Assessment of Water Absorption Capacity and Cooking Time of Wild Under-Exploited Vigna Species towards their Domestication

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
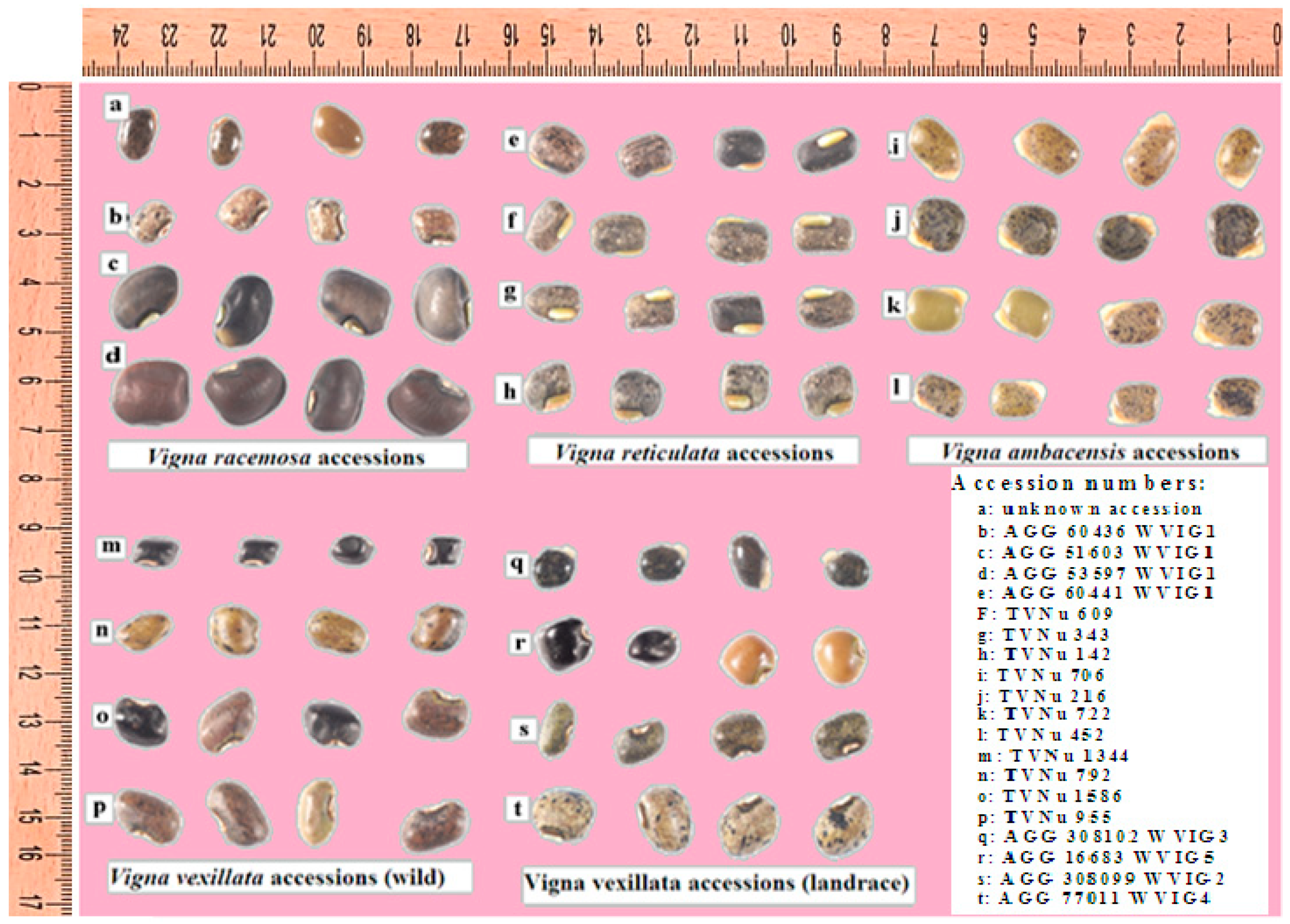
2.1. Sample Collection and Preparation
2.2. Sample Cultivation (Multiplication) Process: Experimental Design and Study Site.
2.3. Seed Soaking Process
2.4. Cooking Process on a Mattson Bean Cooker
2.5. Yield per Plant Data Collection and Evaluation
2.6. Data Analysis
3. Results
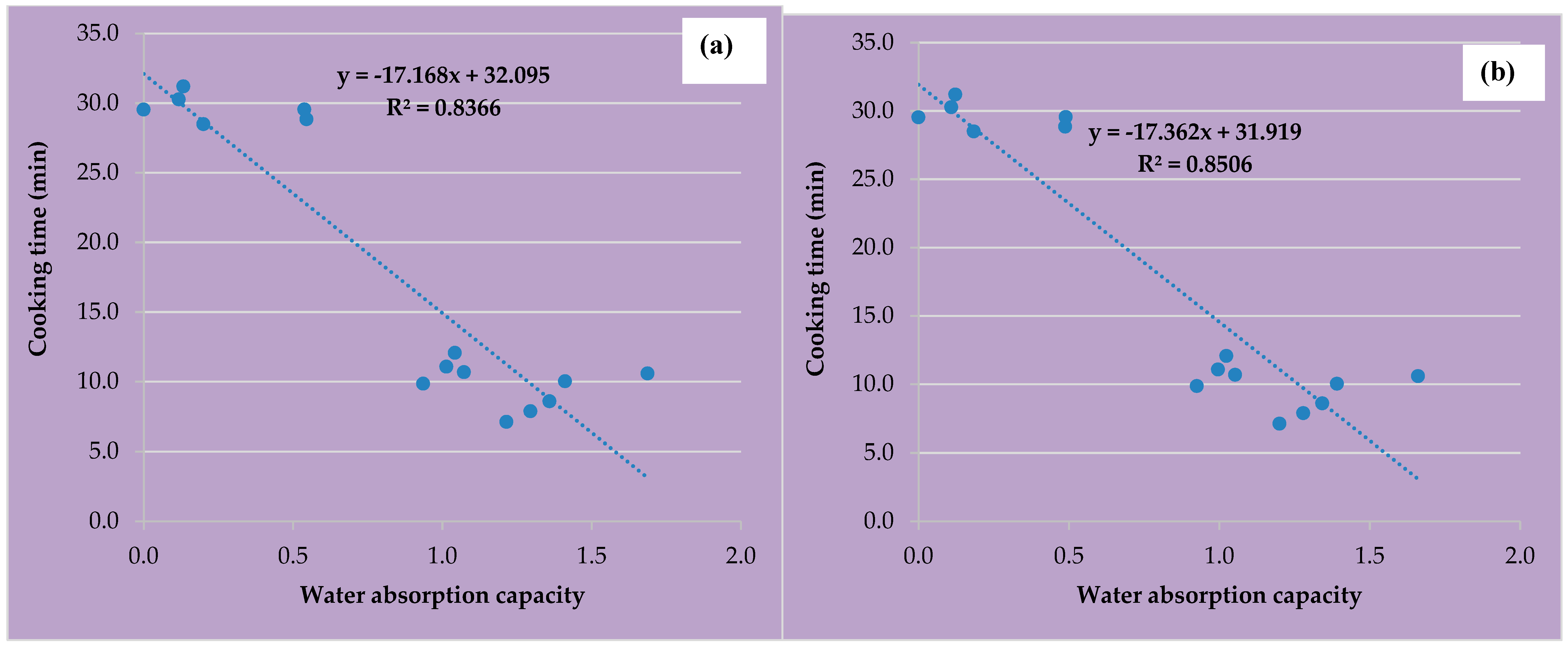
3.1. Cooking Time and Water Absorption Capacity of Domesticated Legumes
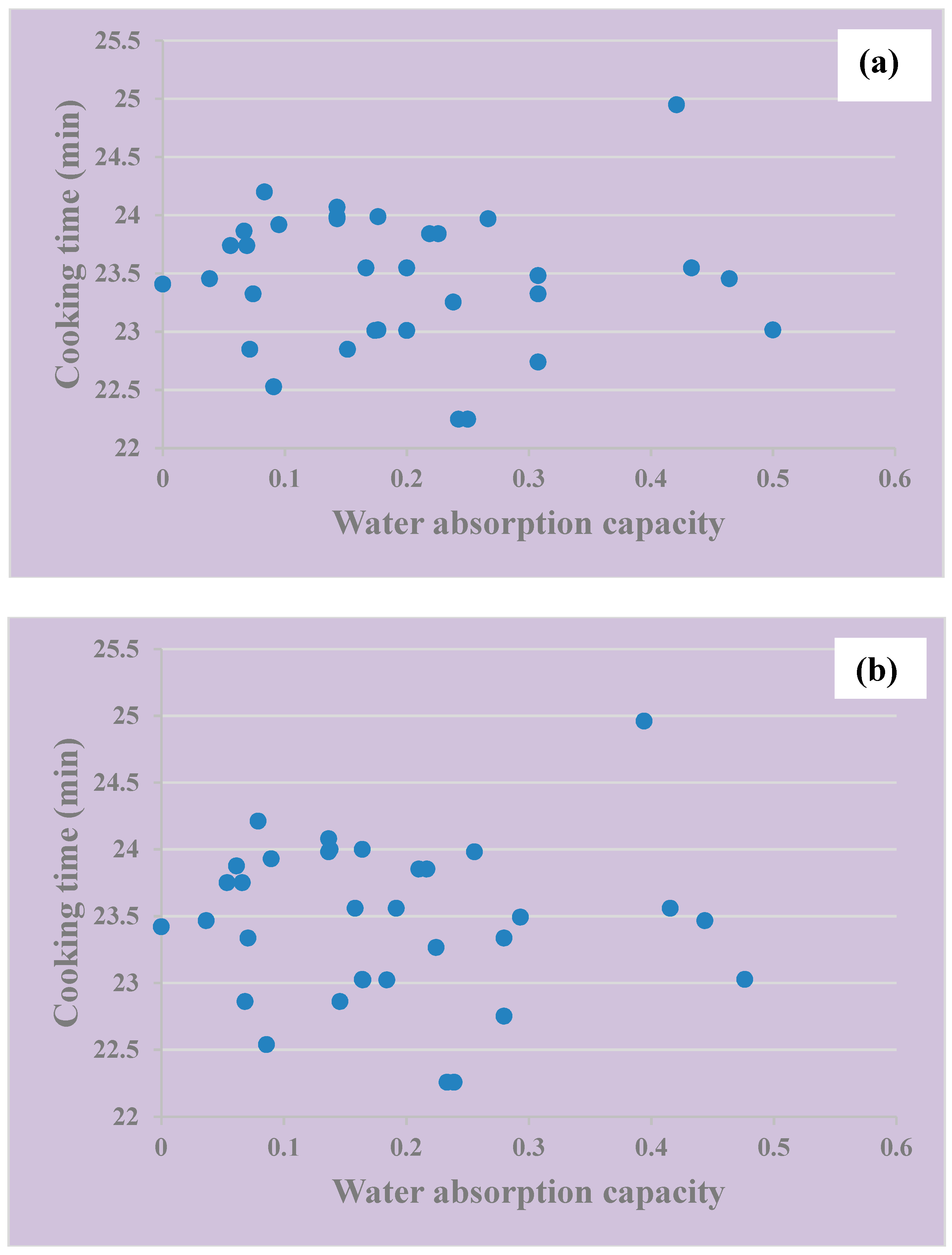
3.2. Cooking Time and Water Absorption Capacity of Vigna ambacensis Accessions
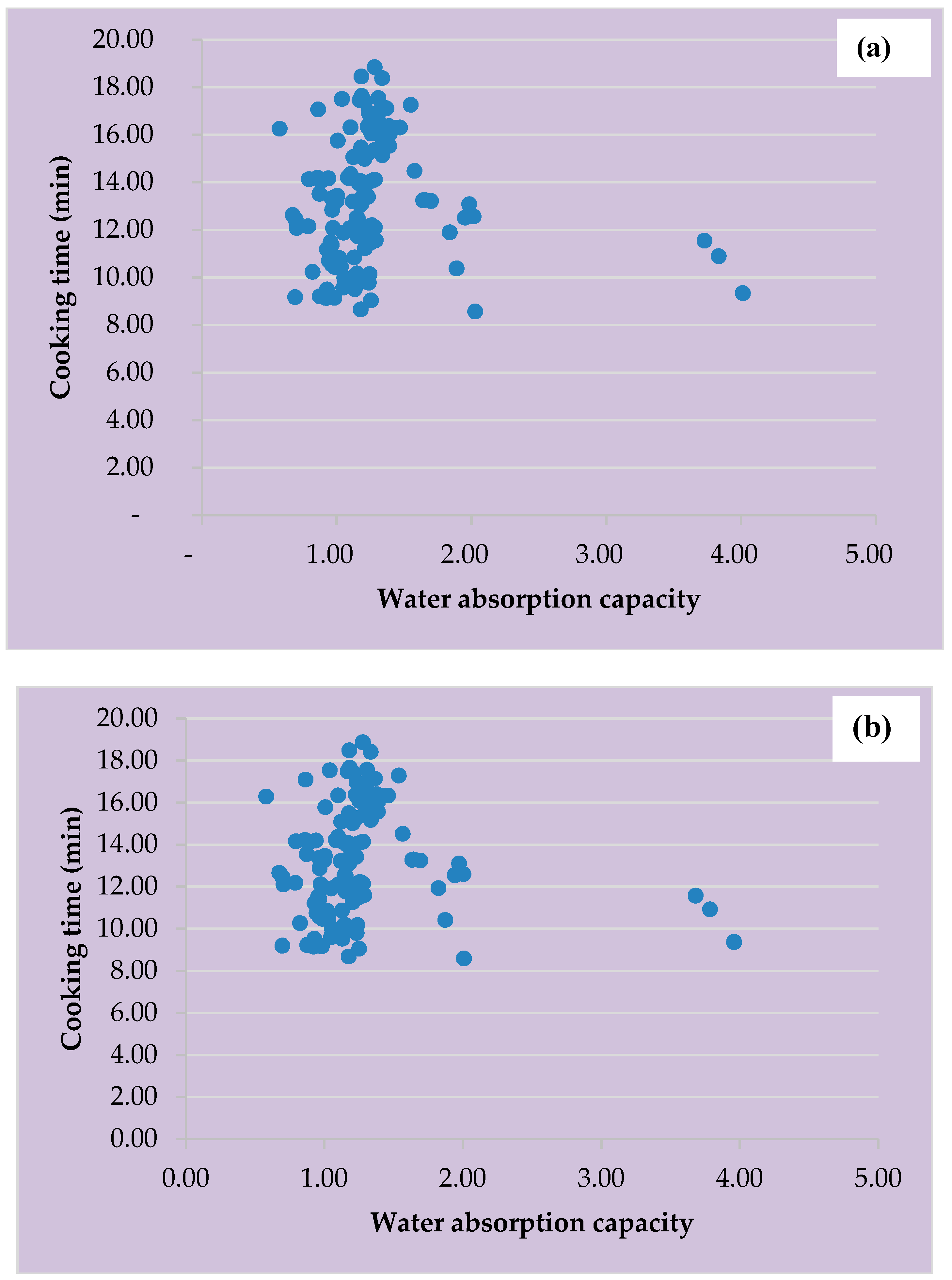
3.3. Cooking Time and Water Absorption Capacity of Vigna vexillata Accessions
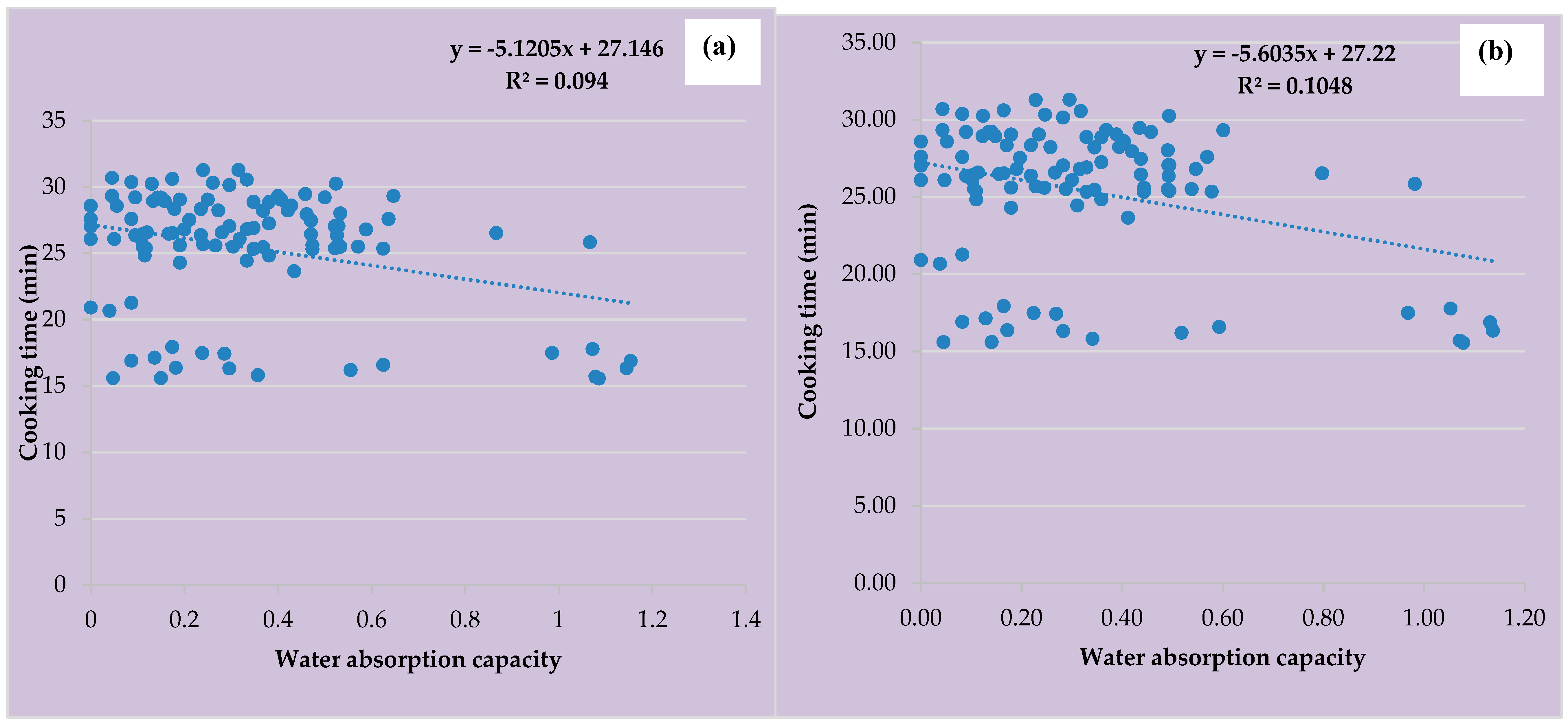
3.4. Cooking Time and Water Absorption capacity of Vigna reticulata Accessions
3.5. Cooking Time and Water Absorption of Vigna racemosa Accessions
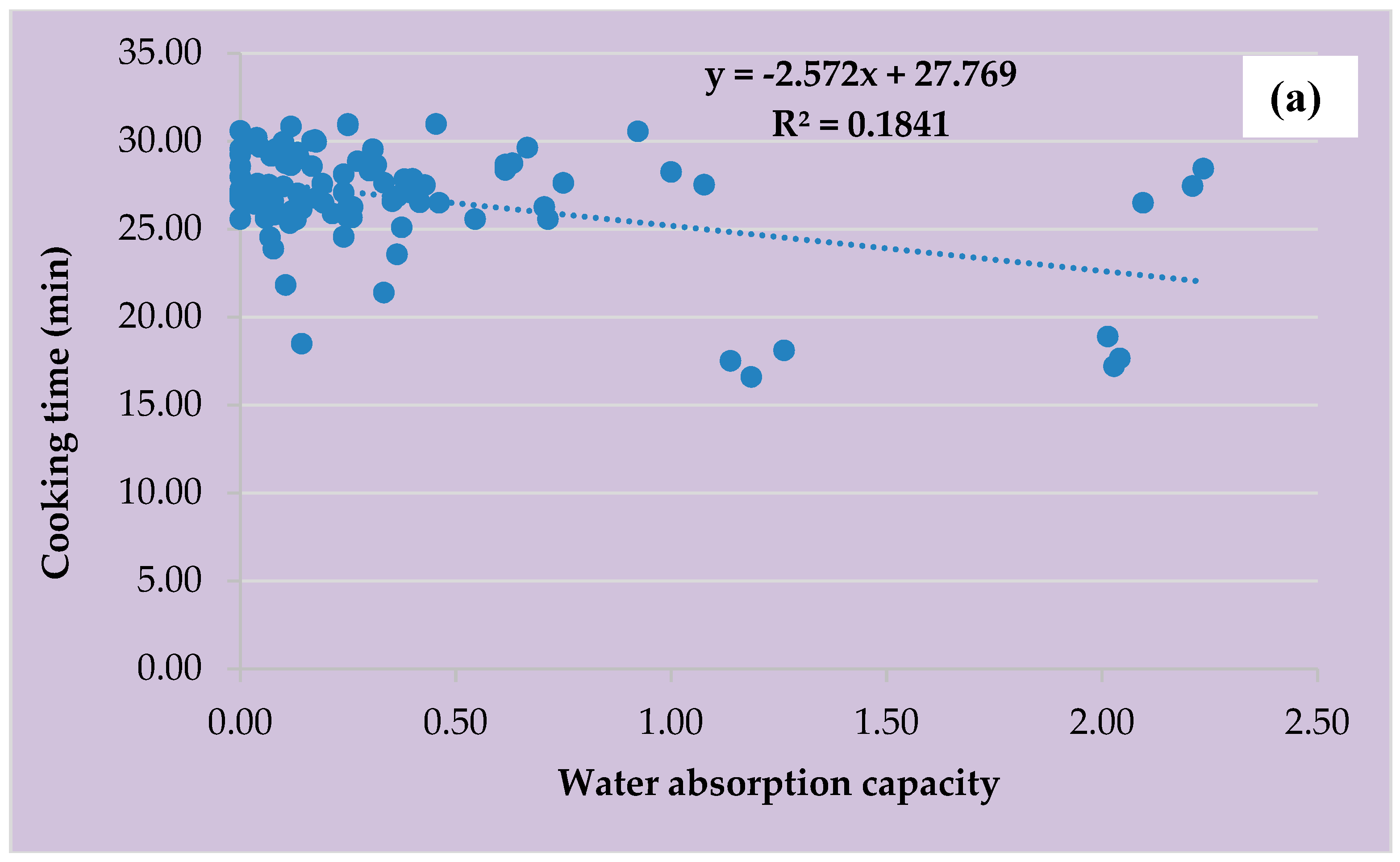
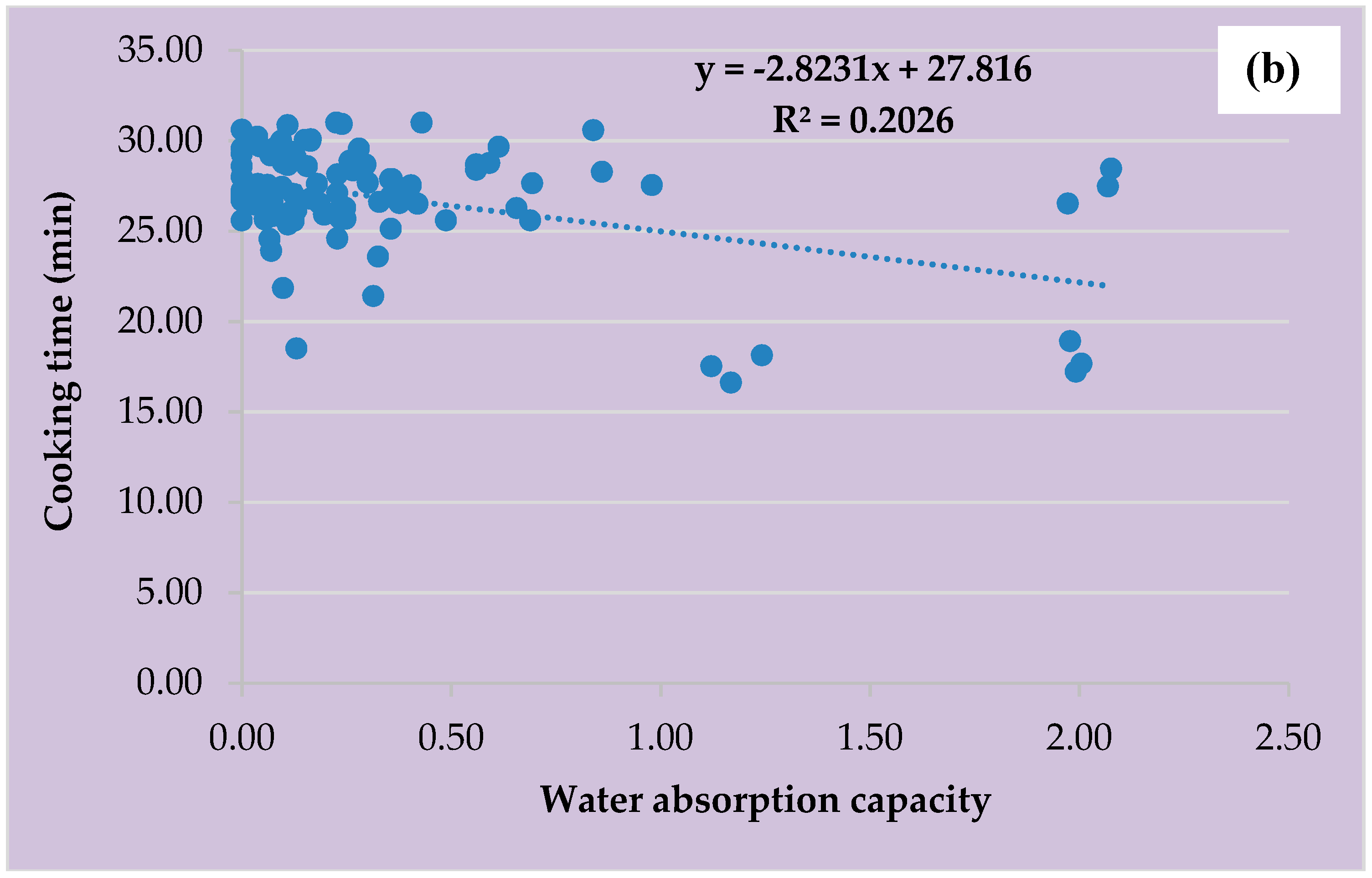
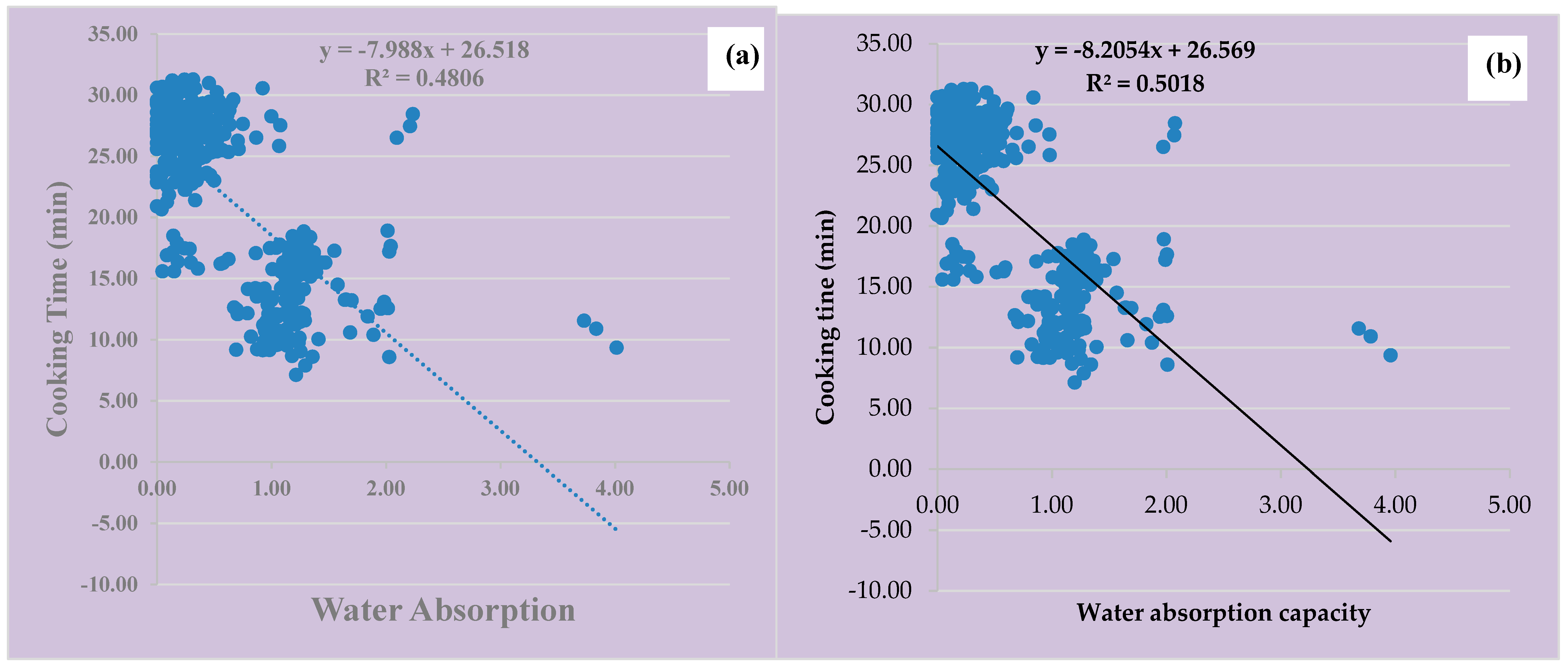
3.6. Water Absorption Capacity, Cooking Time, and Clustering Analysis of the Four Vigna species for Domestication and Crop Improvement
3.7. Descriptive Statistics and Yield Traits of the Wild Vigna Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bhat, R.; Karim, A.A. Exploring the nutritional potential of wild and underutilized legumes. Compr. Rev. Food Sci. Food Saf. 2009, 8, 305–331. [Google Scholar] [CrossRef]
- Harouna, D.V.; Venkataramana, P.B.; Ndakidemi, P.A.; Matemu, A.O. Under-exploited wild Vigna species potentials in human and animal nutrition: A review. Glob. Food Secur. 2018, 18, 1–11. [Google Scholar] [CrossRef]
- Ojiewo, C.; Monyo, E.; Desmae, H.; Boukar, O.; Mukankusi-Mugisha, C.; Thudi, M.; Pandey, M.K.; Saxena, R.K.; Gaur, P.M.; Chaturvedi, S.K.; et al. Genomics, genetics and breeding of tropical legumes for better livelihoods of smallholder farmers. Plant Breed. 2018, 138, 487–499. [Google Scholar] [CrossRef]
- Harouna, D.V.; Venkataramana, P.B.; Matemu, A.O.; Ndakidemi, P.A.; Harouna, D.V.; Venkataramana, P.B.; Matemu, A.O.; Ndakidemi, P.A. Wild Vigna legumes: farmers’ perceptions, preferences, and prospective uses for human exploitation. Agronomy 2019, 9, 284. [Google Scholar] [CrossRef]
- Wang, Q.J.; Mielby, L.A.; Junge, J.Y.; Bertelsen, A.S.; Kidmose, U.; Spence, C.; Byrne, D.V. The role of intrinsic and extrinsic sensory factors in sweetness perception of food and beverages: A review. Foods 2019, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Costell, E.; Tárrega, A.; Bayarri, S. Food acceptance: The role of consumer perception and attitudes. Chemosens. Percept. 2010, 3, 42–50. [Google Scholar] [CrossRef]
- Shafaei, S.M.; Masoumi, A.A.; Roshan, H. Analysis of water absorption of bean and chickpea during soaking using Peleg model. J. Saudi Soc. Agric. Sci. 2016, 15, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Turhan, M.; Sayer, S.; Gunasekaran, S. Application of Peleg model to study water absorption in bean and chickpea during soaking. J. Food Eng. 2002, 53, 153–159. [Google Scholar] [CrossRef]
- Hamid, S.; Muzaffar, S.; Wani, I.A.; Masoodi, F.A.; Bhat, M.M. Physical and cooking characteristics of two cowpea cultivars grown in temperate Indian climate. J. Saudi Soc. Agric. Sci. 2016, 15, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Difo, V.H.; Onyike, E.; Ameh, D.A.; Njoku, G.C.; Ndidi, U.S. Changes in nutrient and antinutrient composition of Vigna racemosa flour in open and controlled fermentation. J. Food Sci. Technol. 2015, 52, 6043–6048. [Google Scholar] [CrossRef]
- Siqueira, B.D.S.; Vianello, R.P.; Fernandes, K.F.; Bassinello, P.Z. Hardness of carioca beans (Phaseolus vulgaris L.) as affected by cooking methods. LWT Food Sci. Technol. 2013, 54, 13–17. [Google Scholar] [CrossRef]
- Garcia, E.H.; Pena-Valdivia, C.B.; Aguirre, J.R.R.; Muruaga, J.S.M. Morphological and agronomic traits of a wild population and an improved cultivar of Common Bean (Phaseolus vulgaris L.). Ann. Bot. 1997, 79, 207–213. [Google Scholar] [CrossRef]
- Gepts, P. Origins of plant agriculture and major crop plants. In Our fragile world, forerunner volumes to the Encyclopedia of Life-Supporting Systems; Tolba, M.K., Ed.; eolss publishers: Oxford, UK, 2001; pp. 629–637. [Google Scholar]
- Padulosi, S.; Thompson, J.; Rudebjer, P. Fighting Poverty, Hunger and Malnutrition with Neglected and Underutilized Species (NUS): Needs, Challenges and the Way Forward; Bioversity International: Rome, Italy, 2013; ISBN 9789290439417. [Google Scholar]
- Pratap, A.; Malviya, N.; Tomar, R.; Gupta, D.S.; Jitendra, K. Vigna. In Alien Gene Transfer in Crop Plants, Volume 2: Achievements and Impacts; Pratap, A., Kumar, J., Eds.; Springer Science+Business Media, LLC: Berlin/Heidelberg, Germany, 2014; pp. 163–189. ISBN 9781461495727. [Google Scholar]
- Tomooka, N.; Naito, K.; Kaga, A.; Sakai, H.; Isemura, T.; Ogiso-Tanaka, E.; Iseki, K.; Takahashi, Y. Evolution, domestication and neo-domestication of the genus Vigna. Plant Genet. Resour. Characterisation Util. 2014, 12, S168–S171. [Google Scholar] [CrossRef]
- Bisht, I.S.; Bhat, K.V.; Lakhanpaul, S.; Latha, M.; Jayan, P.K.; Biswas, B.K.; Singh, A.K. Diversity and genetic resources of wild Vigna species in India. Genet. Resour. Crop Evol. 2005, 52, 53–68. [Google Scholar] [CrossRef]
- Bassett, A.N.; Cichy, K.A.; Ambechew, D.; Bassett, A.N.; Cichy, K.A.; Ambechew, D. Cooking time and sensory analysis of a dry bean diversity panel. Annu. Rep. Bean Improv. Coop. 2017, 60, 154–156. [Google Scholar]
- IASRI Augmented Block Designs. Available online: http://www.iasri.res.in/design/Augmented Designs/home.htm (accessed on 18 January 2019).
- MCWatters, K.H.; CHinnan, M.S.; PHillips, R.D.; Beuchat, L.R.; Reid, L.B.; Mensa-Wilmot, Y.M. Functional, nutritional, mycological, and Akara- making properties of stored cowpea meal. J. Food Sci. 2002, 67, 2229–2234. [Google Scholar] [CrossRef]
- Wang, N.; Daun, J.K. Determination of cooking times of pulses using an automated Mattson cooker apparatus. J. Sci. Food Agric. 2005, 85, 1631–1635. [Google Scholar] [CrossRef]
- Proctor, J.R.; Watts, B.M. Development of a modified Mattson bean cooker procedure based on sensory panel cookability evaluation. Can. Inst. Food Sci. Technol. J. 2013, 20, 9–14. [Google Scholar] [CrossRef]
- Adewale, B.D.; Okonji, C.; Oyekanmi, A.A.; Akintobi, D.A.C.; Aremu, C.O. Genotypic variability and stability of some grain yield components of Cowpea. Afr. J. Agric. Res. 2010, 5, 874–880. [Google Scholar]
- Reguera, M.; Conesa, C.M.; Gil-Gómez, A.; Haros, C.M.; Pérez-Casas, M.Á.; Briones-Labarca, V.; Bolaños, L.; Bonilla, I.; Álvarez, R.; Pinto, K.; et al. The impact of different agroecological conditions on the nutritional composition of quinoa seeds. PeerJ 2018, 6, e4442. [Google Scholar] [CrossRef]
- Smykal, P.; Vernoud, V.; Blair, M.W.; Soukup, A.; Thompson, R.D. The role of the testa during development and in establishment of dormancy of the legume seed. Front. Plant Sci. 2014, 5, 1–19. [Google Scholar]
- Thompson, R.; Burstin, J.; Gallardo, K. Post-genomics studies of developmental processes in legume seeds: Figure 1. Plant Physiol. 2009, 151, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Zamindar, N.; Baghekhandan, M.S.; Nasirpour, A.; Sheikhzeinoddin, M. Effect of line, soaking and cooking time on water absorption, texture and splitting of red kidney beans. J. Food Sci. Technol. 2013, 50, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Jasrotia, R.S.; Yadav, P.K.; Iquebal, M.A.; Arora, V.; Angadi, U.B.; Tomar, R.S.; Rai, A.; Kumar, D. VigSatDB: Genome-wide microsatellite DNA marker database of three species of Vigna for germplasm characterization and improvement. Database 2019, 2019, baz055. [Google Scholar] [PubMed]
- Gore, P.G.; Tripathi, K.; Pratap, A.; Bhat, K.V.; Umdale, S.D.; Gupta, V.; Pandey, A. Delineating taxonomic identity of two closely related Vigna species of section Aconitifoliae: V. trilobata (L.) Verdc. and V. stipulacea (Lam.) Kuntz in India. Genet. Resour. Crop Evol. 2019, 66, 1155–1165. [Google Scholar] [CrossRef]
- Tomooka, N.; Kaga, A.; Isemura, T.; Vaughan, D. Vigna. In Wild Crop Relatives: Genomic and Breeding Resources: Legume Crops and Forages; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany; London, UK, 2011; pp. 1–321. ISBN 9783642143878. [Google Scholar]
- Karuniawan, A.; Iswandi, A.; Kale, P.R.; Heinzemann, J.; Grüneberg, W.J. Vigna vexillata (L.) A. Rich. cultivated as a root crop in Bali and Timor. Genet. Resour. Crop Evol. 2006, 53, 213–217. [Google Scholar] [CrossRef]
- Dachapak, S.; Somta, P.; Poonchaivilaisak, S.; Yimram, T.; Srinives, P. Genetic diversity and structure of the zombi pea (Vigna vexillata (L.) A. Rich) gene pool based on SSR marker analysis. Genetica 2017, 145, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, M.M.; Jaeger De Carvalho, L.M.; Nutti, M.R.; Luiz, J.; De Carvalho, V.; Neto, A.R.H.; Gomes Ribeiro, E.M. Water Absorption, Hard Shell and Cooking Time of Common Beans (Phaseolus vulgaris L.). Afr. J. Food Sci. Technol. 2010, 1, 13–20. [Google Scholar]
- Hatcher, D.W.; Kruger, J.E.; Anderson, M.J. Influence of water absorption on the processing and quality of oriental noodles. Cereal Chem. 1999, 76, 566–572. [Google Scholar] [CrossRef]
- Schlautman, B.; Barriball, S.; Ciotir, C.; Herron, S.; Miller, A.J. Perennial grain legume domestication Phase I: Criteria for candidate species selection. Sustainability 2018, 10, 730. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vigna Species | Genebank/Number of Accession | Total | ||
|---|---|---|---|---|
| GRC, IITA Ibadan, Nigeria | AGG Horsham, Victoria | Self-Collected | ||
| Vigna racemosa | - | 4 | - | 4 |
| Vigna reticulata | 30 | 1 | - | 31 |
| Vigna vexillata | 29 | 6 | - | 35 |
| Vigna ambacensis | 11 | 0 | - | 11 |
| Unknown V. racemosa Accession (Nigeria) | - | - | 1 | 1 |
| Unknown V. reticulata Accession (Nigeria) | - | - | 1 | 1 |
| Unknown Vigna (Tanzania) | - | - | 1 | 1 |
| Total | 70 | 11 | 3 | 84 |
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Checks | Site A | Site B | Site A | Site B | ||||||
| Landrace of Vigna vexillata | 1.33 ± 0.11 a | 1.32 ± 0.13 a | 10.24 ± 0.15 a | 10.26 ± 0.15 a | ||||||
| Cowpea (Vigna unguiculata) | 1.27 ± 0.08 a | 1.27 ± 0.08 a | 16.29 ± 0.15 c | 16.31 ± 0.15 c | ||||||
| Rice Bean (Vigna umbellata) | 1.16 ± 0.06 a | 1.16 ± 0.06 a | 13.20 ± 0.12 b | 13.23 ± 0.12 b | ||||||
| Analysis of Variance (ANOVA) | ||||||||||
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Model | 5 | 1.263 | 0.253 | 1.134 | 0.343 | 5 | 1582.515 | 316.503 | 356.710 | <0.0001 |
| Error | 258 | 57.475 | 0.223 | 258 | 228.919 | 0.887 | ||||
| Corrected Total | 263 | 58.738 | 263 | 1811.434 | ||||||
| Type III Sum of Squares Analysis | ||||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Location (Site) Effect | 1 | 0.001 | 0.001 | 0.004 | 0.950 | 1 | 0.044 | 0.044 | 0.050 | 0.823 |
| Species Effect | 2 | 1.262 | 0.631 | 2.833 | 0.061 | 2 | 1582.470 | 791.235 | 891.749 | <0.0001 |
| Location X Species | 2 | 0.000 | 0.000 | 0.000 | 1.000 | 2 | 0.000 | 0.000 | 0.000 | 1.000 |
| Location × Species/Tukey (HSD)/Analysis of the Differences between the Categories with a Confidence Interval of 95% (Water Absorption Capacity) | |||||
|---|---|---|---|---|---|
| Contrast | Difference | Standardized Difference | Critical value | Pr > Diff | Significant |
| Location-Site A × Species-Check 1 vs. Location-Site B × Species-Check 3 | 0.173 | 1.675 | 2.871 | 0.550 | No |
| Location-Site A × Species-Check 1 vs. Location-Site A × Species-Check 3 | 0.172 | 1.661 | 2.871 | 0.559 | No |
| Location-Site A × Species-Check 1 vs. Location-Site B × Species-Check 2 | 0.063 | 0.620 | 2.871 | 0.990 | No |
| Location-Site A × Species-Check 1 vs. Location-Site A × Species-Check 2 | 0.059 | 0.584 | 2.871 | 0.992 | No |
| Location-Site A × Species-Check 1 vs. Location-Site B × Species-Check 1 | 0.006 | 0.054 | 2.871 | 1.000 | No |
| Location-Site B × Species-Check 1 vs. Location-Site B × Species-Check 3 | 0.167 | 1.619 | 2.871 | 0.587 | No |
| Location-Site B × Species-Check 1 vs. Location-Site A × Species-Check 3 | 0.166 | 1.605 | 2.871 | 0.596 | No |
| Location-Site B × Species-Check 1 vs. Location-Site B × Species-Check 2 | 0.057 | 0.563 | 2.871 | 0.993 | No |
| Location-Site B × Species-Check 1 vs. Location-Site A × Species-Check 2 | 0.054 | 0.527 | 2.871 | 0.995 | No |
| Location-Site A × Species-Check 2 vs. Location-Site B × Species-Check 3 | 0.114 | 1.160 | 2.871 | 0.855 | No |
| Location-Site A × Species-Check 2 vs. Location-Site A × Species-Check 3 | 0.112 | 1.145 | 2.871 | 0.862 | No |
| Location-Site A × Species-Check 2 vs. Location-Site B × Species-Check 2 | 0.004 | 0.038 | 2.871 | 1.000 | No |
| Location-Site B × Species-Check 2 vs. Location-Site B × Species-Check 3 | 0.110 | 1.122 | 2.871 | 0.872 | No |
| Location-Site B × Species-Check 2 vs. Location-Site A × Species-Check 3 | 0.108 | 1.107 | 2.871 | 0.878 | No |
| Location-Site A × Species-Check 3 vs. Location-Site B × Species-Check 3 | 0.001 | 0.014 | 2.871 | 1.000 | No |
| Tukey’s d critical value | 4.061 | ||||
| Location × Species/Tukey (HSD)/Analysis of the Differences between the Categories with a Confidence Interval of 95% (Cooking Time) | |||||
|---|---|---|---|---|---|
| Contrast | Difference | Standardized Difference | Critical value | Pr > Diff | Significant |
| Location-Site B × Species-Check 2 vs. Location-Site A × Species-Check 1 | 6.074 | 29.913 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 2 vs. Location-Site B × Species-Check 1 | 6.048 | 29.785 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 2 vs. Location-Site A × Species-Check 3 | 3.109 | 15.908 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 2 vs. Location-Site B × Species-Check 3 | 3.083 | 15.775 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 2 vs. Location-Site A × Species-Check 2 | 0.026 | 0.135 | 2.871 | 1.000 | No |
| Location-Site A × Species-Check 2 vs. Location-Site A × Species-Check 1 | 6.048 | 29.785 | 2.871 | <0.0001 | Yes |
| Location-Site A × Species-Check 2 vs. Location-Site B × Species-Check 1 | 6.022 | 29.657 | 2.871 | <0.0001 | Yes |
| Location-Site A × Species-Check 2 vs. Location-Site A × Species-Check 3 | 3.083 | 15.775 | 2.871 | <0.0001 | Yes |
| Location-Site A × Species-Check 2 vs. Location-Site B × Species-Check 3 | 3.057 | 15.642 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 3 vs. Location-Site A × Species-Check 1 | 2.991 | 14.514 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 3 vs. Location-Site B × Species-Check 1 | 2.965 | 14.388 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 3 vs. Location-Site A × Species-Check 3 | 0.026 | 0.131 | 2.871 | 1.000 | No |
| Location-Site A × Species-Check 3 vs. Location-Site A × Species-Check 1 | 2.965 | 14.388 | 2.871 | <0.0001 | Yes |
| Location-Site A × Species-Check 3 vs. Location-Site B × Species-Check 1 | 2.939 | 14.262 | 2.871 | <0.0001 | Yes |
| Location-Site B × Species-Check 1 vs. Location-Site A × Species-Check 1 | 0.026 | 0.122 | 2.871 | 1.000 | No |
| Tukey’s d critical value | 4.061 | ||||
| Species/Accession Number | Water Absorption Capacity | Cooking Time (min) | ||||||||
| Site A | Site B | Site A | Site B | |||||||
| Landrace of Vigna vexillata | 1.33 ± 0.11 a | 1.32 ± 0.13 a | 10.24 ± 0.15 a | 10.26 ± 0.15 a | ||||||
| Cowpea (Vigna unguiculata) | 1.27 ± 0.08 a | 1.27 ± 0.08 a | 16.29 ± 0.15 b | 16.31 ± 0.15 b | ||||||
| Rice bean (Vigna umbellata) | 1.16 ± 0.06 a | 1.16 ± 0.06 a | 13.20 ± 0.12 c | 13.23 ± 0.12 c | ||||||
| TVNu1699 | 0.14 ± 0.01 c | 0.13 ± 0.01 c | 24.26 ± 0.07 d | 23.87 ± 0.10 d | ||||||
| TVNu342 | 0.47 ± 0.01 a,b | 0.45 ± 0.01 a,b | 23.34 ± 0.16 d | 23.35 ± 0.18 d | ||||||
| TVNu877 | 0.22 ± 0.01c | 0.21 ± 0.01 c | 24.10 ± 0.19 d | 23.71 ± 0.22 d | ||||||
| TVNu223 | 0.21 ± 0.01 c | 0.21 ± 0.02 c | 23.35 ± 0.55 d | 23.36 ± 0.50 d | ||||||
| TVNu720 | 0.22 ± 0.01 c | 0.20 ± 0.01 c | 23.02 ± 0.50 d | 23.03 ± 0.45 d | ||||||
| TVNu219 | 0.28 ± 0.02 b,c | 0.26 ± 0.01 b,c | 24.06 ± 0.49 d | 24.08 ± 0.50 d | ||||||
| TVNu1840 | 0.11 ± 0.01 c | 0.10 ± 0.01 c | 23.36 ± 0.21 d | 23.37 ± 0.30 d | ||||||
| TVNu1804 | 0.09 ± 0.01 c | 0.08 ± 0.01 c | 23.55 ± 0.52 d | 23.56 ± 0.50 d | ||||||
| TVNu1792 | 0.23 ± 0.01 c | 0.09 ± 0.01 c | 23.28 ± 0.22 d | 23.30 ± 0.30 d | ||||||
| TVNu1644 | 0.09 ± 0.01 c | 0.21 ± 0.01 c | 23.12 ± 0.10 d | 23.13 ± 0.15 d | ||||||
| TVNu1185 | 0.12 ± 0.01 c | 0.11 ± 0.01 c | 23.34 ± 0.33 d | 23.35 ± 0.30 d | ||||||
| Analysis of Variance (ANOVA) | ||||||||||
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Model | 27 | 60.707 | 2.248 | 11.756 | <0.0001 | 27 | 6864.480 | 254.240 | 313.317 | <0.0001 |
| Error | 302 | 57.761 | 0.191 | 302 | 245.057 | 0.811 | ||||
| Corrected Total | 329 | 118.469 | 329 | 7109.537 | ||||||
| Type III Sum of Squares Analysis | ||||||||||
| Source | DF | Sum of squares | Mean squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Location (Site) | 1 | 0.002 | 0.002 | 0.011 | 0.916 | 1 | 0.007 | 0.007 | 0.008 | 0.929 |
| Genotype (Accessions) | 13 | 60.704 | 4.670 | 24.414 | <0.0001 | 13 | 6864.433 | 528.033 | 650.730 | <0.0001 |
| Location × Genotype | 13 | 0.001 | 0.000 | 0.001 | 1.000 | 13 | 0.002 | 0.000 | 0.000 | 1.000 |
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Species/Accession Number | Site A | Site B | Site A | Site B | ||||||
| Landrace of Vigna vexillata | 1.33 ± 0.11 a | 1.32 ± 0.13 a | 10.24 ± 0.15 n | 10.26 ± 0.15 n | ||||||
| Cowpea (Vigna unguiculata) | 1.27 ± 0.08 a,b,c | 1.27 ± 0.08 a,b,c | 16.29 ± 0.15 l | 16.31 ± 0.15 l | ||||||
| Rice Bean (Vigna umbellata) | 1.16 ± 0.06 a,b,c | 1.16 ± 0.06 a,b,c | 13.20 ± 0.12 m | 13.23 ± 0.12 m | ||||||
| TVNu781 | 1.10 ± 0.02 abcd | 1.10 ± 0.01 abcd | 31.04 ± 0.33 a,b | 31.06 ± 0.31 a,b | ||||||
| TVNu837 | 1.07 ± 0.01 abcd | 1.05 ± 0.01 abcd | 29.34 ± 0.32 a,b,c,d,e,f,g | 29.35 ± 0.01 a,b,c,d,e,f,g | ||||||
| TVNu1582 | 0.73 ± 0.01 abcd | 0.67 ± 0.02 abcd | 16.25 ± 0.24 l | 16.26 ± 0.30 l | ||||||
| TVNu1358 | 0.57 ± 0.01 abcd | 0.53 ± 0.01 abcd | 17.37 ± 0.26 l | 17.38 ± 0.28 l | ||||||
| AGG308107WVIG2 | 0.43 ± 0.01 abcd | 0.41 ± 0.01 abcd | 26.32 ± 0.49 f,g,h,i,j | 26.33 ± 0.51 f,g,h,i,j | ||||||
| TVNu1593 | 0.42 ± 0.01 abcd | 0.38 ± 0.01 bcd | 16.22 ± 0.23 l | 16.24 ± 0.20 l | ||||||
| TVNu1591 | 0.41 ± 0.01 abcd | 0.38 ± 0.01 bcd | 26.38 ± 0.40 f,g,h,i,j | 26.39 ± 0.43 f,g,h,i,j | ||||||
| TVNu120 | 0.40 ± 0.01 abcd | 0.38 ± 0.01 bcd | 31.10 ± 0.31 a | 30.71 ± 0.34 a | ||||||
| TVNu333 | 0.40 ± 0.02 abcd | 0.37 ± 0.02 bcd | 26.28 ± 0.40 f,g,h,i,j | 26.30 ± 0.35 f,g,h,i,j | ||||||
| TVNu1546 | 0.39 ± 0.02 bcd | 0.37 ± 0.01 bcd | 29.07 ± 0.13 a,b,c,d,e,f | 29.08 ± 0.15 a,b,c,d,e,f | ||||||
| AGG308101WVIG1 | 0.37 ± 0.01 bcd | 0.34 ± 0.01bcd | 29.36 ± 0.50 a,b,c,d,e | 29.37 ± 0.47 a,b,c,d,e | ||||||
| TVNu1701 | 0.35 ± 0.01 bcd | 0.33 ± 0.01 cd | 24.59 ± 0.50 j | 24.60 ± 0.57 j | ||||||
| AGG308096 WVIG2 | 0.34 ± 0.01 cd | 0.32 ± 0.01cd | 26.47 ± 0.59 f,g,h,i,j | 26.49 ± 0.60 f,g,h,i,j | ||||||
| TVNu1629 | 0.33 ± 0.01 cd | 0.32 ± 0.02 cd | 28.19 ± 1.15 b,c,d,e,f,g,h,i | 28.20 ± 1.20 b,c,d,e,f,g,h,i | ||||||
| TVNu293 | 0.33 ± 0.01 cd | 0.31 ± 0.01 cd | 26.32 ± 0.28 f,g,h,i,j | 26.33 ± 0.30 f,g,h,i,j | ||||||
| TVNu832 | 0.32 ± 0.01 cd | 0.30 ± 0.01 cd | 25.46 ± 0.36 h,i,j | 25.47 ± 0.36 h,i,j | ||||||
| TVNu1796 | 0.32 ± 0.01 cd | 0.30 ± 0.01 cd | 27.38 ± 0.48 c,d,e,f,g,h,i,j | 27.39 ± 0.50 c,d,e,f,g,h,i,j | ||||||
| TVNu1529 | 0.32 ± 0.01 cd | 0.30 ± 0.01 cd | 27.29 ± 0.64 c,d,e,f,g,h,i,j | 27.30 ± 0.64 c,d,e,f,g,h,i,j | ||||||
| TVNu1628 | 0.30 ± 0.01 cd | 0.28 ± 0.01cd | 26.30 ± 0.36 f,g,h,i,j | 26.31 ± 0.33 f,g,h,i,j | ||||||
| TVNu1344 | 0.29 ± 0.01 cd | 0.28 ± 0.01 cd | 30.03 ± 0.44 a,b,c | 30.04 ± 0.44 a,b,c | ||||||
| TVNu1632 | 0.29 ± 0.01 cd | 0.28 ± 0.01 cd | 29.41 ± 0.52 a,b,c,d | 28.25 ± 0.50 a,b,c,d | ||||||
| TVNu1370 | 0.28± 0.01 cd | 0.26 ± 0.02 cd | 28.23 ± 0.39l | 29.43 ± 0.40 l | ||||||
| TVNu1360 | 0.28± 0.01 cd | 0.25 ± 0.01 cd | 27.05 ± 0.71 d,e,f,g,h,i,j | 27.60 ± 0.72 d,e,f,g,h,i,j | ||||||
| TVNu1624 | 0.25± 0.01 cd | 0.23 ± 0.01 d | 26.28 ± 0.46 f,g,h,i,j | 26.29 ± 0.46 f,g,h,i,j | ||||||
| TVNu1621 | 0.25 ± 0.01 cd | 0.23 ± 0.01 d | 17.24 ± 0.47 l | 17.26 ± 0.48 l | ||||||
| AGG62154WVIG_1 | 0.20 ± 0.01 d | 0.19 ± 0.01 d | 21.33 ± 0.17 k | 21.34 ± 0.17 k | ||||||
| TVNu1092 | 0.19 ± 0.01 d | 0.18 ± 0.01 d | 29.02 ± 0.23 a,b,c,d,e,f,g | 29.04 ± 0.55 a,b,c,d,e,f,g | ||||||
| TVNu479 | 0.18 ± 0.01 d | 0.17 ± 0.01 d | 26.50 ± 0.56 e,f,g,h,i,j | 26.52 ± 0.20 e,f,g,h,i,j | ||||||
| AGG308097WVIG 1 | 0.17 ± 0.01 d | 0.16 ± 0.01 d | 28.56 ± 0.50 a,b,c,d,e,f,g | 28.58 ± 0.50 a,b,c,d,e,f,g | ||||||
| TVNu178 | 0.17 ± 0.01 d | 0.16 ± 0.01 d | 17.03 ± 0.54 l | 16.64 ± 0.01 l | ||||||
| TVNu955 | 0.11 ± 0.01 d | 0.11 ± 0.01 d | 25.28 ± 0.47 i,j | 25.29 ± 0.47 i,j | ||||||
| TVNu1378 | 0.11 ± 0.01 d | 0.10 ± 0.00 d | 16.29 ± 0.45 l | 16.31 ± 0.47 l | ||||||
| TVNu1586 | 0.06 ± 0.00 d | 0.05 ± 0.01 d | 28.39 ± 0.29 a,b,c,d,e,f,g | 28.41 ± 0.30 a,b,c,d,e,f,g | ||||||
| TVNu381 | 0.04 ± 0.00 d | 0.042 ± 0.00 d | 25.41 ± 0.63 h,i,j | 25.42 ± 0.64 h,i,j | ||||||
| AGG308099WVIG2 | 0.042 ± 0.01 d | 0.04 ± 0.01 d | 26.16 ± 0.48 h,i,j | 26.17 ± 0.50 h,i,j | ||||||
| Analysis of Variance (ANOVA) | ||||||||||
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Model | 75 | 111.003 | 1.480 | 9.649 | <0.0001 | 75 | 22,437.582 | 299.168 | 368.513 | <0.0001 |
| Error | 398 | 61.050 | 0.153 | 398 | 323.106 | 0.812 | ||||
| Corrected Total | 473 | 172.052 | 473 | 22,760.688 | ||||||
| Type III Sum of Squares Analysis | ||||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Location (Site) | 1 | 0.018 | 0.018 | 0.117 | 0.732 | 1 | 0.012 | 0.012 | 0.015 | 0.903 |
| Genotype (Accessions) | 37 | 110.978 | 2.999 | 19.554 | <0.0001 | 37 | 22,437.529 | 606.420 | 746.983 | <0.0001 |
| Location × Genotype | 37 | 0.013 | 0.000 | 0.002 | 1.000 | 37 | 0.005 | 0.000 | 0.000 | 1.000 |
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Species/Accession Number | Site A | Site B | Site A | Site B | ||||||
| Landrace of Vigna vexillata | 1.33 ± 0.11 a,b,c | 1.32 ± 0.13 a,b,c | 10.24 ± 0.15 h | 10.26 ± 0.15 h | ||||||
| Cowpea (Vigna unguiculata) | 1.27 ± 0.08 a,b,c,d | 1.27 ± 0.08 a,b,c,d | 16.29 ± 0.15 f | 16.31 ± 0.15 f | ||||||
| Rice Bean (Vigna umbellata) | 1.16 ± 0.06 a,b,c,d | 1.16 ± 0.06 a,b,c,d | 13.20 ± 0.12 g | 13.23 ± 0.12 g | ||||||
| TVNu324 | 0.49 ± 0.02 c,d | 0.47 ± 0.02 c,d | 26.51 ± 0.47 a,b,c,d | 26.53 ± 0.48 a,b,c,d | ||||||
| TVNu325 | 2.03 ± 0.02 a,b | 1.99 ± 0.01 a,b | 17.92 ± 0.51 f | 17.93 ± 0.52 f | ||||||
| Unknown _Vigna reticulata | 1.20 ± 0.02 a,b,c,d | 1.18 ± 0.02 a,b,c,d | 17.41 ± 0.44 f | 17.42 ± 0.45 f | ||||||
| TVNu343 | 0.19 ± 0.01 c,d | 0.18 ± 0.01c,d | 30.25 ± 0.41 a | 30.26 ± 0.42 a | ||||||
| TVNu767 | 0.12 ± 0.01 c,d | 0.11 ± 0.01 c,d | 29.18 ± 0.99 a,b,c,d | 29.20 ± 1.00 a,b,c,d | ||||||
| TVNu1520 | 2.18 ± 0.03 a | 2.04 ± 0.03 a | 27.46 ± 0.91 a,b,c,d | 27.48 ± 0.92 a,b,c,d | ||||||
| TVNu349 | 0.31 ± 0.02 c,d | 0.29 ± 0.01 c,d | 29.14 ± 0.74 a,b,c,d | 28.76 ± 0.75 a,b,c,d | ||||||
| TVNu379 | 0.77 ± 0.01 c,d | 0.71 ± 0.02 c,d | 29.38 ± 0.46 a,b,c,d | 28.99 ± 0.44 a,b,c,d | ||||||
| TVNu524 | 0.17 ± 0.01 c,d | 0.17 ± 0.01 c,d | 25.57 ± 0.57 c,d,e | 25.58 ± 0.58 c,d,e | ||||||
| TVNu1698 | 0.12 ± 0.01 c,d | 0.11 ± 0.01 c,d | 26.34 ± 0.56 b,c,d,e | 26.36 ± 0.57 b,c,d,e | ||||||
| TVNu1191 | 0.22 ± 0.01 c,d | 0.21 ± 0.01 c,d | 25.38 ± 1.00 d,e | 25.39 ± 0.99 d,e | ||||||
| TVNu1394 | 0.82 ± 0.02 b, c,d | 0.75 ± 0.01 b, c,d | 28.21 ± 0.99 a,b,c,d | 27.82 ± 0.97 a,b,c,d | ||||||
| TVNu-224 | 0.19 ± 0.01 c,d | 0.18 ± 0.01 c,d | 25.55 ± 0.51 c,d,e | 25.57 ± 0.52 c,d,e | ||||||
| TVNu739 | 0.15 ± 0.01 c,d | 0.14 ± 0.01 c,d | 28.50 ± 0.46 a,b,c,d | 28.52 ± 0.47 a,b,c,d | ||||||
| TVNu56 | 0.24 ± 0.02 c,d | 0.22 ± 0.02 c,d | 27.01 ± 2.73 a,b,c,d | 26.62 ± 2.70 a,b,c,d | ||||||
| TVNu1405 | 0.29 ± 0.02 c,d | 0.26 ± 0.02 c,d | 30.03 ± 0.64 a,b | 29.64 ± 0.62 a,b | ||||||
| TVNu607 | 0.08 ± 0.01 d | 0.08 ± 0.01 d | 27.33 ± 0.49 a,b,c,d | 26.38 ± 0.47 a,b,c,d | ||||||
| TVNu916 | 0.12 ± 0.01 c,d | 0.11 ± 0.01 c,d | 26.37 ± 0.52 b,c,d,e | 27.84 ± 0.55 b,c,d,e | ||||||
| AGG17856WVIG 1 | 0.16 ± 0.01 c,d | 0.15 ± 0.01 c,d | 28.23 ± 1.00 a,b,c,d | 27.35 ± 0.97 a,b,c,d | ||||||
| TVNu1790 | 0.32 ± 0.02 c,d | 0.29 ± 0.02 c,d | 28.43 ± 0.47 a,b,c,d | 28.44 ± 0.47 a,b,c,d | ||||||
| TVNu491 | 0.15 ± 0.01 c,d | 0.14 ± 0.01 c,d | 28.44 ± 0.93 a,b,c,d | 28.45 ± 0.92 a,b,c,d | ||||||
| TVNu1808 | 0.16 ± 0.01 c,d | 0.15 ± 0.01 c,d | 29.36 ± 0.42 a,b,c | 29.38 ± 0.43 a,b,c | ||||||
| TVNu738 | 0.12 ± 0.01 c,d | 0.12 ± 0.01 c,d | 26.42 ± 0.39 a,b,c,d | 26.43 ± 0.40 a,b,c,d | ||||||
| TVNu1779 | 0.19 ± 0.02 c,d | 0.17 ± 0.01 c,d | 26.12 ± 2.04 c,d,e | 25.74 ± 2.01 c,d,e | ||||||
| TVNu605 | 0.42 ± 0.02 c,d | 0.36 ± 0.02 c,d | 29.16 ± 0.51 a,b,c,d | 29.18 ± 0.51 a,b,c,d | ||||||
| TVNu57 | 0.06 ± 0.01 d | 0.06 ± 0.01 d | 28.00 ± 0.55 a,b,c,d | 27.61 ± 0.48 a,b,c,d | ||||||
| TVNu138 | 0.23 ± 0.01 c,d | 0.21 ± 0.01 c,d | 27.19 ± 0.62 a,b,c,d | 26.80 ± 0.60 a,b,c,d | ||||||
| TVNu161 | 0.18 ± 0.01 c,d | 0.16 ± 0.01 c,d | 30.02 ± 0.77 a,b | 29.64 ± 0.76 a,b | ||||||
| TVNu758 | 0.16 ± 0.01 c,d | 0.15 ± 0.01 c,d | 27.10 ± 0.30 a,b,c,d | 27.11 ± 0.30 a,b,c,d | ||||||
| TVNu1825 | 0.25 ± 0.02 c,d | 0.23 ± 0.02 c,d | 25.50 ± 0.91 d,e | 25.51 ± 0.91 d,e | ||||||
| TVNu1522 | 0.19 ± 0.01 c,d | 0.17 ± 0.01 c,d | 22.56 ± 0.57 e | 22.57 ± 0.57 e | ||||||
| TVNu1388 | 0.18 ± 0.01 c,d | 0.16 ± 0.01 c,d | 26.53 ± 0.69 a,b,c,d | 26.54 ± 0.70 a,b,c,d | ||||||
| Analysis of Variance (ANOVA) | ||||||||||
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Model | 69 | 131.740 | 1.909 | 11.989 | <0.0001 | 69 | 22845.864 | 331.099 | 225.891 | <0.0001 |
| Error | 386 | 61.473 | 0.159 | 386 | 565.779 | 1.466 | ||||
| Corrected Total | 455 | 193.213 | 455 | 23411.643 | ||||||
| Type III Sum of Squares Analysis | ||||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Location (Site) | 2 | 0.025 | 0.013 | 0.080 | 0.924 | 2 | 0.052 | 0.026 | 0.018 | 0.982 |
| Genotype (Accessions) | 34 | 88.722 | 2.609 | 16.385 | <0.0001 | 34 | 12987.598 | 381.988 | 260.610 | <0.0001 |
| Location × Genotype | 33 | 0.033 | 0.001 | 0.006 | 1.000 | 33 | 0.000 | 0.000 | 0.000 | 1.000 |
| Water Absorption | Cooking Time (min) | |||||||||
| Species/Accession Number | Site A | Site B | Site A | Site B | ||||||
| Landrace of Vigna vexillata | 1.33 ± 0.11 a | 1.32 ± 0.13 a | 10.24 ± 0.15 d | 10.26 ± 0.15 d | ||||||
| Cowpea (Vigna unguiculata) | 1.27 ± 0.08 a | 1.27 ± 0.08 a | 16.29 ± 0.15 b | 16.31 ± 0.15 b | ||||||
| Rice Bean (Vigna umbellata) | 1.16 ± 0.06 a | 1.16 ± 0.06 a | 13.20 ± 0.12 c | 13.23 ± 0.12 c | ||||||
| AGG53597WVIG1 | 1.35 ± 0.03 a | 1.33 ± 0.02 a | 8.26 ± 0.42 d | 7.87 ± 0.40 d | ||||||
| AGG51603WVIG1 | 1.29 ± 0.01 a | 1.27 ± 0.02 a | 10.15 ± 0.22 d,e | 10.17 ± 0.25 d,e | ||||||
| AGG52867WVIG1 | 1.04 ± 0.04 a | 1.02 ± 0.00 a | 11.27 ± 0.41 d | 11.28 ± 0.42 d | ||||||
| Unknown Vigna legume | 0.43 ± 0.01 a,b | 0.39 ± 0.02 a,b | 29.35 ± 0.31 a | 28.97 ± 0.30 a | ||||||
| Unknown Vigna racemosa | 0.08 ± 0.01 b | 0.08 ± 0.00 b | 30.33 ± 0.48 a | 30.34 ± 0.50 a | ||||||
| Analysis of Variance (ANOVA) | ||||||||||
| Water Absorption Capacity | Cooking Time (min) | |||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Model | 15 | 13.441 | 0.896 | 4.279 | <0.0001 | 15 | 4957.993 | 330.533 | 386.632 | <0.0001 |
| Error | 278 | 58.223 | 0.209 | 278 | 237.663 | 0.855 | ||||
| Corrected Total | 293 | 71.664 | 293 | 5195.656 | ||||||
| Type III Sum of Squares Analysis | ||||||||||
| Source | DF | Sum of Squares | Mean Squares | F | p | DF | Sum of Squares | Mean Squares | F | p |
| Location (Site) | 1 | 0.004 | 0.004 | 0.017 | 0.896 | 1 | 0.006 | 0.006 | 0.007 | 0.934 |
| Genotypes (Accessions) | 7 | 13.436 | 1.919 | 9.165 | <0.0001 | 7 | 4957.947 | 708.278 | 828.489 | <0.0001 |
| Location × Genotype | 7 | 0.003 | 0.000 | 0.002 | 1.000 | 7 | 0.001 | 0.000 | 0.000 | 1.000 |
| Class | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| Object | 47 | 20 | 8 | 2 | 1 | 4 | 4 |
| TVNu324_VRe | TVNu1632_VV | TVNu325_VRe | Check 3 | TVNu1520_VRe | AGG308107WVIG2 _VV | TVNu379_VRe | |
| TVNu342_VA | TVNu1701_VV | Check 2 | TVNu781_VV | AGG62154WVIG_1_VV | TVNu1582_VV | ||
| AGG308101WVIG1 _VV | TVNu1629_VV | Unknown _Vigna reticulata | TVNu1624_VV | TVNu1358_VV | |||
| TVNu1344_VV | TVNu767_VRe | AGG51603WVIG1_VRa | AGG308097WVIG 1_VV | TVNu1394_VRe | |||
| AGG308096 WVIG2_VV | TVNu343_VRe | AGG53597WVIG1_VRa | |||||
| TVNu120 _VV | TVNu333_VV | Check 1 | |||||
| TVNu1529_VV | TVNu1370_VV | TVNu837_VV | |||||
| TVNu720_VA | TVNu349_VRe | AGG52867WVIG1_VRa | |||||
| TVNu223_VA | TVNu1378_VV | ||||||
| TVNu1546_VV | TVNu1405_VRe | ||||||
| TVNu1698_VRe | TVNu1593_VV | ||||||
| TVNu877_VA | Unknown Vigna | ||||||
| TVNu524_VRe | TVNu381_VV | ||||||
| TVNu1699_VA | TVNu479_VV | ||||||
| TVNu1191_VRe | TVNu605 _VRe | ||||||
| TVNu1621_VV | TVNu1360_VV | ||||||
| TVNu607_VRe | TVNu1790_VRe | ||||||
| TVNu56_VRe | TVNu1808_VRe | ||||||
| TVNu- 224_VRe | Unknown_Vigna_racemosa | ||||||
| TVNu739_VRe | TVNu161_VRe | ||||||
| TVNu916_VRe | |||||||
| TVNu955_VV | |||||||
| TVNu1092 _VV | |||||||
| TVNu1591_VV | |||||||
| TVNu178_VV | |||||||
| TVNu293_VV | |||||||
| TVNu1840_VA | |||||||
| AGG17856WVIG_1_VRe | |||||||
| TVNu738_VRe | |||||||
| TVNu1796_VV | |||||||
| TVNu1792_VA | |||||||
| TVNu832_VV | |||||||
| TVNu219_VA | |||||||
| TVNu491_VRe | |||||||
| TVNu1628_VV | |||||||
| TVNu1779_VRe | |||||||
| TVNu138_VRe | |||||||
| AGG308099WVIG2_VV | |||||||
| TVNu1804_VA | |||||||
| TVNu1586_VV | |||||||
| TVNu57_VRe | |||||||
| TVNu1825_VRe | |||||||
| TVNu1644_VA | |||||||
| TVNu758_VRe | |||||||
| TVNu1388_VRe | |||||||
| TVNu1522_VRe | |||||||
| TVNu1185_VA |
| Species | Descriptive Parameters | Water Absorption Capacity | Cooking Time (min) | Yield per Plant (g) | |||
|---|---|---|---|---|---|---|---|
| Site A | Site B | Site A | Site B | Site A | Site B | ||
| Landrace of Vigna vexillata | Mean | 1.33 | 1.32 | 10.24 | 10.26 | 13.45 | 7.62 |
| CV (%) | 9.50 | 9.37 | 1.46 | 1.45 | 4.59 | 6.34 | |
| Range | 0.69–4.01 | 0.70–3.96 | 8.56–11.89 | 8.59–11.91 | 9.00–26.55 | 4.94–17.31 | |
| Cowpea (Vigna unguiculata) | Mean | 1.27 | 1.27 | 16.29 | 16.31 | 52.690 | 26.657 |
| CV (%) | 1.85 | 1.82 | 0.93 | 0.93 | 5.48 | 5.42 | |
| Range | 0.58–1.58 | 0.58–1.57 | 14.06–18.84 | 14.09–18.87 | 28.80–106.08 | 14.71–53.35 | |
| Rice Bean (Vigna umbellata) | Mean | 1.16 | 1.16 | 13.20 | 13.23 | 86.04 | 61.92 |
| CV (%) | 4.05 | 4.02 | 0.92 | 0.91 | 2.378 | 2.361 | |
| Range | 0.67–2.02 | 0.68–2.00 | 11.73–14.98 | 11.76–15.01 | 60.27–109.76 | 43.51–78.86 | |
| Vigna ambacensis | Mean | 0.20 | 0.18 | 23.45 | 23.43 | 1.74 | 0.78 |
| CV (%) | 11.21 | 11.20 | 0.44 | 0.42 | 22.36 | 14.25 | |
| Range | 0.00–0.50 | 0.00–0.58 | 22.25–24.95 | 22.26–24.96 | 0.72–5.36 | 0.43–1.65 | |
| Vigna vexillata | Mean | 0.34 | 0.32 | 25.42 | 25.40 | 16.84 | 12.54 |
| CV (%) | 7.80 | 7.95 | 1.73 | 1.70 | 9.48 | 6.77 | |
| Range | 0.00–1.15 | 0.00–1.13 | 15.54–31.28 | 15.55–31.30 | 9.48–63.00 | 7.61–35.26 | |
| Vigna reticulata | Mean | 0.39 | 0.37 | 26.77 | 26.78 | 10.60 | 6.78 |
| CV (%) | 13.83 | 14.04 | 1.20 | 1.27 | 10.55 | 10.62 | |
| Range | 0.00–2.24 | 0.00–2.08 | 16.60–30.98 | 16.58–40.00 | 4.32–30.36 | 2.58–17.69 | |
| Vigna racemosa | Mean | 0.84 | 0.81 | 17.70 | 17.72 | 28.25 | 18.28 |
| CV (%) | 16.70 | 17.06 | 14.83 | 14.81 | 37.02 | 38.37 | |
| Range | 0.00–1.69 | 0.00–1.66 | 7.11–31.19 | 7.12–31.22 | 2.08–49.00 | 1.21–34.70 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harouna, D.V.; Venkataramana, P.B.; Matemu, A.O.; Ndakidemi, P.A. Assessment of Water Absorption Capacity and Cooking Time of Wild Under-Exploited Vigna Species towards their Domestication. Agronomy 2019, 9, 509. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090509
Harouna DV, Venkataramana PB, Matemu AO, Ndakidemi PA. Assessment of Water Absorption Capacity and Cooking Time of Wild Under-Exploited Vigna Species towards their Domestication. Agronomy. 2019; 9(9):509. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090509
Chicago/Turabian StyleHarouna, Difo Voukang, Pavithravani B. Venkataramana, Athanasia O. Matemu, and Patrick Alois Ndakidemi. 2019. "Assessment of Water Absorption Capacity and Cooking Time of Wild Under-Exploited Vigna Species towards their Domestication" Agronomy 9, no. 9: 509. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090509