Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach

 ,
,  ,
,  , ,
, ,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Greenhouse Conditions
2.2. Experimental Design, Nitrogen Application, Cultural Practices and Biostimulant Characteristic
2.3. Yield and Biometric Assessments, Leaf Color Measurements and SPAD Index
2.4. Mineral Composition Analysis
2.5. Starch and Soluble Carbohydrates Analysis
2.6. Carotenoids Analysis
2.7. Polyphenols Analysis
2.8. Free and Total Amino Acids Analysis
2.9. Statistics, Principal Component Analysis, and Heat Map
3. Results
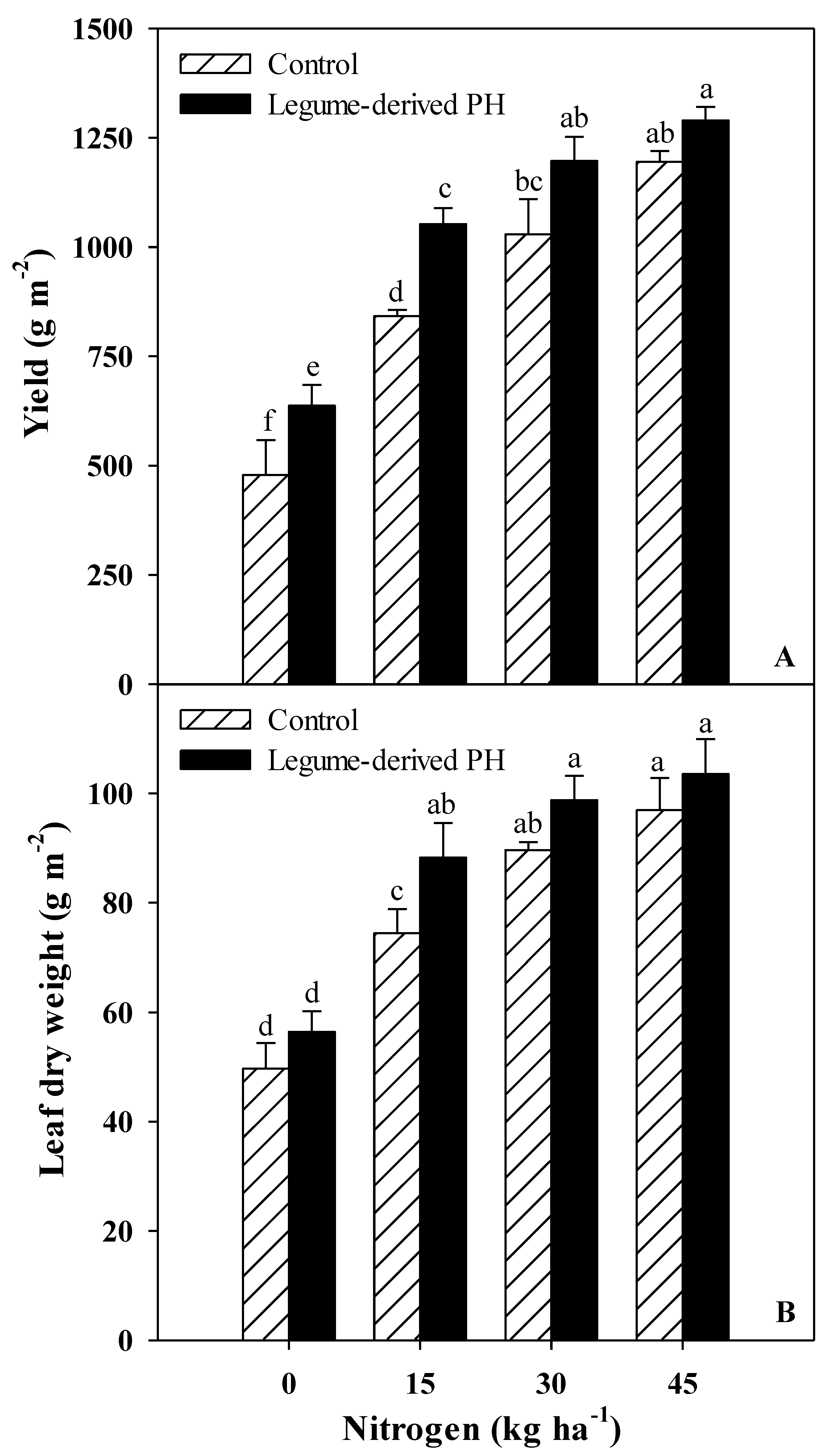
3.1. Marketable Yield and Morphometric Measurements
3.2. SPAD Index and Leaf Colorimetry
3.3. Protein Content and Leaf Mineral Composition
3.4. Carbohydrates, Carotenoids and Polyphenols Content
3.5. Free Amino Acid Contents
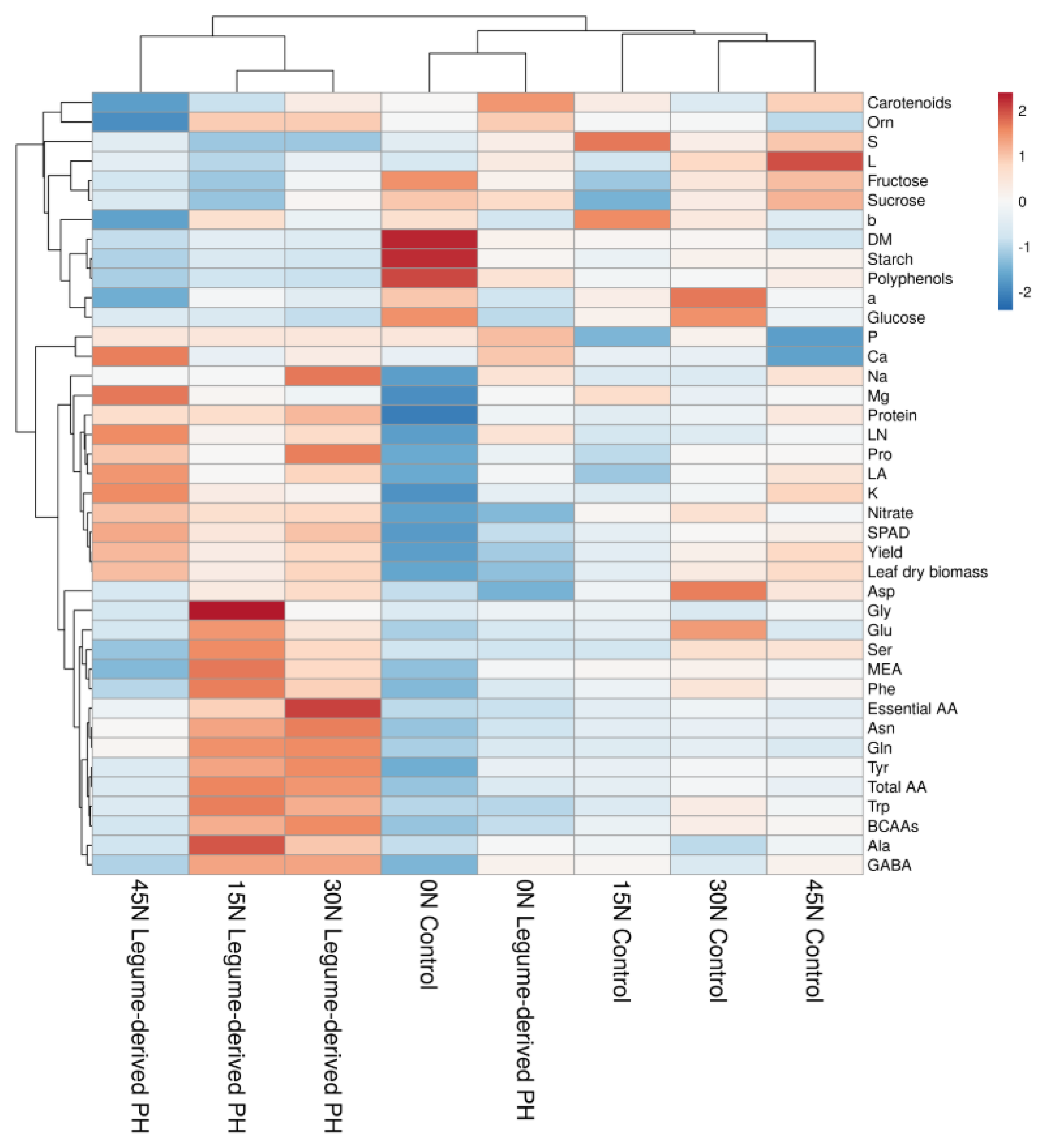
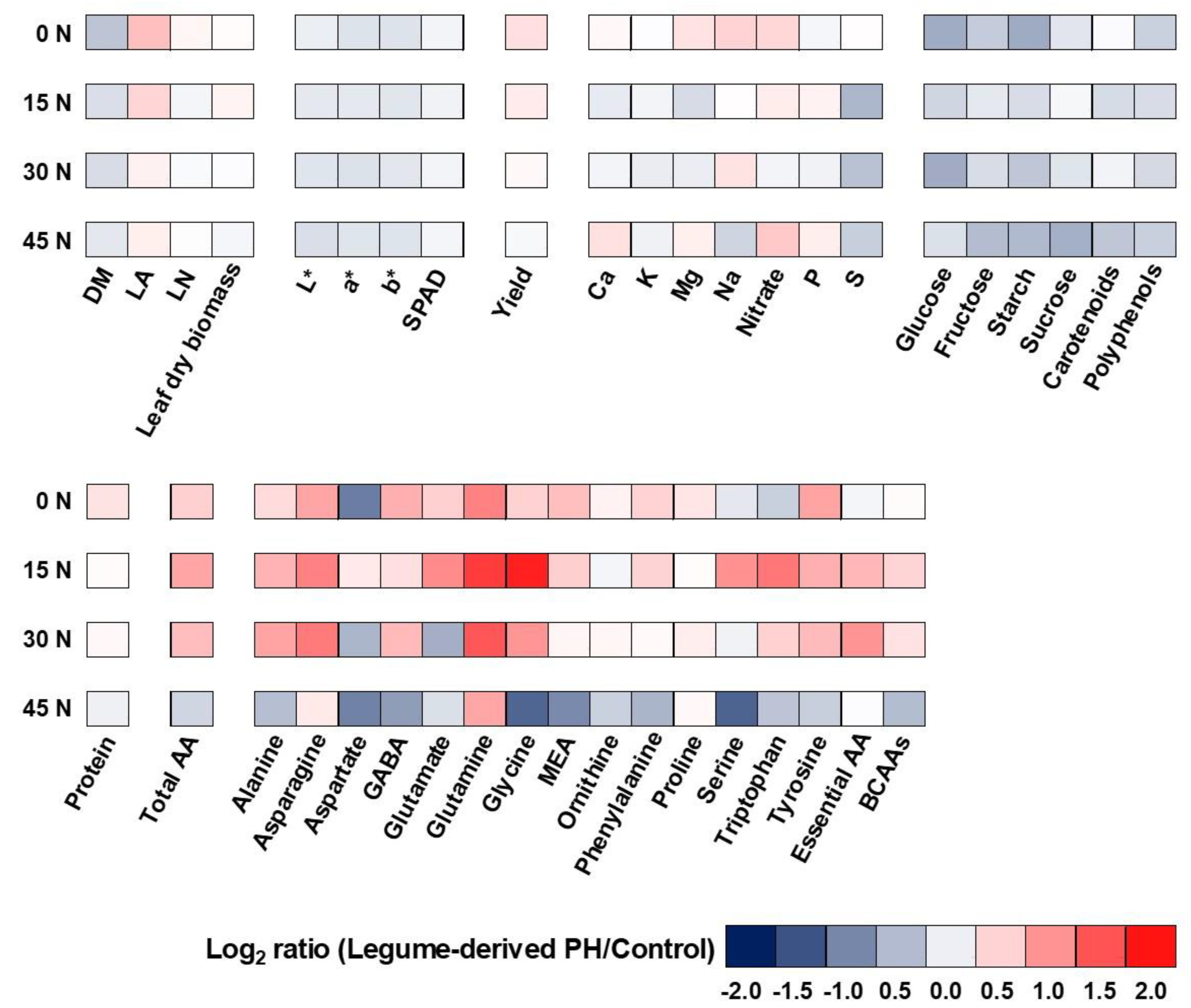
3.6. Heat Map Analysis
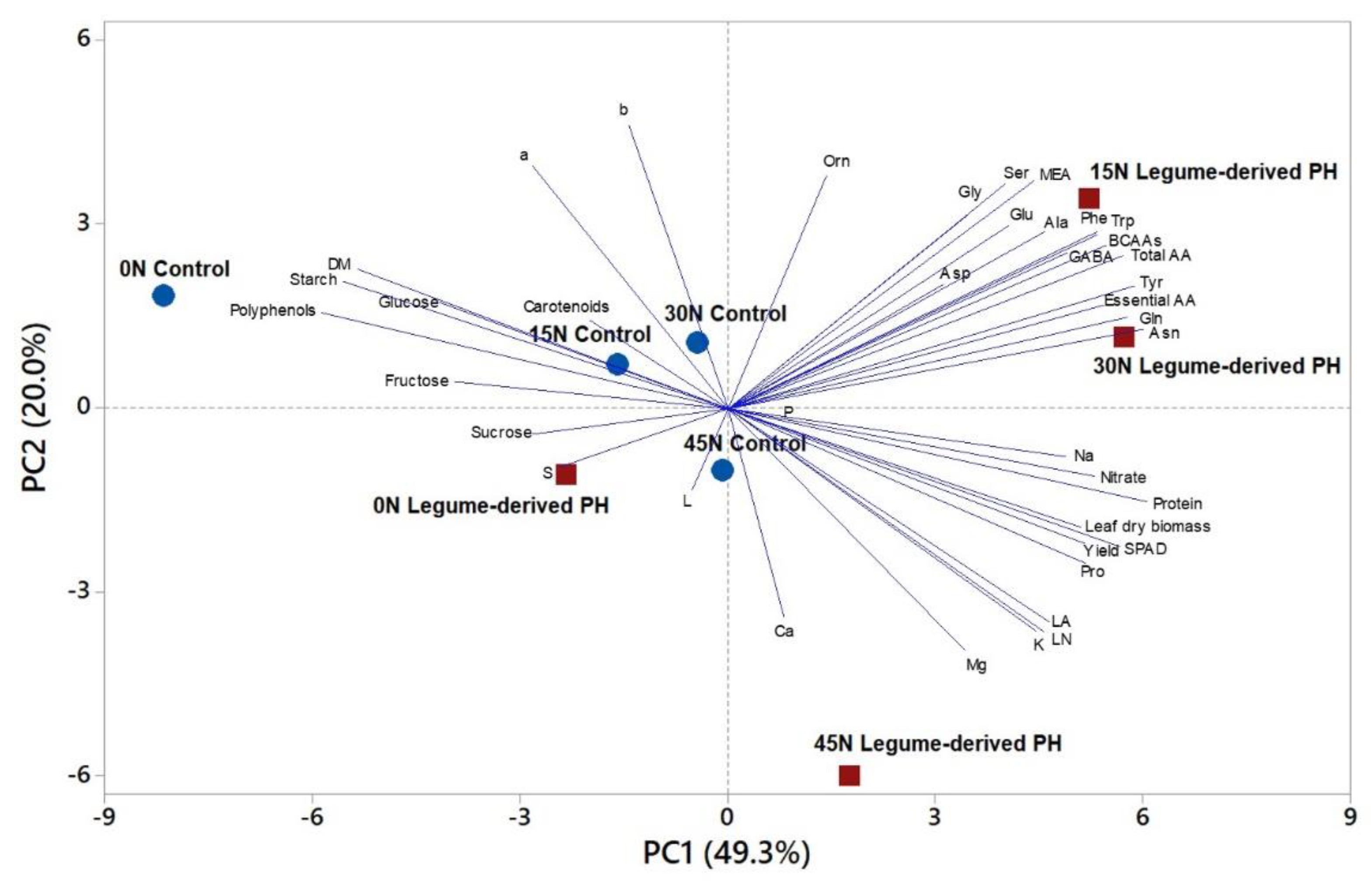
3.7. Principal Component Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Global Agriculture towards 2050, High-Level Expert Forum, How to Feed the World 2050, Rome 12–13 October 2009. Food and Agriculture Organization of United Nations (FAO). 2009. Available online: http://www.fao.org/fileadmin/templates/wsfs/docs/Issues_papers/HLEF2050_Global_Agriculture.pdf (accessed on 24 June 2019).
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. PNAS 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassaletta, L.; Billen, G.; Garnier, J.; Bouwman, L.; Velazquez, E.; Mueller, N.D.; Gerber, J.S. Nitrogen use in the global food system: Past trends and future trajectories of agronomic performance, pollution, trade, and dietary demand. Environ. Res. Lett. 2016, 11, 095007. [Google Scholar] [CrossRef]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. The influence of drip irrigation or subirrigation on zucchini squash grown in closed-loop substrate culture with high and low nutrient solution concentrations. HortScience 2009, 44, 306–311. [Google Scholar] [CrossRef]
- Colla, G.; Cardona Suárez, C.M.; Cardarelli, M.; Rouphael, Y. Improving nitrogen use efficiency in melon by grafting. HortScience 2010, 45, 559–565. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Mirabelli, C.; Cardarelli, M. Nitrogen-use efficiency traits of mini-watermelon in response to grafting and nitrogen-fertilization doses. J. Plant Nutr. Soil Sci. 2011, 174, 933–941. [Google Scholar] [CrossRef]
- Gupta, S.K.; Gupta, A.B.; Gupta, R. Pathophysiology of nitrate toxicity in humans in view of the changing trends of the global nitrogen cycle with special reference to India. In The Indian nitrogen Assessment; Abrol, Y.P., Adhya, T.K., Aneja, V.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 459–468. [Google Scholar] [CrossRef]
- Lassaletta, L.; Billen, G.; Grizzetti, B.; Anglade, J.; Garnier, J. 50 year trends in nitrogen use efficiency of world cropping systems: The relationship between yield and nitrogen input to cropland. Environ. Res. Lett. 2014, 9, 105011. [Google Scholar] [CrossRef]
- Sutton, M.A.; Bleeker, A.; Howard, C.M.; Bekunda, M.; Grizzetti, B.; de Vries, W.; van Grinsven, H.J.M.; Abrol, Y.P.; Adhya, T.K.; Billen, G.; et al. Our Nutrient World: The challenge to produce more food and energy with less pollution. Cent. Ecol. Hydrol. 2013, 8, 95–108. [Google Scholar]
- Aires, A.; Carvalho, R.; Rosa, E.A.S.; Saavedra, M.J. Effects of agriculture production systems on nitrate and nitrite accumulation on baby-leaf salads. Food Sci. Nutr. 2013, 1, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Kim, H.J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in fruits and vegetables. Sci. Hortic. 2018, 237, 231–238. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Santamaria, P. Nitrate in vegetables: Toxicity, content, intake and EC regulation. Sci. Food Agric. 2006, 86, 10–17. [Google Scholar] [CrossRef]
- Fallovo, C.; Rouphael, Y.; Rea, E.; Battistelli, A.; Colla, G. Nutrient solution concentration and growing season affect yield and quality of Lactuca sativa L. var. acephala in floating raft culture. J. Sci. Food Agric. 2009, 89, 1682–1689. [Google Scholar] [CrossRef]
- Hirel, B.; Le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.; Kopriva, S.; De Kok, L. Nutrient use efficiency in plants–Concepts and approaches. In Plant Ecophysiol; Springer International Publishing: Basel, Switzerland, 2014. [Google Scholar] [CrossRef]
- Erisman, J.W.; Leach, A.; Bleeker, A.; Atwell, B.; Cattaneo, L.; Galloway, J. An integrated approach to a nitrogen useefficiency (NUE) indicator for the food production–Consumption Chain. Sustainability 2018, 10, 925. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Carillo, P.; Fuggi, A.; Troccoli, A.; Woodrow, P. Metabolic profiling of cauliflower under traditional and reduced tillage systems. Aust. J. Crop Sci. 2013, 7, 1317–1323. [Google Scholar]
- Rouphael, Y.; Colla, G. Synergistic biostimulatory action: Designing the next generation of plant biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Fiorentino, N.; Ventorino, V.; Woo, S.L.; Pepe, O.; De Rosa, A.; Gioia, L.; Romano, I.; Lombardi, N.; Napolitano, M.; Colla, G.; et al. Trichoderma-based biostimulants modulate rhizosphere microbial populations and improve N uptake efficiency, yield and nutritional quality of leafy vegetables. Fron. Plant Sci. 2018, 9, 743. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 64, 145–158. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. HortScience 2017, 52, 1214. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical and metabolomic approaches. Front. Plant Sci. 2014, 5, 375. [Google Scholar] [CrossRef]
- Zhu, K.; Zhou, H.; Qian, H. Antioxidant and free radical-scavenging activities of wheat germ protein hydrolysates (WGPH) prepared with alcalase. Process Biochem. 2006, 41, 1296–1302. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. The use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar] [CrossRef]
- Li, X.; Zeng, R.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Du Jardin, P. Plant biostimulants: Definition; concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Carillo, P.; Morrone, B. Converting Residual Biomasses into Energy through Anaerobic Digestion. In Municipal Solid Waste: Management Strategies, Challenges and Future Directions; Tzortzakis, N., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2017; pp. 271–292. [Google Scholar]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef] [PubMed]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Biemond, H.; Vos, J.; Struik, P. Effects of nitrogen on accumulation and partitioning of dry matter and nitrogen of vegetables. Spinach. Neth. J. Agric. Sci. 1996, 44, 227–239. [Google Scholar]
- Smolders, E.; Buysse, J.; Merckx, R. Growth analysis of soil-grown spinach plants at different N-regimes. Plant Soil 1993, 154, 73–80. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M.; et al. A combined phenotypic and metabolomic approach for elucidating the biostimulant action of a plant-derived protein hydrolysate on tomato grown under limited water availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef]
- Kumar, P.; Lucini, L.; Rouphael, Y.; Cardarelli, C.; Kalunke, R.M.; Colla, G. Insight into the role of grafting and arbuscularmycorrhiza on cadmium stress tolerance in tomato. Front. Plant Sci. 2015, 6, 477. [Google Scholar] [CrossRef]
- Bremner, J.M. Total nitrogen. In Methods of Soil Analysis; Black, C.A., Evans, D.D., White, D.D., Ensminger, E., Clark, F.E., Eds.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 149–1178. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Attico, A.; Woodrow, P.; Oliva, M.A.; Fuggi, A.; Carillo, P. An improved fluorimetric HPLC method for quantifying tocopherols in Brassica rapa L. subsp. sylvestris after harvest. J. Food Comp. Anal. 2012, 27, 145–150. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar] [CrossRef]
- Carillo, P.; Raimondi, G.; Kyriacou, M.C.; Pannico, A.; El-Nakhel, C.; Cirillo, V.; Colla, G.; De Pascale, S.; Rouphael, Y. Morpho-physiological and homeostatic adaptive responses triggered by omeprazole enhance lettuce tolerance to salt stress. Sci. Hortic. 2019, 249, 22–30. [Google Scholar] [CrossRef]
- Ferchichi, S.; Hessini, K.; Dell’Aversana, E.; D’Amelia, L.; Woodrow, P.; Ciarmiello, L.F.; Fuggi, A.; Carillo, P. Hordeum vulgare and Hordeum maritimum respond to extended salinity stress displaying different temporal accumulation pattern of metabolites. Funct. Plant Biol. 2018, 11, 1096–1109. [Google Scholar] [CrossRef]
- Woodrow, P.; Ciarmiello, L.F.; Annunziata, M.G.; Pacifico, S.; Iannuzzi, F.; Mirto, A.; D’Amelia, L.; Dell’Aversana, E.; Piccolella, S.; Fuggi, A.; et al. Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiol. Plant 2017, 159, 290–312. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil. 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ciarmiello, L.F.; Piccirillo, P.; Carillo, P.; De Luca, A.; Woodrow, P. Determination of the genetic relatedness of fig (Ficuscarica L.) accessions using RAPD fingerprint and their agro-morphological characterization. S. Afr. J. Bot. 2015, 97, 40–47. [Google Scholar] [CrossRef]
- Mirto, A.; Iannuzzi, F.; Carillo, P.; Ciarmiello, L.F.; Woodrow, P.; Fuggi, A. Metabolic characterization and antioxidant activity in sweet cherry (Prunusavium L.) Campania accessions: Metabolic characterization of sweet cherry accessions. Food Chem. 2018, 240, 559–566. [Google Scholar] [CrossRef]
- Ganeshamurthy, A.N.; Rupa, T.R.; Kalaivanan, D.; Radha, T.K. Nitrogen management paradigm in horticulture systems in India. In The Indian Nitrogen Assessment; Abrol, Y.P., Adhya, T.K., Aneja, V.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 133–147. [Google Scholar] [CrossRef]
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving nitrogen use efficiency in crops for sustainable agriculture. Sustainability 2011, 9, 1452–1485. [Google Scholar] [CrossRef]
- Cirillo, C.; De Micco, V.; Arena, C.; Carillo, P.; Pannico, A.; De Pascale, S.; Rouphael, Y. Biochemical, physiological and anatomical mechanisms of adaptation of Callistemon citrinus and Viburnum lucidum to NaCl and CaCl2 salinization. Front. Plant Sci. 2019, 10, 742. [Google Scholar] [CrossRef]
- Rouphael, Y.; Raimondi, G.; Lucini, L.; Carillo, P.; Kyriacou, M.C.; Colla, G.; Cirillo, V.; Pannico, A.; El-Nakhel, C.; De Pascale, S. Physiological and metabolic responses triggered by omeprazole improve tomato plant tolerance to NaCl stress. Front. Plant Sci. 2018, 9, 249. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Carillo, P.; Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; dell’Aversana, E.; D’Amelia, L.; Colla, G.; Caruso, G.; De Pascale, S.; Rouphael, Y. Sensory and functional quality characterization of protected designation of origin ‘Piennolo del Vesuvio’ cherry tomato landraces from Campania-Italy. Food Chem. 2019, 292, 166–175. [Google Scholar] [CrossRef]
- Diab, H.; Limami, A.M. Reconfiguration of N metabolism upon hypoxia stress and recovery: Roles of alanine aminotransferase (AlaAT) and glutamate dehydrogenase (GDH). Plants 2016, 5, 25. [Google Scholar] [CrossRef]
- Carillo, P. GABA Shunt in durum wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef]
- Molina-Rueda, J.J.; Garrido-Aranda, A.; Gallardo, F. Glutamate decarboxylase. In Amino Acids in Higher Plants; D’Mello, J.P.F., Ed.; CABI: Wallingford, UK, 2015. [Google Scholar]
- Liu, C.; Zhao, L.; Yu, G. The dominant glutamic acid metabolic flux to produce γ-amino butyric acid over proline in Nicotianatabacum leaves under water stress relates to its significant role in antioxidant activity. J. Integr. Plant Biol. 2011, 53, 608–618. [Google Scholar] [CrossRef]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]
- Fukaki, H.; Tasaka, M. Hormone interactions during lateral root formation. Plant Mol. Biol. 2008, 69, 437. [Google Scholar] [CrossRef]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring HarbPerspect Biol. 2010, 2, a001537. [Google Scholar] [CrossRef]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of the tricarboxylic acid (TCA) cycle and nitrogen metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef]
- Buchanan, B.B. The carbon (formerly dark) reactions of photosynthesis. Photosynthesis Res. 2016, 128, 215–217. [Google Scholar] [CrossRef]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium deficiency in plants: An urgent problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Sudhakar, P.; Latha, P.; Reddy, P.V. Chapter 4–Photosynthetic rates. In Phenotyping Crop Plants for Physiological and Biochemicaltraits; Sudhakar, P., Latha, P., Reddyeds, P.V., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 33–39. [Google Scholar] [CrossRef]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Carillo, P.; Carfagna, S. Ammonium metabolism stimulation of glucose-6P dehydrogenase and phosphoenolpyruvate carboxylase in young barley roots. J. Plant Physiol. 1998, 153, 61–66. [Google Scholar] [CrossRef]
- Zhao, L.S.; Li, K.; Wang, Q.M.; Song, X.Y.; Su, H.N.; Xie, B.B.; Zhang, X.Y.; Huang, F.; Chen, X.L.; Zhou, B.C.; et al. Nitrogen starvation impacts the photosynthetic performance of Porphyridium cruentum as revealed by chlorophyll a fluorescence. Sci. Rep. 2017, 7, 8542. [Google Scholar] [CrossRef]
- Jones, C.G.; Hartley, S.E. A protein competition model of phenolic allocation. Oiko 1999, 86, 27–44. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein hydrolysate or plant extract-based biostimulants enhanced yield and quality performances of greenhouse perennial wall rocket grown in different seasons. Plants 2019, 8, 208. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Carillo, P.; Pizzolongo, F.; Romano, R.; Sifola, M.I. Chemical eustress elicits tailored responses and enhances the functional quality of novel food Perilla Frutescens. Molecules 2019, 24, 185. [Google Scholar] [CrossRef]
- Rossini, F.; Provenzano, M.E.; Sestili, F.; Ruggeri, R. Synergistic effect of sulfur and nitrogen in the organic and mineral fertilization of durum wheat: Grain yield and quality traits in the Mediterranean environment. Agronomy 2018, 8, 189. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | Leaf Number | Leaf Area | Leaf Dry Matter |
|---|---|---|---|
| (no. plant−1) | (cm2 plant−1) | (%) | |
| Nitrogen (kg ha−1) (N) | |||
| 0 | 6.9 ± 0.3 | 63.6 ± 6.7 c | 9.8 ± 0.6 a |
| 15 | 7.1 ± 0.1 | 68.4 ± 5.3 c | 8.6 ± 0.3 b |
| 30 | 7.3 ± 0.2 | 87.1 ± 4.9 b | 8.5 ± 0.3 b |
| 45 | 7.6 ± 0.2 | 97.5 ± 6.5 a | 8.1 ± 0.2 b |
| NS | *** | *** | |
| Biostimulant (B) | |||
| Control | 6.9 ± 0.1 | 68.3 ± 5.0 | 9.1 ± 0.4 |
| Legume-derived PH | 7.6 ± 0.1 | 90.0 ± 4.5 | 8.4 ± 0.1 |
| t-test | 0.002 | 0.004 | 0.094 |
| N × B | |||
| 0N Control | 6.4 ± 0.2 | 49.3 ± 0.9 | 10.7 ± 0.9 |
| 15N Control | 6.9 ± 0.1 | 56.8 ± 1.5 | 8.8 ± 0.5 |
| 30N Control | 7.0 ± 0.0 | 79.0 ± 4.5 | 8.8 ± 0.7 |
| 45N Control | 7.2 ± 0.1 | 88.2 ± 3.9 | 8.1 ± 0.4 |
| 0N Legume-derived PH | 7.5 ± 0.4 | 77.9 ± 4.7 | 8.9 ± 0.1 |
| 15N Legume-derived PH | 7.3 ± 0.1 | 79.9 ± 2.0 | 8.4 ± 0.3 |
| 30N Legume-derived PH | 7.6 ± 0.4 | 95.1 ± 5.7 | 8.3 ± 0.1 |
| 45N Legume-derived PH | 8.0 ± 0.1 | 106.9 ± 10.4 | 8.0 ± 0.3 |
| NS | NS | NS |
| Source of Variance | SPAD Index | L* | a* | b* | ||
|---|---|---|---|---|---|---|
| 33 DAS | 41 DAS | 48 DAS | ||||
| Nitrogen (kg ha−1) (N) | ||||||
| 0 | 39.92 ± 0.5 d | 40.61 ± 0.5 d | 41.11 ± 0.8 d | 39.22 ± 0.8 | 13.78 ± 0.3 | 18.61 ± 0.5 |
| 15 | 43.36 ± 0.5 c | 43.65 ± 0.6 c | 44.55 ± 0.4 c | 38.60 ± 0.4 | 13.79 ± 0.2 | 19.14 ± 0.5 |
| 30 | 44.56 ± 0.6 b | 45.51 ± 0.6 b | 46.12 ± 0.4 b | 39.58 ± 0.7 | 13.88 ± 0.6 | 18.68 ± 0.8 |
| 45 | 45.12 ± 0.6 a | 46.95 ± 0.5 a | 47.85 ± 0.4 a | 40.11 ± 1.4 | 13.50 ± 0.5 | 18.16 ± 1.1 |
| *** | *** | *** | NS | NS | NS | |
| Biostimulant (B) | ||||||
| Control | 42.08 ± 0.6 | 43.13 ± 0.8 | 43.94 ± 0.9 | 39.70 ± 0.7 | 13.93 ± 0.3 | 18.89 ± 0.6 |
| Legume-derived PH | 44.40 ± 0.7 | 45.23 ± 0.7 | 45.88 ± 0.7 | 39.05 ± 0.5 | 13.54 ± 0.2 | 18.41 ± 0.4 |
| t-test | 0.016 | 0.050 | 0.091 | 0.469 | 0.323 | 0.519 |
| N × B | ||||||
| 0N Control | 38.85 ± 0.3 | 39.57 ± 0.1 | 39.57 ± 0.7 | 38.78 ± 0.7 | 14.03 ± 0.4 | 18.93 ± 0.4 |
| 15N Control | 42.35 ± 0.3 | 42.39 ± 0.4 | 43.61 ± 0.2 | 38.71 ± 0.6 | 13.83 ± 0.0 | 19.34 ± 0.5 |
| 30N Control | 43.35 ± 0.1 | 44.36 ± 0.7 | 45.39 ± 0.2 | 40.09 ± 1.3 | 14.18 ± 1.1 | 18.83 ± 1.3 |
| 45N Control | 43.79 ± 0.3 | 46.19 ± 0.6 | 47.17 ± 0.5 | 41.23 ± 2.4 | 13.69 ± 1.0 | 18.44 ± 2.3 |
| 0N Legume-derived PH | 40.99 ± 0.5 | 41.64 ± 0.1 | 42.64 ± 0.1 | 39.66 ± 1.5 | 13.53 ± 0.4 | 18.28 ± 1.0 |
| 15N Legume-derived PH | 44.37 ± 0.4 | 44.91 ± 0.3 | 45.50 ± 0.2 | 38.49 ± 0.7 | 13.74 ± 0.4 | 18.93 ± 0.8 |
| 30N Legume-derived PH | 45.78 ± 0.2 | 46.65 ± 0.4 | 46.84 ± 0.6 | 39.06 ± 0.9 | 13.59 ± 0.6 | 18.53 ± 1.4 |
| 45N Legume-derived PH | 46.45 ± 0.4 | 47.71 ± 0.4 | 48.53 ± 0.1 | 38.99 ± 1.6 | 13.30 ± 0.3 | 17.88 ± 0.6 |
| NS | NS | NS | NS | NS | NS | |
| Source of Variance | Protein | Nitrate | Mineral Elements (g kg−1dw) | |||||
|---|---|---|---|---|---|---|---|---|
| (g kg−1dw) | (mg kg−1fw) | P | K | Ca | Mg | S | Na | |
| Nitrogen (kg ha−1) (N) | ||||||||
| 0 | 25.6 ± 1.8 b | 968 ± 132 b | 3.80 ± 0.2 | 91.3 ± 3.6 | 1.81 ± 0.2 | 8.88 ± 0.7 c | 1.26 ± 0.1 | 1.08 ± 0.1 |
| 15 | 29.9 ± 1.6 a | 3198 ± 254 a | 3.37 ± 0.2 | 97.8 ± 3.4 | 1.70 ± 0.1 | 10.57 ± 0.8 ab | 1.31 ± 0.1 | 1.13 ± 0.1 |
| 30 | 31.1 ± 1.7 a | 3620 ± 224 a | 3.66 ± 0.2 | 98.3 ± 3.2 | 1.73 ± 0.1 | 9.75 ± 0.4 bc | 1.22 ± 0.1 | 1.31 ± 0.1 |
| 45 | 31.4 ± 0.6 a | 3342 ± 348 a | 3.36 ± 0.2 | 106.2 ± 2.2 | 1.75 ± 0.1 | 11.17 ± 0.6 a | 1.30 ± 0.0 | 1.25 ± 0.2 |
| * | *** | NS | NS | NS | ** | NS | NS | |
| Biostimulant (B) | ||||||||
| Control | 27.4 ± 1.3 | 2478 ± 367 | 3.33 ± 0.1 | 95.8 ± 2.7 | 1.64 ± 0.0 | 9.60 ± 0.5 | 1.35 ± 0.1 | 1.10 ± 0.1 |
| Legume-derived PH | 31.6 ± 0.7 | 3044 ± 393 | 3.76 ± 0.1 | 101.0 ± 2.4 | 1.85 ± 0.1 | 10.59 ± 0.4 | 1.20 ± 0.0 | 1.28 ± 0.1 |
| t-test | 0.012 | 0.315 | 0.021 | 0.163 | 0.049 | 0.050 | 0.062 | 0.141 |
| N × B | ||||||||
| 0N Control | 22.3 ± 2.2 | 784 ± 303 | 3.67 ± 0.3 | 86.6 ± 6.5 | 1.67 ± 0.0 | 7.70 ± 0.9 b | 1.18 ± 0.1 | 0.90 ± 0.1 |
| 15N Control | 27.9 ± 2.3 | 2854 ± 387 | 3.06 ± 0.2 | 95.2 ± 4.2 | 1.70 ± 0.1 | 10.96 ± 1.4 a | 1.48 ± 0.1 | 1.06 ± 0.2 |
| 30N Control | 28.8 ± 2.8 | 3498 ± 487 | 3.58 ± 0.4 | 97.6 ± 4.1 | 1.68 ± 0.0 | 9.65 ± 0.8 ab | 1.34 ± 0.2 | 1.13 ± 0.2 |
| 45N Control | 30.9 ± 0.2 | 2677 ± 293 | 3.02 ± 0.1 | 104.0 ± 2.3 | 1.51 ± 0.1 | 10.08 ± 0.7 ab | 1.39 ± 0.0 | 1.31 ± 0.4 |
| 0N Legume-derived PH | 28.9 ± 0.5 | 1091 ± 92 | 3.93 ± 0.2 | 96.0 ± 0.5 | 1.94 ± 0.4 | 10.06 ± 0.6 ab | 1.34 ± 0.2 | 1.27 ± 0.1 |
| 15N Legume-derived PH | 32.0 ± 1.9 | 3543 ± 12 | 3.68 ± 0.2 | 100.5 ± 5.9 | 1.70 ± 0.1 | 10.18 ± 0.9 ab | 1.14 ± 0.1 | 1.20 ± 0.2 |
| 30N Legume-derived PH | 33.4 ± 0.9 | 3701 ± 282 | 3.74 ± 0.3 | 99.1 ± 5.8 | 1.77 ± 0.1 | 9.85 ± 0.3 ab | 1.10 ± 0.0 | 1.48 ± 0.1 |
| 45N Legume-derived PH | 32.0 ± 1.3 | 4007 ± 280 | 3.71 ± 0.2 | 108.5 ± 3.7 | 1.99 ± 0.1 | 12.26 ± 0.6 a | 1.21 ± 0.0 | 1.18 ± 0.0 |
| NS | NS | NS | NS | NS | * | NS | NS | |
| Source of Variance | Starch | Glucose | Fructose | Sucrose | Carotenoids | Polyphenols |
|---|---|---|---|---|---|---|
| (mg g−1dw) | (mg g−1dw) | (mg g−1dw) | (mg g−1dw) | (µg g−1dw) | (µg GAE g−1dw) | |
| Nitrogen (kg ha−1) (N) | ||||||
| 0 | 8.95 ± 1.2 | 33.85 ± 4.0 | 14.52 ± 1.4 | 3.03 ± 0.3 | 5.49 ± 0.3 | 32.16 ± 1.7 a |
| 15 | 6.81 ± 0.7 | 31.53 ± 3.9 | 11.20 ± 0.9 | 2.06 ± 0.4 | 5.12 ± 0.3 | 27.37 ± 1.0 b |
| 30 | 7.00 ± 1.0 | 33.95 ± 3.1 | 13.40 ± 1.0 | 2.74 ± 0.4 | 5.18 ± 0.3 | 27.45 ± 0.7 b |
| 45 | 6.78 ± 0.6 | 30.78 ± 2.4 | 13.41 ± 1.5 | 2.79 ± 0.3 | 5.08 ± 0.4 | 27.52 ± 1.4 b |
| NS | NS | NS | NS | NS | * | |
| Biostimulant (B) | ||||||
| Control | 8.15 ± 0.8 | 35.77 ± 2.4 | 13.92 ± 0.9 | 2.76 ± 0.3 | 5.28 ± 0.2 | 30.09 ± 1.2 |
| Legume-derived PH | 6.61 ± 0.5 | 29.29 ± 1.7 | 12.34 ± 0.8 | 2.55 ± 0.2 | 5.16 ± 0.2 | 27.16 ± 0.6 |
| t-test | 0.094 | 0.041 | 0.218 | 0.565 | 0.685 | 0.040 |
| N × B | ||||||
| 0N Control | 10.37 ± 2.2 | 39.18 ± 7.0 | 15.64 ± 2.5 | 3.06 ± 0.6 | 5.24 ± 0.5 | 34.16 ± 3.0 |
| 15N Control | 7.04 ± 1.3 | 33.17 ± 6.6 | 11.22 ± 1.1 | 1.99 ± 0.5 | 5.30 ± 0.3 | 28.29 ± 1.7 |
| 30N Control | 7.61 ± 1.5 | 39.23 ± 2.8 | 13.86 ± 1.8 | 2.77 ± 0.7 | 5.05 ± 0.6 | 28.58 ± 1.0 |
| 45N Control | 7.60 ± 0.7 | 31.48 ± 2.2 | 14.95 ± 1.5 | 3.21 ± 0.3 | 5.53 ± 0.5 | 29.31 ± 2.5 |
| 0N Legume-derived PH | 7.52 ± 0.8 | 28.52 ± 1.5 | 13.39 ± 1.8 | 3.00 ± 0.5 | 5.74 ± 0.2 | 30.15 ± 0.8 |
| 15N Legume-derived PH | 6.58 ± 0.8 | 29.89 ± 5.3 | 11.18 ± 1.6 | 2.13 ± 0.6 | 4.94 ± 0.4 | 26.45 ± 1.1 |
| 30N Legume-derived PH | 6.39 ± 1.4 | 28.67 ± 3.5 | 12.93 ± 1.2 | 2.70 ± 0.4 | 5.32 ± 0.3 | 26.32 ± 0.7 |
| 45N Legume-derived PH | 5.95 ± 0.7 | 30.09 ± 4.7 | 11.87 ± 2.6 | 2.38 ± 0.3 | 4.63 ± 0.4 | 25.73 ± 0.7 |
| NS | NS | NS | NS | NS | NS |
| Aminoacid | Nitrogen (kg ha−1) (N) | Biostimulant (B) | N × B | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (µmol g−1dw) | 0 | 15 | 30 | 45 | Sig. | Control | Legume-Derived PH | t-Test | 0N Control | 15N Control | 30N Control | 45N Control | 0N Legume-Derived PH | 15N Legume-Derived PH | 30N Legume-Derived PH | 45N Legume-Derived PH | Sig. |
| Alanine | 21.43 ± 2.8 | 31.55 ± 4.2 | 25.08 ± 4.4 | 21.18 ± 2.1 | NS | 20.70 ± 1.2 | 28.91 ± 3.1 | 0.023 | 18.23 ± 2.1 | 23.46 ± 2.4 | 17.65 ± 2.7 | 23.47 ± 1.2 | 24.63 ± 5.0 | 39.65 ± 4.2 | 32.5 ± 5.8 | 18.89 ± 3.9 | NS |
| Asparagine | 3.37 ± 0.5 c | 9.61 ± 1.8 ab | 10.57 ± 2.5 a | 7.05 ± 0.8 b | *** | 5.27 ± 0.7 | 10.03 ± 1.5 | 0.008 | 2.39 ± 0.4 | 5.99 ± 1.3 | 6.43 ± 1.5 | 6.25 ± 1.0 | 4.36 ± 0.2 | 13.22 ± 0.8 | 14.71 ± 3.4 | 7.84 ± 1.3 | NS |
| Aspartate | 3.17 ± 0.5 c | 6.99 ± 0.9 b | 10.45 ± 0.9 a | 6.46 ± 1.2 b | *** | 7.55 ± 1.0 | 5.98 ± 0.9 | 0.264 | 4.03 ± 0.3 | 6.19 ± 1.1 | 11.82 ± 1.2 | 8.14 ± 1.8 | 2.31 ± 0.5 | 7.78 ± 1.5 | 9.07 ± 1.1 | 4.77 ± 1 | NS |
| Mono ethanolamine | 3.74 ± 0.4 b | 5.91 ± 0.7 a | 5.40 ± 0.6 a | 3.69 ± 0.5 b | ** | 4.31 ± 0.4 | 5.06 ± 0.6 | 0.277 | 2.90 ± 0.3 c | 4.82 ± 0.7abc | 4.94 ± 0.8 abc | 4.58 ± 0.6 bc | 4.59 ± 0.2bc | 7.00 ± 0.9 a | 5.85 ± 1 ab | 2.81 ± 0.6c | * |
| Phenylalanine | 0.79 ± 0.1 b | 1.24 ± 0.1 a | 1.26 ± 0.1 a | 0.94 ± 0.1 ab | * | 0.98 ± 0.1 | 1.14 ± 0.1 | 0.271 | 0.65 ± 0.0 | 1.02 ± 0.2 | 1.17 ± 0.2 | 1.07 ± 0.2 | 0.93 ± 0.2 | 1.45 ± 0.1 | 1.35 ± 0.2 | 0.81 ± 0.1 | NS |
| GABA | 28.95 ± 4.0 | 41.62 ± 4.3 | 38.43 ± 6.1 | 30.75 ± 3.7 | NS | 30.73 ± 2.3 | 39.14 ± 4.0 | 0.081 | 21.25 ± 2.1 | 35.67 ± 3.0 | 29.28 ± 4.7 | 36.73 ± 2.5 | 36.64 ± 4.0 | 47.58 ± 6.9 | 47.58 ± 8.9 | 24.77 ± 5 | NS |
| Glycine | 1.27 ± 0.2 b | 3.28 ± 0.9 a | 1.39 ± 0.3 b | 1.18 ± 0.2 b | *** | 1.23 ± 0.1 | 2.33 ± 0.5 | 0.057 | 1.04 ± 0.2 bc | 1.38 ± 0.2 bc | 0.93 ± 0.1 bc | 1.57 ± 0.2 bc | 1.49 ± 0.3 bc | 5.18 ± 0.5 a | 1.84 ± 0.5 b | 0.8 ± 0.1 c | *** |
| Glutamate | 8.31 ± 1.0 b | 17.74 ± 3.6 a | 20.75 ± 2.5 a | 9.73 ± 1.3 b | *** | 13.01 ± 2.2 | 15.25 ± 2.2 | 0.481 | 6.80 ± 1.2 c | 11.42 ± 2.2 bc | 23.84 ± 4.0 a | 9.97 ± 1.7 bc | 9.81 ± 1.3 bc | 24.06 ± 4.6 a | 17.65 ± 2.4 ab | 9.48 ± 2.3 bc | ** |
| Glutamine | 9.54 ± 1.7 b | 31.45 ± 7.7 a | 33.30 ± 8.7 a | 19.00 ± 3.7 b | *** | 12.96 ± 2.0 | 33.68 ± 5.3 | 0.001 | 5.93 ± 0.5 c | 14.80 ± 3.0 bc | 17.61 ± 6.3 bc | 13.49 ± 2.0 bc | 13.16 ± 0.7 bc | 48.10 ± 3.3 a | 48.98 ± 9.6 a | 24.5 ± 5.8 b | * |
| Isoleucine | 1.57 ± 0.2 b | 2.65 ± 0.2 a | 3.01 ± 0.4 a | 2.21 ± 0.3 ab | * | 2.19 ± 0.2 | 2.53 ± 0.3 | 0.335 | 1.36 ± 0.1 | 2.23 ± 0.3 | 2.60 ± 0.5 | 2.56 ± 0.5 | 1.78 ± 0.3 | 3.06 ± 0.2 | 3.42 ± 0.6 | 1.87 ± 0.3 | NS |
| Histidine | 0.46 ± 0.1 b | 1.15 ± 0.1 a | 1.23 ± 0.2 a | 0.93 ± 0.1 a | *** | 0.79 ± 0.1 | 1.09 ± 0.1 | 0.066 | 0.32 ± 0.0 | 0.93 ± 0.1 | 1.00 ± 0.2 | 0.91 ± 0.1 | 0.60 ± 0.1 | 1.37 ± 0.1 | 1.46 ± 0.3 | 0.94 ± 0.1 | NS |
| Leucine | 2.54 ± 0.3 | 3.26 ± 0.3 | 3.46 ± 0.4 | 2.65 ± 0.2 | NS | 2.92 ± 0.2 | 3.04 ± 0.3 | 0.719 | 2.73 ± 0.6 | 2.85 ± 0.4 | 3.20 ± 0.3 | 2.88 ± 0.3 | 2.36 ± 0.2 | 3.66 ± 0.1 | 3.72 ± 0.7 | 2.41 ± 0.4 | NS |
| Lysine | 2.48 ± 0.3 | 2.68 ± 0.1 | 2.39 ± 0.2 | 1.96 ± 0.2 | NS | 2.34 ± 0.1 | 2.41 ± 0.2 | 0.715 | 2.39 ± 0.3 | 2.55 ± 0.0 | 2.34 ± 0.2 | 2.08 ± 0.1 | 2.56 ± 0.5 | 2.81 ± 0.1 | 2.44 ± 0.3 | 1.84 ± 0.3 | NS |
| Methionine | 0.33 ± 0.1 | 0.51 ± 0.1 | 0.50 ± 0.1 | 0.37 ± 0.0 | NS | 0.34 ± 0.0 | 0.51 ± 0.1 | 0.009 | 0.21 ± 0.0 | 0.37 ± 0.0 | 0.38 ± 0.1 | 0.39 ± 0.1 | 0.46 ± 0.1 | 0.65 ± 0.0 | 0.61 ± 0.1 | 0.34 ± 0.1 | NS |
| Ornhitine | 0.72 ± 0.1 | 0.74 ± 0.0 | 0.71 ± 0.0 | 0.58 ± 0.0 | NS | 0.66 ± 0.0 | 0.72 ± 0.1 | 0.346 | 0.66 ± 0.0 | 0.72 ± 0.0 | 0.65 ± 0.0 | 0.61 ± 0.1 | 0.79 ± 0.2 | 0.76 ± 0.0 | 0.77 ± 0.1 | 0.54 ± 0.1 | NS |
| Proline | 1.82 ± 0.1 c | 1.94 ± 0.1 bc | 2.33 ± 0.2 a | 2.27 ± 0.2 ab | * | 1.90 ± 0.1 | 2.28 ± 0.1 | 0.011 | 1.60 ± 0.1 | 1.82 ± 0.1 | 2.09 ± 0.2 | 2.09 ± 0.1 | 2.05 ± 0.2 | 2.07 ± 0.2 | 2.57 ± 0.1 | 2.44 ± 0.3 | NS |
| Serine | 5.72 ± 0.5 b | 8.76 ± 1.7 a | 9.53 ± 0.6 a | 6.84 ± 1.3 ab | * | 7.49 ± 0.7 | 7.93 ± 1.0 | 0.728 | 5.74 ± 0.9 bc | 5.78 ± 1.1 bc | 9.33 ± 0.7 ab | 9.11 ± 1.6 ab | 5.69 ± 0.7 bc | 11.73 ± 2.0 a | 9.74 ± 1.2 ab | 4.57 ± 1 c | ** |
| Tyrosine | 0.84 ± 0.1 b | 1.43 ± 0.2 a | 1.53 ± 0.2 a | 1.08 ± 0.1 ab | * | 0.99 ± 0.1 | 1.45 ± 0.2 | 0.020 | 0.59 ± 0.0 | 1.05 ± 0.2 | 1.17 ± 0.2 | 1.15 ± 0.2 | 1.09 ± 0.2 | 1.82 ± 0.1 | 1.89 ± 0.4 | 1 ± 0.2 | NS |
| Threonine | 1.69 ± 0.4 c | 6.35 ± 2.0 b | 9.42 ± 3.0 a | 4.58 ± 1.6 b | *** | 1.87 ± 0.2 | 9.15 ± 1.6 | 0.000 | 1.00 ± 0.1 d | 2.16 ± 0.2 d | 2.88 ± 0.3 d | 1.45 ± 0.2 d | 2.39 ± 0.5 d | 10.55 ± 1.4 b | 15.96 ± 1.3 a | 7.7 ± 1.5 c | *** |
| Tryptophan | 0.21 ± 0.0 b | 0.58 ± 0.1 a | 0.61 ± 0.1 a | 0.36 ± 0.1b | *** | 0.37 ± 0.0 | 0.51 ± 0.1 | 0.153 | 0.22 ± 0.0 cd | 0.35 ± 0.1 cd | 0.50 ± 0.1 bc | 0.39 ± 0.1 cd | 0.19 ± 0.0 d | 0.81 ± 0.2 a | 0.72 ± 0.1 ab | 0.33 ± 0.1 cd | * |
| Valine | 2.06 ± 0.2 c | 3.73 ± 0.4 ab | 4.18 ± 0.6 a | 2.94 ± 0.3 bc | ** | 2.82 ± 0.3 | 3.64 ± 0.4 | 0.106 | 1.67 ± 0.2 | 2.91 ± 0.4 | 3.43 ± 0.7 | 3.26 ± 0.5 | 2.46 ± 0.3 | 4.55 ± 0.3 | 4.93 ± 0.8 | 2.63 ± 0.4 | NS |
| Total amino acids | 102 ± 10 b | 184 ± 25 a | 189 ± 24 a | 128 ± 12 b | *** | 124 ± 10 | 178 ± 19 | 0.017 | 83 ± 7 c | 131 ± 14 bc | 146 ± 22 b | 135 ± 14 bc | 120 ± 11 bc | 238 ± 9 a | 232 ± 23 a | 121 ± 22 bc | * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy 2019, 9, 450. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9080450
Carillo P, Colla G, Fusco GM, Dell’Aversana E, El-Nakhel C, Giordano M, Pannico A, Cozzolino E, Mori M, Reynaud H, et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy. 2019; 9(8):450. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9080450
Chicago/Turabian StyleCarillo, Petronia, Giuseppe Colla, Giovanna Marta Fusco, Emilia Dell’Aversana, Christophe El-Nakhel, Maria Giordano, Antonio Pannico, Eugenio Cozzolino, Mauro Mori, Hélène Reynaud, and et al. 2019. "Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach" Agronomy 9, no. 8: 450. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9080450