in the 20-year-old suspension cell culture of Panax japonicus C.A. Meyer. Indus. Crops Prod. 2022, 177, 114417. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, D.; Lee, H.L.; Lim, C.E.; Jung, K.; Kang, K.S. Beneficial effects of Panax ginseng for the treatment and prevention of neurodegenerative diseases: Past findings and future directions. J. Ginseng Res. 2018, 42, 239–247. [Google Scholar] [CrossRef]
- Kim, J.H. Pharmacological and medical applications of Panax ginseng and ginsenosides: A review for use in cardiovascular diseases. J. Ginseng Res. 2018, 42, 264–269. [Google Scholar] [CrossRef]
- Shao, J.W.; Jiang, J.L.; Zou, J.J.; Yang, M.Y.; Chen, F.M.; Zhang, Y.J.; Lee, J. Therapeutic potential of ginsenosides on diabetes: From hypoglycemic mechanism to clinical trials. J. Funct. Foods 2020, 64, 103630. [Google Scholar] [CrossRef]
- Hong, H.; Baatar, D.; Hwang, S.G. Anticancer activities of ginsenosides, the main active components of ginseng. J. Ginseng Res. 2000, 24, 8858006. [Google Scholar] [CrossRef]
- Lee, J.W.; Kwon, N.; Kim, J.U.; Bang, K.H.; Jung, S.M.; Lee, S.W.; Kim, D.H.; Kim, Y.C.; Jo, I.H.; Park, Y.D. In vitro micropropagation of commercial ginseng cultivars (Panax ginseng Meyer) via somatic embryogenesis compared to traditional seed production. Horiculturae 2023, 9, 435. [Google Scholar] [CrossRef]
- Lee, J.W.; Jo, I.H.; Kim, J.U.; Hong, C.E.; Kim, Y.C.; Kim, D.H.; Park, Y.D. Improvement of seed dehiscence and germination in ginseng by stratification, gibberellin, and/or kinetin treatments. Hortic. Environ. Biotechnol. 2018, 59, 293–301. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, O.R.; Kim, K.T.; Yang, D.C. High frequency of plant regeneration through cyclic secondary somatic embryogenesis in Panax ginseng. J. Ginseng Res. 2012, 36, 442–448. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kim, D.H.; Bang, K.H.; Kim, J.U.; Hyun, D.Y.; Lee, S.W.; Kang, S.W.; Cha, S.W.; Kim, K.H.; Choi, J.K.; et al. A high yielding and salt resistance ginseng variety “Cheonryang”. Korean J. Breed. Sci. 2013, 45, 434–439. [Google Scholar] [CrossRef]
- Lee, J.W.; Do, G.R.; Jo, I.H.; Hong, C.E.; Bang, K.H.; Kim, J.U.; Park, Y.D. Zygotic embryo culture is an efficient way to optimize in vitro growth in Panax ginseng. Indus. Crops Prod. 2021, 167, 1134971. [Google Scholar] [CrossRef]
- Manchanda, P.; Gosal, S.S. Effect of activated charcoal, carbon sources and gelling agents on direct somatic embryogenesis and regeneration in sugarcane via leaf roll segments. Sugar Tech 2012, 14, 168–173. [Google Scholar] [CrossRef]
- Shohael, A.M.; Chakrabarty, D.; Ali, M.B.; Yu, K.W.; Hahn, E.J.; Lee, H.L.; Paek, K.Y. Enhancement of eleutherosides production in embryogenic cultures of Eleutherococcus sessiliflorus in response to sucrose-induced osmotic stress. Process Biochem. 2006, 41, 512–518. [Google Scholar] [CrossRef]
- Nhut, D.T.; Huy, N.P.; Binh, N.V.; Nam, N.B.; Thuy, L.N.; Ha, D.T.; Chien, H.X.; Huong, T.T.; Cuong, L.K.; Hien, V.T. Shoot regeneration and micropropagation of Panax vietnamensis Ha et Grushv. from ex vitro leaf-derived callus. Afr. J. Biotechnol. 2011, 10, 19499–19504. [Google Scholar] [CrossRef]
- Nhut, D.T.; Vinh, B.V.T.; Hien, T.T.; Huy, N.P.; Nam, N.B.; Chien, H.X. Effects of spermidine, proline and carbohydrate sources on somatic embryogenesis from main root transverse thin cell layers of Vietnamese ginseng (Panax vietnamensis Ha et. Grushv.). Afr. J. Biotechnol. 2012, 11, 1084–1091. [Google Scholar] [CrossRef]
- Wojtania, A.; Skrzypek, E.; Marasek-Ciolakowska, A. Soluble sugar, starch and phenolic status during rooting of easy and difficult-to-root magnolia cultivars. Plant Cell Tiss. Organ Cult. 2019, 136, 499–510. [Google Scholar] [CrossRef]
- Heydari, H.R.; Chamani, E. High-frequency plant regeneration and genetic fidelity assessment of regenerants by molecular and biochemical markers in Woodland Sage (Salvia nemorosa L.). Indus. Crops Prod. 2022, 187, 115404. [Google Scholar] [CrossRef]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef]
- Gaj, M.D. Direct somatic embryogenesis as a rapid and efficient system for in vitro regeneration of Arabidopsis thaliana. Plant Cell Tiss. Organ Cult. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Konar, S.; Karmakar, J.; Ray, A.; Adhikari, S.; Bandyopadhyay, T.K. Regeneration of plantlets through somatic embryogenesis from root derived calli of Hibiscus sabdariffa L. (Roselle) and assessment of genetic stability by flow cytometry and ISSR analysis. PLoS ONE 2018, 13, e0202324. [Google Scholar] [CrossRef]
- Mamgain, J.; Mujib, A.; Ejaz, B.; Gulzar, B.; Malik, M.Q.; Syeed, R. Flow cytometry and start codon targeted (SCoT) genetic fidelity assessment of regenerated plantlets in Tylophora indica (Burm.f.) Merrill. Plant Cell Tiss. Organ Cult. 2022, 150, 129–140. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Sun, H.J.; Song, I.J.; Bae, T.W.; Kang, H.G.; Ko, S.M.; Kwon, Y.I.; Kim, I.W.; Lee, J.C.; Park, S.Y.; et al. Plant regeneration of Korean wild ginseng (Panax ginseng Meyer) mutant lines induced by γ-irradiation (60Co) of adventitious roots. J. Ginseng Res. 2014, 38, 220–225. [Google Scholar] [CrossRef]
- Kim, J.Y.; Adhikari, P.B.; Ahn, C.H.; Kim, D.H.; Chang Kim, Y.C.; Han, J.Y.; Kondeti, S.; Choi, Y.E. High frequency somatic embryogenesis and plant regeneration of interspecific ginseng hybrid between Panax ginseng and Panax quinquefolius. J. Ginseng Res. 2019, 43, 38–48. [Google Scholar] [CrossRef]
- Choi, Y.E.; Soh, W.Y. Enhanced somatic single embryo formation by plasmolyzing pretreatment from cultured ginseng cotyledons. Plant Sci. 1997, 130, 197–2064. [Google Scholar] [CrossRef]
- Choi, Y.E.; Yang, D.C.; Yoon, E.S.; Choi, K.T. High-efficiency plant production via direct somatic single embryogenesis from preplasmolysed cotyledons of Panax ginseng and possible dormancy of somatic embryos. Plant Cell Rep. 1999, 18, 493–499. [Google Scholar] [CrossRef]
- Kim, O.T.; Kim, T.S.; In, D.S.; Bang, K.H.; Kim, Y.C.; Choi, Y.E.; Cha, S.W.; Seong, N.S. Optimization of direct somatic embryogenesis from mature zygotic embryos of Panax ginseng C. A. Meyer. J. Plant Biol. 2006, 49, 348–352. [Google Scholar] [CrossRef]
- Lee, J.W.; Jo, I.H.; Kim, J.U.; Hong, C.E.; Bang, K.H.; Park, Y.D. Determination of mutagenic sensitivity to gamma rays in ginseng (Panax ginseng) dehiscent seeds, roots, and somatic embryos. Hortic. Environ. Biotechnol. 2019, 60, 721–731. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, J.U.; Bang, K.H.; Kwon, N.; Kim, Y.C.; Jo, I.H.; Park, Y.D. Efficient somatic embryogenesis, regeneration and acclimatization of Panax ginseng Meyer: True-to-type conformity of plantlets as confirmed by ISSR analysis. Plants 2023, 12, 1270. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A.C. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Raomai, S.; Kumaria, S.; Tandon, P. Plant regeneration through direct somatic embryogenesis from immature zygotic embryos of the medicinal plant, Paris polyphylla Sm. Plant Cell Tiss. Organ Cult. 2014, 118, 445–455. [Google Scholar] [CrossRef]
- Jang, W.; Jang, Y.; Kim, N.H.; Waminal, N.E.; Kim, Y.C.; Lee, J.W.; Yang, T.J. Genetic diversity among cultivated and wild Panax ginseng populations revealed by high-resolution microsatellite markers. J. Ginseng Res. 2020, 44, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Brown, D.C.W. High efficiency plant production of North American ginseng via somatic embryogenesis from cotyledon explants. Plant Cell Rep. 2006, 25, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Traore, A.; Guiltinan, M. Effects of carbon source and explant type on somatic embryogenesis of four cacao genotypes. Hortic. Sci. 2006, 41, 753–758. [Google Scholar] [CrossRef]
- Faisal, M.; Abdel-Salam, E.M.; Alatar, A.A.; Qahtan, A.A. Induction of somatic embryogenesis in Brassica juncea L. and analysis of regenerants using ISSR-PCR and flow cytometer. Saudi J. Biol. Sci. 2021, 28, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.K.; Kim, Y.W.; Hong, Y.P.; Park, S.Y. Improvement of somatic embryogenesis and plantlet conversion in Oplopanax elatus, an endangered medicinal woody plant. SpringerPlus 2013, 2, 428. [Google Scholar] [CrossRef]
- Blanc, G.; Lardet, L.; Martin, A.; Jacob, J.L.; Carron, M.P. Differential carbohydrate metabolism conducts morphogenesis in embryogenic callus of Hevea brasiliensis (Müll. Arg.). J. Exp. Bot. 2002, 53, 1453–1462. [Google Scholar] [CrossRef]
- Baskaran, P.; Kumari, A.; Van Staden, J. Embryogenesis and synthetic seed production in Mondia whitei. Plant Cell Tiss. Organ Cult. 2015, 121, 205–214. [Google Scholar] [CrossRef]
- Karami, O.; Deljou, A.; Esna-Ashari, M.; Ostad-Ahmadi, P. Effect of sucrose concentrations on somatic embryogenesis in carnation (Dianthus caryophyllus L.). Sci. Hortic. 2006, 110, 340–344. [Google Scholar] [CrossRef]
- Du, Y.; Cheng, F.; Zhong, Y. Induction of direct somatic embryogenesis and shoot organogenesis and histological study in tree peony (Paeonia sect. Moutan). Plant Cell Tiss. Organ Cult. 2020, 141, 557–570. [Google Scholar] [CrossRef]
- Woo, H.A.; Ku, S.S.; Jie, E.Y.; Kim, H.; Kim, H.S.; Cho, H.S.; Jeong, W.J.; Park, S.U.; Min, S.R.; Kim, S.W. Efficient plant regeneration from embryogenic cell suspension cultures of Euonymus alatus. Sci. Rep. 2021, 11, 15120. [Google Scholar] [CrossRef] [PubMed]
- Tereso, S.; Zoglauer, K.; Milhinhos, A.; Miguel, C.; Oliveira, M.M. Zygotic and somatic embryo morphogenesis in Pinus pinaster: Comparative histological and histochemical study. Tree Physiol. 2007, 27, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Etienne, H.; Bertrand, B.; Georget, F.; Lartaud, M.; Montes, F.; Dechamp, E.; Verdeil, J.L.; Barry-Etienne, D. Development of coffee somatic and zygotic embryos to plants differs in the morphological, histochemical and hydration aspects. Tree Physiol. 2013, 33, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.R.; Dutta Gupta, S. High-frequency plant regeneration through cyclic secondary somatic embryogenesis in black pepper (Piper nigrum L.). Plant Cell Rep. 2006, 24, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, D.H.; Kim, Y.C.; Kim, K.H.; Han, J.Y.; Choi, Y.E. In vitro grown thickened taproots, a new type of soil transplanting source in Panax ginseng. J. Ginseng Res. 2016, 40, 409–414. [Google Scholar] [CrossRef]
- Mathur, A.; Mathur, A.K.; Gangwar, A.; Yadav, S.; Verma, P.; Sangwan, R.S. Anthocyanin production in a callus line of Panax sikkimensis Ban. In Vitro Cell Dev. Biol. Plant 2010, 46, 13–21. [Google Scholar] [CrossRef]
- Yaseen, M.; Ahmad, T.; Sablok, G.; Standardi, A.; Hafiz, I.A. Review: Role of carbon sources for in vitro plant growth and development. Mol. Biol. Rep. 2013, 40, 2837–2849. [Google Scholar] [CrossRef]
- Choi, Y.E.; Yang, D.C.; Yoon, E.S.; Choi, K.T. Plant regeneration via adventitious bud formation from cotyledon explants of Panax ginseng C. A. Meyer. Plant Cell Rep. 1998, 17, 731–736. [Google Scholar] [CrossRef]
- Takahashi, W.; Sugawara, F.; Yamamoto, N.; Bando, E.; Matsushita, J.; Tanaka, O. Plant regeneration in Actinidia polygama Miq. by leaf, stem, and petiole culture with zeatin, and from stem-derived calli on low-sucrose medium. J. For. Res. 2004, 9, 85–88. [Google Scholar] [CrossRef]
- Martins, J.P.R.; Verdoodt, V.; Pasqual, M.; De Proft, M. Impacts of photoautotrophic and photomixotrophic conditions on in vitro propagated Billbergia zebrina (Bromeliaceae). Plant Cell Tiss. Organ Cult. 2015, 123, 121–132. [Google Scholar] [CrossRef]
- Zahid, N.A.; Jaafar, H.Z.E.; Hakiman, M. Alterations in microrhizome induction, shoot multiplication and rooting of ginger (Zingiber officinale Roscoe) var. Bentong with regards to sucrose and plant growth regulators application. Agronomy 2021, 11, 320. [Google Scholar] [CrossRef]
- Nhut, D.T.; Le, B.V.; Fukai, S.; Tanaka, M.; Van, T.T. Effects of activated charcoal, explant size, explant position and sucrose concentration on plant and shoot regeneration of Lilium longiflorum via young stem culture. Plant Growth Regul. 2021, 33, 59–65. [Google Scholar] [CrossRef]
- Chandra, S.; Bandopadhyay, R.; Kumar, V.; Chandra, R. Acclimatization of tissue cultured plantlets: From laboratory to land. Biotechnol. Lett. 2010, 32, 1199–1205. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gu, H.; Ye, C.; Guo, C.; Zhu, Y.; Huang, H.; Liu, Y.; He, X.; Yang, M.; Zhu, S. Planting density affects Panax notoginseng growth and ginsenoside accumulation by balancing primary and secondary metabolism. Front. Plant Sci. 2021, 12, 628294. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Song, M.; Kim, S.H.; Jang, D.S.; Kim, J.B.; Ha, B.K.; Kim, S.H.; Lee, K.J.; Kang, S.Y.; Jeong, I.Y. The improvement of ginsenoside accumulation on Panax ginseng as a result of γ-irradiation. J. Ginseng Res. 2013, 37, 332–340. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, W.H.; Sun, X.Y.; Li, K.H.; Liu, K.J.; Wang, J.; Wang, Y.; Tan, X.; You, X.L. A culture system for the stable and high-efficiency proliferation of adventitious roots of Panax notoginseng and ginsenoside accumulation. Indus. Crops Prod. 2021, 157, 12882. [Google Scholar] [CrossRef]
- Kim, Y.J.; Zhang, D.; Yang, D.C. Biosynthesis and biotechnological production of ginsenosides. Biotechnol. Adv. 2015, 33, 717–735. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carbon Source (3%) | Somatic Embryo Induction Rate (%) *** | Mean Number of Somatic Embryos/Explant ** | EFC Index |
|---|---|---|---|
| Fructose | 42.3 ± 4.0 c | 9.6 ± 2.6 b | 4.1 |
| Glucose | 72.7 ± 10.6 b | 9.6 ± 3.4 b | 7.0 |
| Maltose | 66.3 ± 7.7 b | 9.6 ± 2.8 b | 6.4 |
| Sucrose | 93.3 ± 3.5 a | 16.2 ± 3.7 a | 15.1 |
| Sucrose Concentration (%) | Somatic Embryo Induction Rate (%) *** | Mean Number of Somatic Embryos/Explant ** | EFC Index |
|---|---|---|---|
| 1 | 72.4 ± 8.3 b | 8.4 ± 4.1 b | 4.0 |
| 3 | 91.3 ± 4.5 a | 21.5 ± 6.3 a | 18.5 |
| 5 | 97.7 ± 2.8 a | 23.8 ± 7.8 a | 21.3 |
| 7 | 97.3 ± 4.3 a | 12.8 ± 7.5 b | 10.0 |
| 9 | 65.1 ± 11.7 b | 10.5 ± 2.3 b | 5.1 |
| Sucrose Concentration (%) | Morphological Characteristics |
|---|---|
| 1 | Soft explant with light-red pigment accumulation, easily separated shoots, long and weak shoots, occasional shoot necrosis |
| 2 | Soft explants with light-red pigment accumulation, easily separated shoots, vigorous shoot growth |
| 3 | Hard explant with red pigment accumulation, difficult to separate shoots, short and poor shoot structure, secondary somatic embryogenesis |
| 4 | Very hard explant with dark-red pigment accumulation, difficult to separate shoots, short and poor shoot structure, secondary somatic embryogenesis |
| Sucrose Concentration (%) | Mean Number of Regenerated Plants | ||||
|---|---|---|---|---|---|
| Total *** | W ** | S * | R ** | Plantlet ** | |
| 1 | 18.6 ± 2.3 a | 10.3 ± 1.9 a | 5.5 ± 1.6 a | 2.8 ± 1.1 bc | 13.1 ± 2.2 a |
| 2 | 15.7 ± 2.8 ab | 8.6 ± 1.1 ab | 2.3 ± 1.1 b | 4.9 ± 1.4 a | 13.5 ± 2.1 a |
| 3 | 13.4 ± 0.7 b | 6.8 ± 0.8 bc | 2.5 ± 1.4 b | 4.1 ± 0.1 ab | 10.9 ± 0.8 a |
| 4 | 7.0 ± 1.4 c | 4.5 ± 0.6 c | 1.3 ± 0.4 b | 1.2 ± 0.9 c | 5.7 ± 1.2 b |
| Sucrose Concentration (%) | Aerial Part | Underground Part | ||||
|---|---|---|---|---|---|---|
| Shoot Length *** | Shoot Diameter *** | Shoot Weight ** | Root Length ns | Root Diameter *** | Root Weight ** | |
| 1 | 5.3 ± 1.3 a | 0.93 ± 0.21 a | 89.5 ± 60.0 a | 2.9 ± 0.1 | 0.7 ± 0.1 c | 26.0 ± 13.5 b |
| 2 | 2.2 ± 0.4 b | 0.55 ± 0.10 b | 47.7 ± 12.6 b | 3.3 ± 0.3 | 1.2 ± 0.3 b | 59.8 ± 25.2 a |
| 3 | 1.9 ± 0.8 b | 0.52 ± 0.16 b | 36.6 ± 17.0 b | 3.7 ± 0.5 | 1.6 ± 0.5 a | 54.7 ± 26.2 a |
| 4 | 1.8 ± 0.6 b | 0.61 ± 0.09 b | 34.6 ± 14.3 b | 3.3 ± 0.4 | 1.5 ± 0.4 ab | 54.2 ± 23.2 a |
| Control | IGR | 2-Year-Old IGR | ||
|---|---|---|---|---|
| PPD (mg/g) ns | Rb1 *** | 1.98 ± 0.11 a | 1.65 ± 0.05 b | 2.17 ± 0.12 a |
| Rb2 *** | 0.47 ± 0.02 c | 0.75 ± 0.03 a | 0.52 ± 0.01 b | |
| Rb3 *** | 0.09 ± 0.00 c | 0.12 ± 0.00 a | 0.11 ± 0.00 b | |
| Rc *** | 0.57 ± 0.03 b | 0.89 ± 0.04 a | 0.62 ± 0.00 b | |
| Rd *** | 0.82 ± 0.02 a | 0.45 ± 0.02 b | 0.82 ± 0.04 a | |
| PPT (mg/g) ** | Re ns | 7.72 ± 0.55 a | 7.04 ± 0.65 a | 8.14 ± 0.81 a |
| Rf *** | 0.59 ± 0.04 c | 0.94 ± 0.05 a | 0.70 ± 0.04 b | |
| Rg1 *** | 1.12 ± 0.08 b | 4.07 ± 0.28 a | 0.95 ± 0.02 b | |
| Rg2 *** | 0.83 ± 0.04 c | 1.29 ± 0.08 a | 1.02 ± 0.08 b | |
| Rh1 *** | 0.04 ± 0.00 c | 0.08 ± 0.00 a | 0.04 ± 0.00 b | |
| PPD/PPT ** | 0.38 ± 0.03 a | 0.29 ± 0.02 b | 0.39 ± 0.03 a | |
| Total Ginsenoside Content (mg/g) * | 14.22 ± 0.53 b | 17.30 ± 1.05 a | 15.08 ± 0.97 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-W.; Kim, J.-U.; Bang, K.-H.; Kim, D.-H.; Jo, I.-H.; Park, Y.-D. Efficient Micropropagation of Genetically Stable Panax ginseng Meyer by Somatic Embryogenesis. Agronomy 2023, 13, 1139. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041139
Lee J-W, Kim J-U, Bang K-H, Kim D-H, Jo I-H, Park Y-D. Efficient Micropropagation of Genetically Stable Panax ginseng Meyer by Somatic Embryogenesis. Agronomy. 2023; 13(4):1139. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041139
Chicago/Turabian StyleLee, Jung-Woo, Jang-Uk Kim, Kyong-Hwan Bang, Dong-Hwi Kim, Ick-Hyun Jo, and Young-Doo Park. 2023. "Efficient Micropropagation of Genetically Stable Panax ginseng Meyer by Somatic Embryogenesis" Agronomy 13, no. 4: 1139. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041139