Yield and Nutritional Quality of Vesuvian Piennolo Tomato PDO as Affected by Farming System and Biostimulant Application

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growing Conditions and Experimental Protocol
2.2. Yield, Biometric Assessments and Leaf Color Measurements
2.3. Juice Total Soluble Solids and Fruit Dry Matter Content
2.4. Mineral and Organic Acids Analysis
2.5. Antioxidant Activity Analysis
2.6. Antioxidant Molecules Analysis
2.7. Statistical Processing
3. Results and Discussion
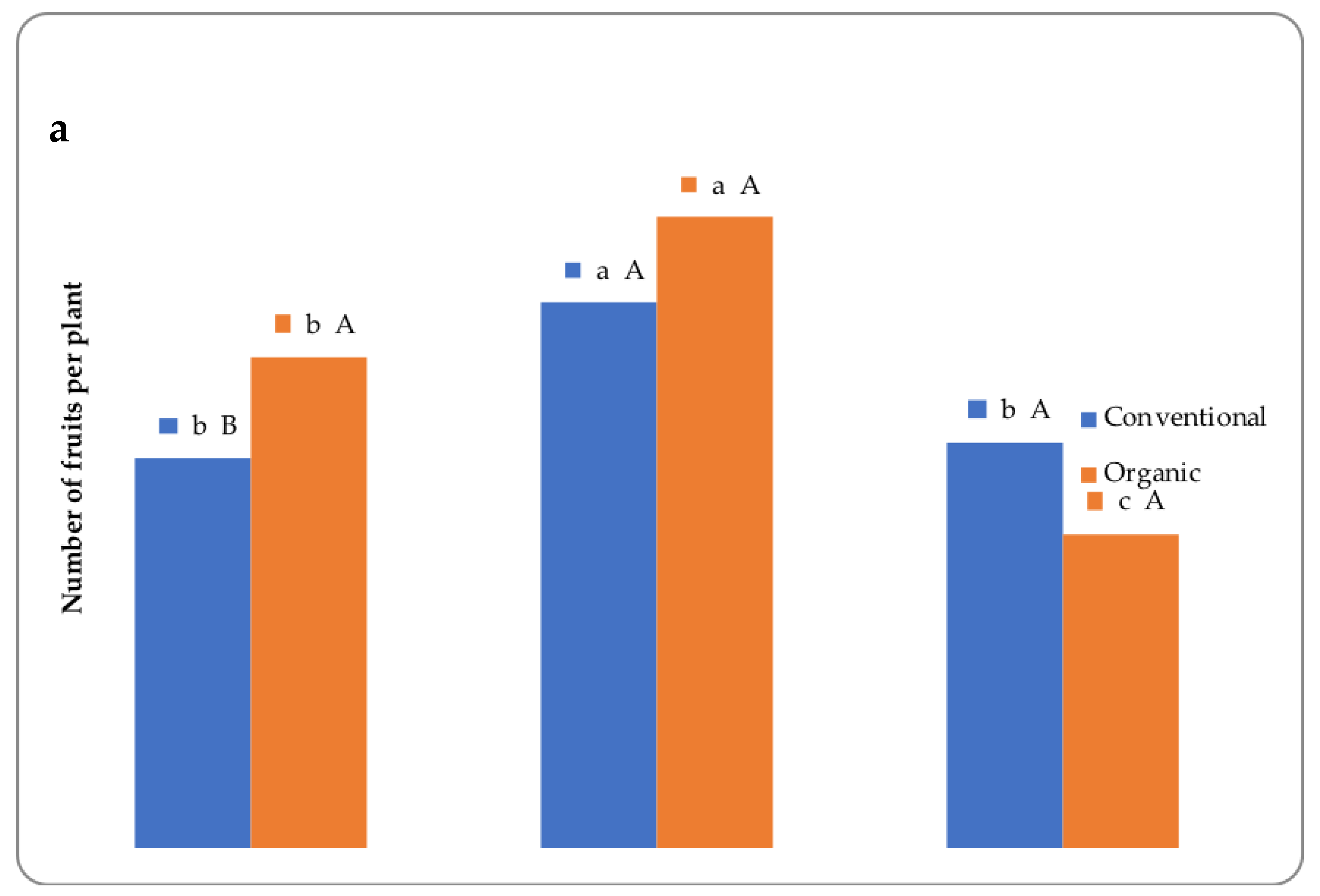
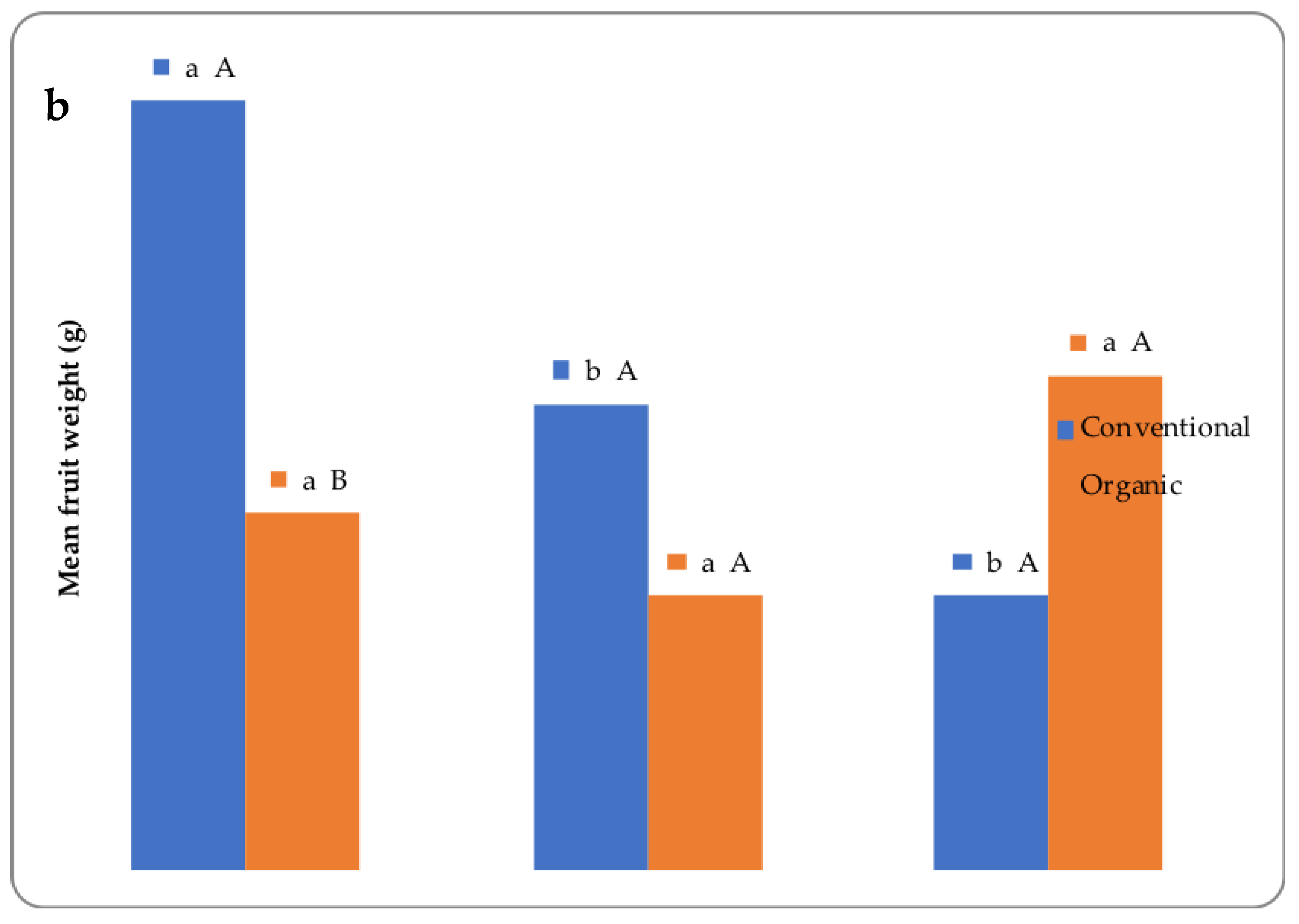
3.1. Yield and Morphometric Measurements
3.2. Fruit Colorimetry, Nutritional Quality and Mineral Profile
3.3. Antioxidant Activity and Bioactive Content
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dorais, M.; Alsanius, B. Recent advances in organic horticulture technology and management. Sci. Hortic. 2016, 208, 1–2. [Google Scholar] [CrossRef]
- Willer, H.; Lernoud, J. The World of Organic Agriculture—Statistics and Emerging Trends 2016. FiBL-IFOAM Rep. Bonn. Available online: https://shop.fibl.org/fileadmin/documents/shop/1698-organicworld-2016.pdf (accessed on 23 August 2019).
- Dorais, M. Organic production of vegetables: State of the art and challenges. Can. J. Plant Sci. 2007, 85, 1055–1066. [Google Scholar] [CrossRef]
- Ponisio, L.C.; M’Gonigle, L.K.; Mace, K.C.; Palomino, J.; de Valpine, P.; Kremen, C. Diversification practices reduce organic to conventional yield gap. Proc. R. Soc. B 2015, 282, 20141396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seufert, V.; Ramankutty, N.; Foley, J.A. Comparing the yields of organic and conventional agriculture. Nature 2012, 485, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Orsini, F.; Maggio, A.; Rouphael, Y.; De Pascale, S. ‘Physiologically quality’ of organically grown vegetables. Sci. Hortic. 2016, 208, 131–139. [Google Scholar] [CrossRef]
- De Ponti, T.; Rijk, B.; Van Ittersum, M.K. The crop yield gap between organic and conventional agriculture. Agric. Syst. 2012, 108, 1–9. [Google Scholar] [CrossRef]
- Berry, P.M.; Sylvester-Bradley, R.; Philipps, L.; Hatch, D.J.; Cuttle, S.P.; Rayns, F.W.; Goslin, P. Is the productivity of organic farms restricted by the supply available nitrogen? Soil Use Manag. 2002, 18, 248–255. [Google Scholar] [CrossRef]
- Lester, G.E.; Saftner, R.A. Organically versus conventionally grown produce: Common production inputs, nutritional quality, and nitrogen delivery between the two systems. J. Agric. Food. Chem. 2011, 59, 10401–10406. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y. Biostimulants in horticulture. Sci. Hortic. 2015, 196, 1–2. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic action of a microbial-based biostimulant and a plant derived-protein hydrolysate enhances lettuce tolerance to alkalinity and salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Colla, G. Role of arbuscular mycorrhizal fungi in alleviating the adverse effects of acidity and aluminium toxicity in zucchini squash. Sci. Hortic. 2015, 188, 97–105. [Google Scholar] [CrossRef]
- Schaafsma, G. Safety of protein hydrolysates, fractions thereof and bioactive peptides in human nutrition. Eur. J. Clin. Nutr. 2009, 63, 1161–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerdán, M.; Sánchez-Sánchez, A.; Jordá, J.D.; Juárez, M.; Sánchez-Andreu, J. Effect of commercial amino acids on iron nutrition of tomato plants grown under lime-induced iron deficiency. J. Plant Nutr. Soil Sci. 2013, 176, 859–866. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar applications of protein hydrolysate, plant and seaweed extracts increase yield but differentially modulate fruit quality of greenhouse tomato. Hort. Sci. 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical and metabolomic approaches. Front. Plant Sci. 2014, 5, 375. [Google Scholar] [CrossRef]
- Botta, A. Enhancing plant tolerance to temperature stress with amino acids: An approach to their mode of action. Acta Hortic. 2013, 1009, 29–35. [Google Scholar] [CrossRef]
- Colla, G.; Svecova, E.; Rouphael, Y.; Cardarelli, M.; Reynaud, H.; Canaguier, R.; Planques, B. Effectiveness of a plantderived protein hydrolysate to improve crop performances under different growing conditions. Acta Hortic. 2013, 1009, 175–179. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The effect of a plant-derived protein hydrolysate on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein hydrolysate stimulates growth in tomato coupled with N-dependent gene expression involved in N assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the biostimulant action of vegetal-derived protein hydrolysates by high-throughput plant phenotyping and metabolomics: A case study on tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of TCA cycle and N metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activities of two protein hydrolysates on the growth and nitrogen metabolism in maize seedlings. J. Plant. Nutr. Soil. Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Matsumiya, Y.; Kubo, M. Soybean. In Soybean and Nutrition; El-Shemy, H., Ed.; InTech Europe: Rijeka, Croatia, 2011; pp. 215–230. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hort. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Caruso, G.; Stoleru, V.; De Pascale, S.; Cozzolino, E.; Pannico, A.; Giordano, M.; Teliban, G.; Cuciniello, A.; Rouphael, Y. Production, leaf quality and antioxidants of perennial wall rocket as affected by crop cycle and mulching type. Agronomy 2019, 9, 194. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.; Bonini, P.; Colla, G. Plant and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soterious, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved abts radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fogliano, V.; Verde, V.; Randazzo, G.; Ritieni, A. Method for measuring antioxidant activity and its application to monitoring the antioxidant capacity of wines. J. Agric. Food Chem. 1999, 47, 1035–1040. [Google Scholar] [CrossRef] [PubMed]
- Kampfenkel, K.; Van Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin–Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Sadler, G.; Davis, J.; Dezman, D. Rapid extraction of lycopene and β-carotene from reconstituted tomato paste and pink grapefruit homogenates. J. Food Sci. 1990, 55, 1460–1461. [Google Scholar] [CrossRef]
- Caruso, G.; Conti, S.; La Rocca, G. Influence of crop cycle and nitrogen fertilizer form on yield and nitrate content in dierent species of vegetables. Adv. Hortic. Sci. 2011, 25, 81–89. [Google Scholar]
- Parrado, J.; Bautista, J.; Romero, E.J.; García-Martínez, A.M.; Friaza, V.; Tejada, M. Production of a carob enzymatic extract: Potential use as a biofertilizer. Bioresour. Technol. 2008, 99, 2312–2318. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Cavani, L.; Ciavatta, C. Attività biostimolante negli idrolizzanti proteici. L’Informatore Agrario 2007, 44, 46–52. [Google Scholar]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant application with a tropical plant extract enhances Corchorus olitorius adaptation to sub-optimal nutrient regimens by improving physiological parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Rengasamy, K.R.R.; Pendota, S.C.; Gruz, J.; Plaˇcková, L.; Novák, O.; Doležal, K.; Van Staden, J. Bioactive molecules derived from smoke and seaweed Ecklonia maxima showing phytohormone-like activity in Spinacia oleracea L. New Biotechnol. 2019, 48, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Luziatelli, F.; Ficca, A.G.; Colla, G.; Baldassarre Švecová, E.; Ruzzi, M. Foliar application of vegetal-derived bioactive compounds stimulates the growth of beneficial bacteria and enhances microbiome biodiversity in lettuce. Front. Plant. Sci. 2019, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Villari, G.; Borrelli, C.; Russo, G. Effects of crop method and harvest seasons on yield and quality of green asparagus under tunnel in southern Italy. Adv. Hort. Sci. 2012, 26, 51–58. [Google Scholar]
- Golubkina, N.A.; Seredin, T.M.; Antoshkina, M.S.; Kosheleva, O.V.; Teliban, G.C.; Caruso, G. Yield, quality, antioxidants and elemental composition of new leek cultivars under greenhouse organic or conventional system. Horticulturae 2018, 4, 39. [Google Scholar] [CrossRef]
- Kader, A.A. Postharvest Technology of Horticultural Crops; University of California, Division of Agriculture and Natural Resources Publication: Davis, CA, USA, 2002; Volume 3311, p. 535. [Google Scholar]
- Levander, O.A. Fruit and vegetable contributions to dietary mineral intake in human health and disease. Hort. Sci. 1990, 25, 1486–1488. [Google Scholar]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A vegetal biopolymer-based biostimulant promoted root growth in melon while triggering brassinosteroids and stress-related compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-wide identification of differentially expressed genes in Solanum Lycopersicon L. in response to an alfalfa-protein hydrolysate using microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef]
- Conti, S.; Villari, G.; Faugno, S.; Melchionna, G.; Somma, S.; Caruso, G. Effects of organic vs. conventional farming system on yield and quality of strawberry grown as an annual or biennial crop in southern Italy. Sci. Hortic. 2014, 180, 63–71. [Google Scholar] [CrossRef]
- Trevisan, S.; Manoli, A.; Quaggiotti, S.A. Novel biostimulant, belonging to protein hydrolysates, mitigates abiotic stress effects on maize seedlings grown in hydroponics. Agronomy 2019, 9, 28. [Google Scholar] [CrossRef]
- Wang, W.; Xia, M.X.; Chen, J.; Yuan, R.; Deng, F.N.; Shen, F.F. Gene expression characteristics and regulation mechanisms of superoxide dismutase and its hysiological roles in plants under stress. Biochem. 2016, 81, 465–480. [Google Scholar] [CrossRef]
- Hyun, M.W.; Yun, Y.H.; Kim, J.Y.; Kim, S.H. Fungal and plant phenylalanine ammonialyase. Mycobiol. 2011, 39, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source of Variance | LAI | Aerial Biomass | Marketable Yield | Marketable Fruits | |
|---|---|---|---|---|---|
| Mean Weight | Number | ||||
| (m2 m−2) | (g dw m−2) | (t ha−1) | (g per Fruit) | No. per Plant | |
| Year (Y) | ns | ns | ns | ns | ns |
| Farming system (F) | ns | ns | ns | ns | ns |
| Biostimulant (B) | * | * | * | ns | * |
| Y × F | ns | ns | ns | ns | ns |
| Y × B | ns | ns | ns | ns | ns |
| F × B | ns | ns | ns | * | * |
| Y × F× B | ns | ns | ns | ns | ns |
| Year (Y) | |||||
| 2016 | 4.92 | 350.3 | 14.5 | 15.5 | 23.1 |
| 2017 | 5.00 | 375.5 | 15.0 | 15.7 | 24.3 |
| Farming system (F) | |||||
| Organic | 5.05 | 368.0 | 14.7 | 15.4 | 24.0 |
| Conventional | 4.87 | 357.8 | 14.8 | 15.8 | 23.3 |
| Biostimulant formulate (B) | |||||
| Control | 4.54 c | 336.4 b | 13.4 c | 15.4 | 21.8 b |
| Legume-derived protein hydrolysate (PH) | 5.31 a | 385.5 a | 15.9 a | 15.4 | 26.0 a |
| Tropical plant extract (PE) | 5.03 b | 366.8 a | 14.9 b | 16.0 | 23.2 b |
| Source of Variance | Dry Matter | TSS | Fruit Colorimetry | Organic Acids (g kg−1 dw) | |||||
|---|---|---|---|---|---|---|---|---|---|
| (%) | (°Brix) | L * | A * | B * | Malate | Oxalate | Citrate | Isocitrate | |
| Farming system | ns | ns | ns | * | * | * | * | ns | * |
| Biostimulant | ns | * | * | * | ns | * | * | * | * |
| F × B | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| Farming system | |||||||||
| Organic | 8.6 | 7.3 | 40.8 | 31.3 | 20.4 | 13.4 | 1.25 | 43.8 | 0.54 |
| Conventional | 8.9 | 7.5 | 43.6 | 34.4 | 23.5 | 10.9 | 1.07 | 40.5 | 0.43 |
| Biostimulant formulate | |||||||||
| Control | 8.4 b | 6.9 b | 38.1 b | 29.0 b | 21.1 b | 10.8 b | 1.07 b | 38.7 b | 0.41 b |
| Legume-derived protein hydrolysate | 8.9 a | 7.6 a | 44.9 a | 35.1 a | 22.1 a | 12.4 a | 1.22 a | 43.3 a | 0.53 a |
| Tropical plant extract | 9.0 a | 7.6 a | 43.6 a | 34.4 a | 22.6 a | 13.3 a | 1.18 a | 44.6 a | 0.51 a |
| Source of Variation | Mineral Composition (g kg−1 dw) | |||||
|---|---|---|---|---|---|---|
| P | K | S | Ca | Mg | Na | |
| Farming system | ns | ns | * | ns | ns | ns |
| Biostimulant | * | * | * | ns | * | * |
| F × B | ns | ns | ns | ns | ns | ns |
| Farming system | ||||||
| Organic | 0.87 | 36.43 | 0.76 a | 5.51 | 1.44 | 0.31 |
| Conventional | 0.93 | 35.01 | 0.68 b | 6.02 | 1.52 | 0.29 |
| Biostimulant formulate | ||||||
| Control | 0.82 b | 33.83 b | 0.62 c | 5.60 | 1.30 b | 0.28 b |
| Legume-derived protein hydrolysate | 1.00 a | 36.66 a | 0.83 a | 5.72 | 1.59 a | 0.30 ab |
| Tropical plant extract | 0.87 ab | 36.68 a | 0.72 b | 5.92 | 1.56 a | 0.32 a |
| Source of Variation | Antioxidant Capacity | Lycopene | Total Phenols | Total Ascorbic Acid | |
|---|---|---|---|---|---|
| Lipophilic | Hydrophilic | ||||
| (mmol Trolox eq. 100g−1 dw) | (mmol Ascorbate eq. 100g−1 dw) | (mg 100g−1 fw) | (mg Gallic Acid eq. 100g−1 dw) | (mg 100g−1 fw) | |
| Farming system | * | ns | * | ns | * |
| Biostimulant | * | ns | * | * | * |
| F × B | ns | ns | ns | ns | ns |
| Farming system | |||||
| Organic | 8.1 a | 11.0 | 171.0 b | 1.9 | 23.9 a |
| Conventional | 7.7 b | 10.8 | 188.2 a | 1.9 | 18.5 b |
| Biostimulant formulate | |||||
| Control | 5.8 b | 10.7 | 150.2 b | 1.8 b | 14.5 c |
| Legume-derived protein hydrolysate | 9.1 a | 11.1 | 196.3 a | 2.0 a | 29.9 a |
| Tropical plant extract | 8.7 a | 10.9 | 192.0 a | 2.0 a | 19.2 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caruso, G.; De Pascale, S.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Bonini, P.; Colla, G.; Rouphael, Y. Yield and Nutritional Quality of Vesuvian Piennolo Tomato PDO as Affected by Farming System and Biostimulant Application. Agronomy 2019, 9, 505. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090505
Caruso G, De Pascale S, Cozzolino E, Cuciniello A, Cenvinzo V, Bonini P, Colla G, Rouphael Y. Yield and Nutritional Quality of Vesuvian Piennolo Tomato PDO as Affected by Farming System and Biostimulant Application. Agronomy. 2019; 9(9):505. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090505
Chicago/Turabian StyleCaruso, Gianluca, Stefania De Pascale, Eugenio Cozzolino, Antonio Cuciniello, Vincenzo Cenvinzo, Paolo Bonini, Giuseppe Colla, and Youssef Rouphael. 2019. "Yield and Nutritional Quality of Vesuvian Piennolo Tomato PDO as Affected by Farming System and Biostimulant Application" Agronomy 9, no. 9: 505. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090505