1. Introduction
The night lily (or long yellow daylily),
H. citrina Baroni, is one of the most economically important plant species in the genus
Hemerocallis (family Asphodelaceae, previously Hemerocallidaceae or Liliaceae) [
1]. In East Asian countries, the night lily is a popular specialty vegetable crop, and its dried immature flower buds serve as a primary food source [
1,
2]. In recent years, a marked increase in the production and consumption of night lilies has occurred in China. For instance, in 2018, more than 60,370 hectares of night lilies were cultivated, yielding 561,300 tons of immature flowers, translating to an economic value of nearly CNY 4 billion [
1,
2,
3]. Research on night lilies has made significant progress, including the collection and molecular characterization of germplasm resources and the development of various genetic and genomic resources, such as genome sequences and molecular markers [
1,
2,
4,
5,
6,
7,
8]. This research lays a solid foundation for expediting the breeding of this crop through molecular mapping.
The night lily is an herbaceous perennial and it is commercially propagated asexually by dividing the crown or by making cuttings, while seed propagation is used for breeding [
3,
9,
10]. In sexual propagation, the night lily undergoes a prolonged juvenile (vegetative) growth phase, typically requiring two to three years to reach the flowering stage. After several years of growth, the night lily plants are composed of several interconnected crowns, which can be separated into individual plants. There are two primary challenges in the propagation of night lilies. In seed production, the extended growth period necessary for new plant development is particularly cumbersome in genetics and breeding research, especially when developing segregating populations for molecular-marker-related studies [
5,
11]. In commercial production, the annual rate of new crown development is limited, averaging an 8:1 ratio depending on the genotype [
10,
12]. Thus, for a newly released variety, it may take more than 10 years to produce enough plants to meet market demand [
9].
One solution for addressing these bottlenecks in plant production is plant tissue culture and micropropagation, which rely on the totipotency of plant cells for regenerating plants [
13]. During in vitro plant regeneration, an explant undergoes cell division and differentiation, during which organs and tissues form throughout its growth. This process can be realized through somatic embryogenesis or organogenesis [
14,
15,
16]. In vitro micropropagation can accelerate vegetative propagation and has been widely used for commercial production of many horticultural crops [
17,
18,
19,
20,
21]. Krikorian and Kann were the first to demonstrate the totipotency of
H. citrina plants [
22]. Since then, extensive studies have been conducted on tissue culture and micropropagation in night lilies. Factors affecting callus induction, shoot and plant regeneration in the night lily have been investigated to optimize various culture conditions, including plant genotypes, explants, basal growth media and supplemental plant growth regulators (PGRs) [
10,
23,
24,
25,
26]. For explants, micropropagation has been performed with young inflorescences, flower stems (scapes) or petals [
26,
27,
28,
29,
30], ovaries [
22], flower stems (scapes), leaves [
11], suspension culture cells [
31,
32], isolated protoplasts [
23,
33], anther filaments and immature seed embryos [
10,
34]. More recently, Matand et al. [
30] reported high-frequency in vitro regeneration of adventitious shoots in daylily stem tissue (scapes) using Murashige and Skoog (MS) medium [
35] supplemented with thidiazuron (TDZ), which enhances culture efficiency in various tissues and genotypes [
36,
37]. The authors suggested this protocol for the mass multiple shoot formation and rapid propagation of this crop to meet market demands and expedite exogenous gene transfer and breeding selection processes.
Currently, there is no established protocol for the large-scale production of night lily plants through in vitro micropropagation. Additionally, the perennial growth habit of these plants indicates the seasonal availability of explants for night lilies. Hence, this study aimed to develop an efficient and rapid protocol for tissue culture of night lilies. We evaluated various factors that affect the propagation of the night lily, including basal growth media, plant growth regulators (PGRs), explants for callus induction, shoot regeneration, subculture, rooting and transplanting. All our experiments were performed with the night lily cultivar ‘Datong Huang Hua’ (‘Datong Yellow Flower’), which is a popular variety that has been widely grown in Shanxi Province for many years and has a high yield [
1,
2]. Its genome is also being assembled in the senior author’s laboratory (unpublished data). In this work, we proposed a protocol for the rapid and large-scale production of night lily plants through micropropagation.
2. Materials and Methods
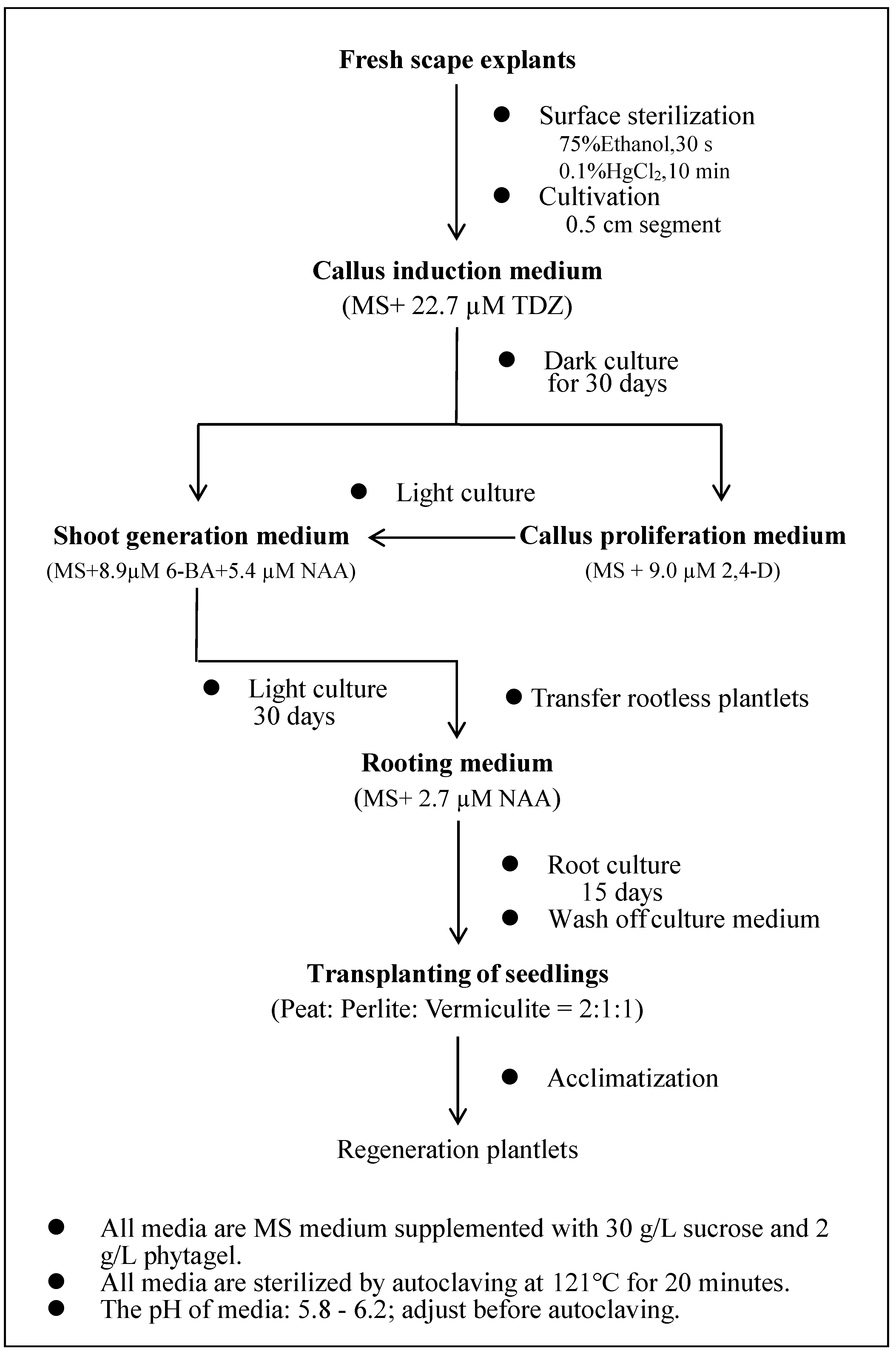
2.1. Plant Materials and Explant Culture Conditions
Young leaf or flower stem (scape) segments from the night lily variety ‘Datong Huanghua’ were used as explants in tissue culture in this study. This variety was grown in the Hemerocallis germplasm nursery at Shanxi Agricultural University (Taigu, Shanxi Province, China) for 10 years. The leaf was collected in late March when the leaf started to grow. The scape was collected in June before the flower bud started to expand.
The explants were initially rinsed with tap water and placed in sterile jars. Subsequently, the explants were rinsed once with sterile deionized water (dH
2O) on an ultraclean bench, the surface disinfected with 75% ethanol for 30 s, and rinsed twice with dH
2O, followed by sterilization in 0.1% HgCl
2 for 10 min with continuous shaking. Afterwards, the explants were rinsed five times with dH
2O. The entire procedure was performed under sterile conditions in a laminar flow hood. Detailed information (names and vendors) on all the chemicals and components of the growth media used in this study is provided in
Supplemental Files 1a and 1b.
The explants were cultured in 200 mL glass jars containing approximately 25 mL of culture medium containing MS basal salts, vitamins, sucrose (30 g/L) and phytagel (2 g/L), along with different concentrations of PGRs depending on the specific treatment (see below). The pH of the MS medium was adjusted to 5.8~6.2 with 1 M NaOH, followed by autoclaving at 121 °C for 20 min. The tissue culture room condition was 25 ± 2 °C incubation temperature. There were two types of photoperiod conditions: dark culture conditions (24 h of total darkness) and light culture conditions (16 h photoperiod (6:00~22:00)). The explants were subcultured onto fresh media every four weeks throughout the entire experimental period.
2.2. Comparison of Callus Indcution Efficieny Using Leaf and Scape Exaplants
Young leaf segments of Datong Huanghua plants (1.0 cm × 0.5 cm) were inoculated on MS medium supplemented with 2.2 µM 6-BA and 9.0 µM 2,4-D for callus induction [
38]. The effect of TDZ on callus induction was tested at five different concentrations (0.5, 4.5, 11.4, 22.7 and 45.4 µM) in a completely randomized design (CRD); each treatment had 6 jars and each jar had 8 explants. The explants were incubated in the dark and then transferred to light following callus emergence. The rate of callus formation was calculated after 60 days of light culture, along with observations of callus division and differentiation.
When young scapes were used as the explants, scape segments with 0.5 cm in length obtained using Method 3 described below were inoculated in MS medium supplemented with different concentrations of TDZ (0.5, 4.5, 11.4, 22.7, 45.4, 68.1 and 90.8 µM), 6-BA (0, 2.2, 6.7 and 11.1 µM) and 2,4-D (0, 0.5, 2.3 and 4.5 µM) for a total of 23 treatments; each treatment had 9 jars and each jar had 5 explants. The callus formation rate was recorded after 30 days of dark culture and the shoot regeneration rate was recorded after 30 days of light culture. Observations of the callus growth status were also made.
From this experiment, the scape was shown to be a better explant than the leaf for callus induction and shoot regeneration. Thus, the scape was used as the explant in all subsequent investigations.
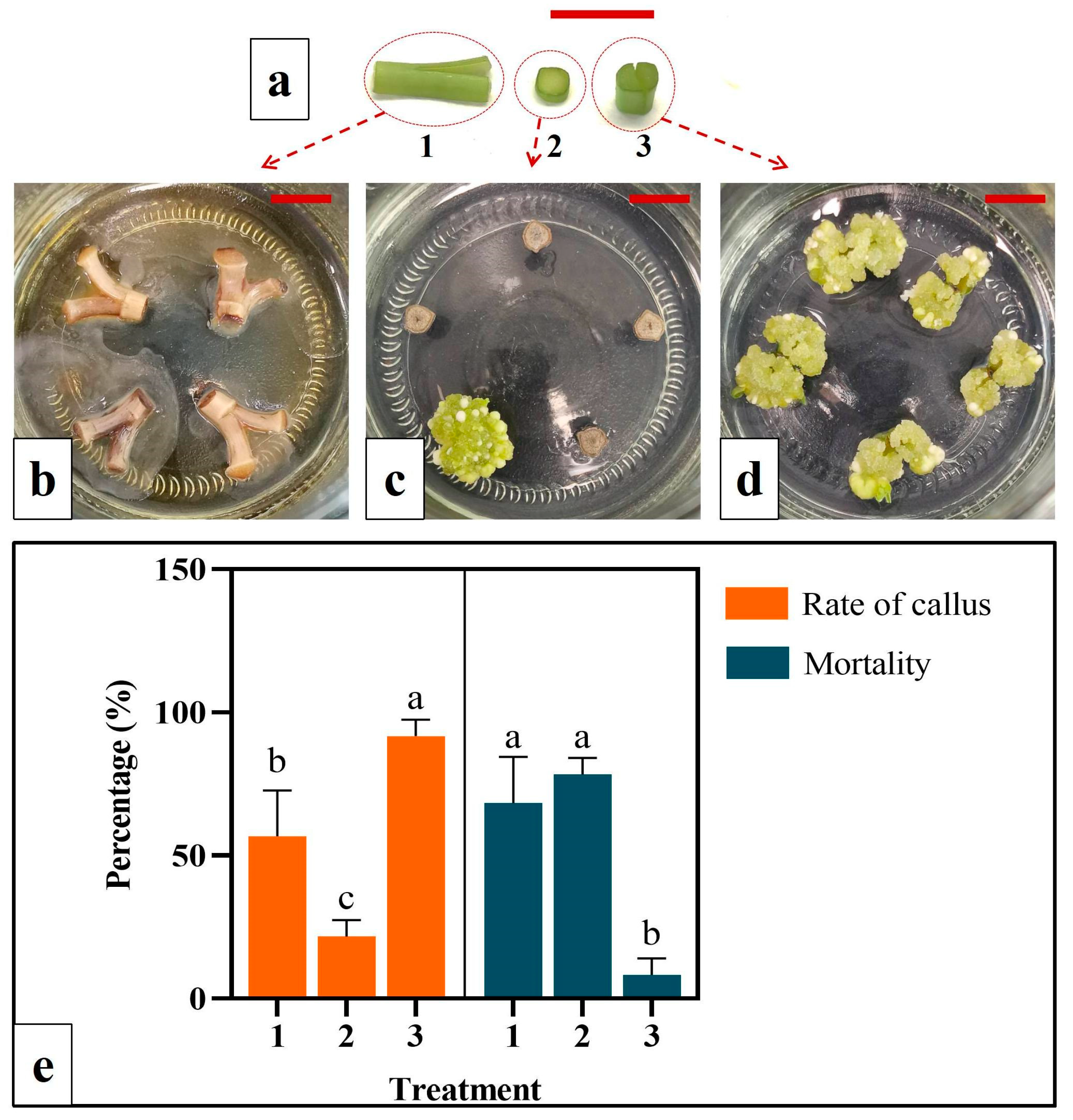
2.3. Evaluation of Callus Indcution Efficiency among Different Cutting Methods for Scape Explant Preparation
Three cutting/inoculation methods for scape tissue were evaluated for explant culture (detailed in the Results section). Method 1: A 1.0 cm long scape segment was cut into halves longitudinally [
30]. Each half was further cut at the lower end (root end) aiming to maximize the wounding. The cut surface was placed in contact with the medium for callus formation. Method 2: The scape was cut into 0.1 cm slices along their cross-sections, which were inoculated with the lower end facing upwards on the medium. Method 3: A 0.5 cm segment of the scape was cut longitudinally at its lower end and inoculated on the medium with the lower end facing upwards. Scape explants from different methods were cultivated in MS medium supplemented with 22.7 µM TDZ. For each treatment (cutting method), 12 jars were used with 5 explants within each jar. The explants were cultured in the dark and observed daily. The callus formation and degeneration rates were recorded after 30 days of culture.
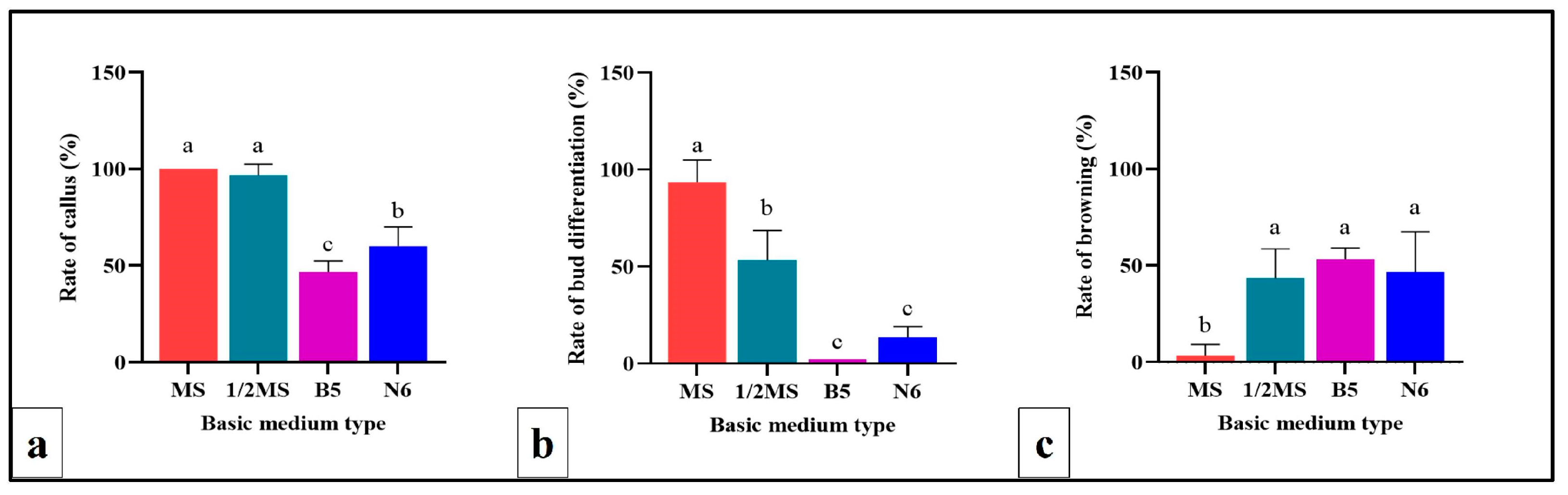
2.4. Evaluation of Basic Culture Media on Callus Indcution and Shoot Regeneration from Scape Explant
The efficiency of the four basic culture media for callus induction and shoot regeneration from scape explants was assessed. These media included MS, ½MS, Gamborg B-5 (B5) [
39] and Chu N6 (N6) [
40] (refer to
Supplemental File 1a for details). In each case, TDZ (22.7 µM TDZ) was added to the medium. Each treatment had 6 jars, with 5 scape explants inoculated per jar. The explants were cultured in the dark for culture condition and observed daily. Callus formation and browning rates were recorded after 30 days of dark culture. Subsequently, cultures were transferred to light conditions, with shoot regeneration rates recorded after an additional 30 days of light culture.
2.5. Callus Subculture, Shoot Regeneration and Rooting
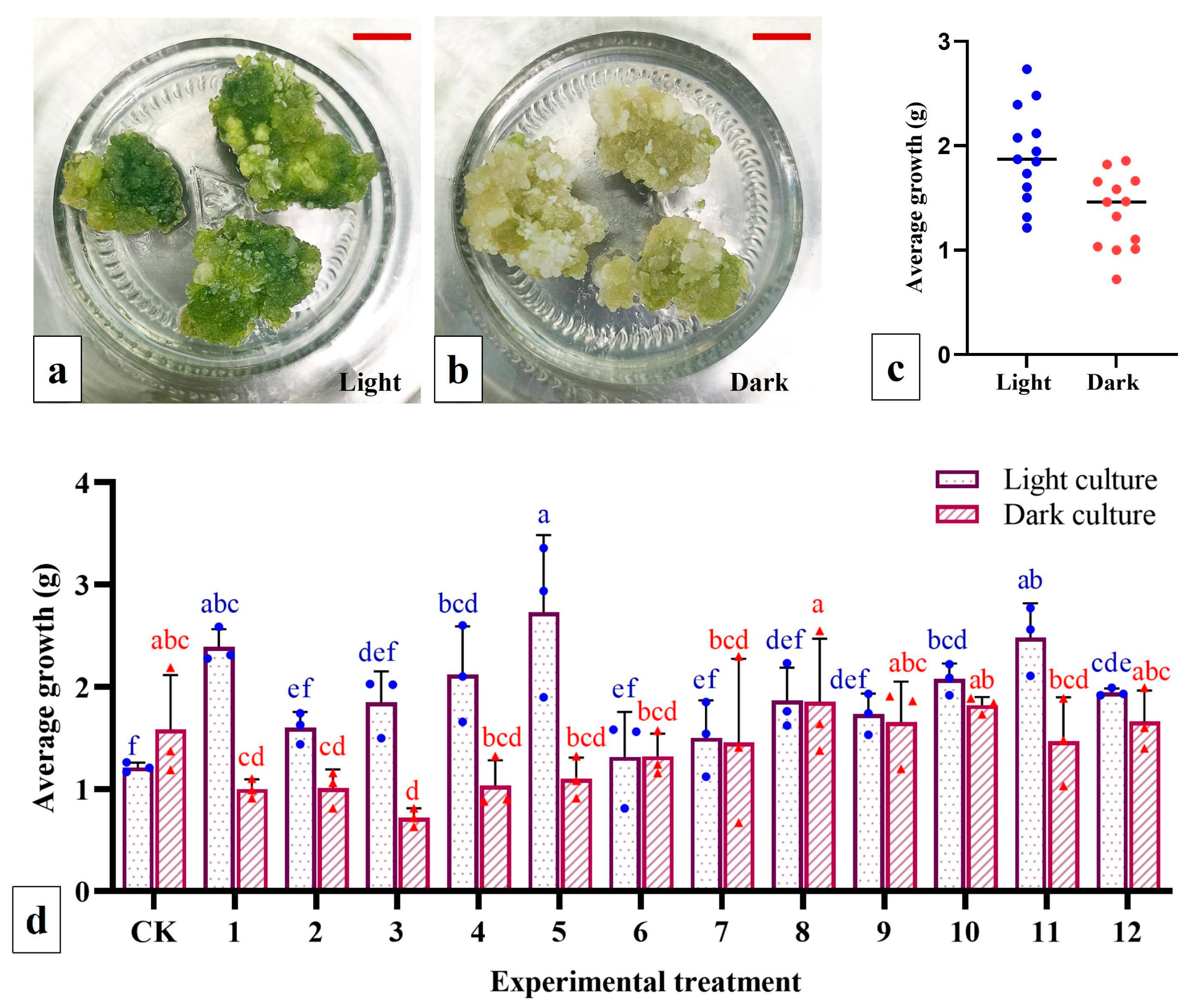
The embryogenic-like calli (light yellow in color and compact) generated from scape explants in the primary culture were selected for subculture/proliferation. These calli were segmented into pieces (~0.5 cm3), weighed and subsequently inoculated on MS proliferation medium containing different concentrations of NAA and 2,4-D (13 treatments in total; see below for details). Each treatment included 3 jars with three calli in each jar. Light and dark cultures were generated separately in the culture chamber, and the weight of the proliferated calli was recorded after 30 days. Additionally, the average growth rate of the calli was calculated.
The embryogenic-like calli of scape explants from the primary culture were selected and cut into small pieces (~0.5 cm3) and inoculated on MS shoot regeneration medium containing different concentrations of 6-BA (4.4, 8.9 and 13.3 µM) and NAA (2.7 and 5.4 µM) (for a total of 6 treatments). Each treatment had 9 jars; there were five calli in each jar. The shoot regeneration rate was recorded after 30 days of light cultivation.
Shoots with a height of ~3 cm in height were transplanted into rooting media including full-strength, ½ and ¼ MS, each of which was supplemented with different concentrations of NAA and 2,4-D (see below). For each treatment, there were 6 jars and 5 shoots in each jar. The rooting rate, the average number of roots and the rooting conditions of rooted seedlings were obtained after culture for 30 days.
2.6. Transplanting
Seedlings with well-developed root systems and robust growth were transplanted into plastic trays containing a mixture of peat, perlite and vermiculite as the growth substrate. Four treatments were evaluated for their effect on seedling establishment and growth: the ratios of peat, perlite and vermiculite were 3:1:1, 2:1:1 and 1:1:1, respectively, and the proportions of peat and perlite were 1:1. For each treatment, 50 tissue-cultured seedlings were evaluated. The seedling survival rate was recorded 40 days after transplanting.
2.7. Data Analysis
In different experiments, various parameters were calculated to evaluate the effects of treatments, described as follows:
Callus production rate = (# of calli formed/# explants inoculated) × 100%
Callus mortality rate = (# dead calli/total # of calli) × 100%
Callus browning rate = (# calli with browning/total number of calli) × 100%
Shoot regeneration rate = (# of explants with shoots/# calli inoculated) × 100%
Callus growth rate = Mean weight of calli at harvest − initial weight of the calli
Rooting rate = (# rooted seedlings/total # seedlings inoculated) × 100%
Average number of roots = total number of roots/total number of rooted seedlings
Seedling survival rate = (# seedlings survived/# seedlings transplanted) × 100%
All the experiments were analyzed with a completely randomized design (CRD). Except for the rooting substrate optimization experiment, the data from all the other experiments were analyzed with one-way ANOVA using SPSS Statistic 23.0. All plots and graphs were generated with Graph Pad Prism 9.0.0. The data are presented as the mean ± standard deviation (SD). Duncan’s multiple range test was used to assess significant differences between means at p < 0.05.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}