Regulation of Spraying 6-BA in the Late Jointing Stage on the Fertile Floret Development and Grain Setting in Winter Wheat

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Design
2.2. Measurements and Methods
2.2.1. Observations of Young Spike Development in Wheat
2.2.2. Measurements of Dry Matter in Spike and Non-Spike Organs
2.2.3. Measurements of Soluble Sugar and Sucrose Contents
2.2.4. Measurements of Cytokinin and Auxin Contents
2.2.5. Measurements of Yield and Yield Components
2.3. Formulae for Calculations
2.4. Statistical Analysis
3. Results and Analysis
3.1. The Effects of 6-BA Application on the Dynamics of Floret Development in Wheat
3.2. The Effects of 6-BA Application on the Number of Fertile Florets and the Number of Grains in Wheat
3.3. The Effects of 6-BA Application on the Accumulation and Distribution of Dry Matter in the Spike and Non-Spike Organs in Wheat
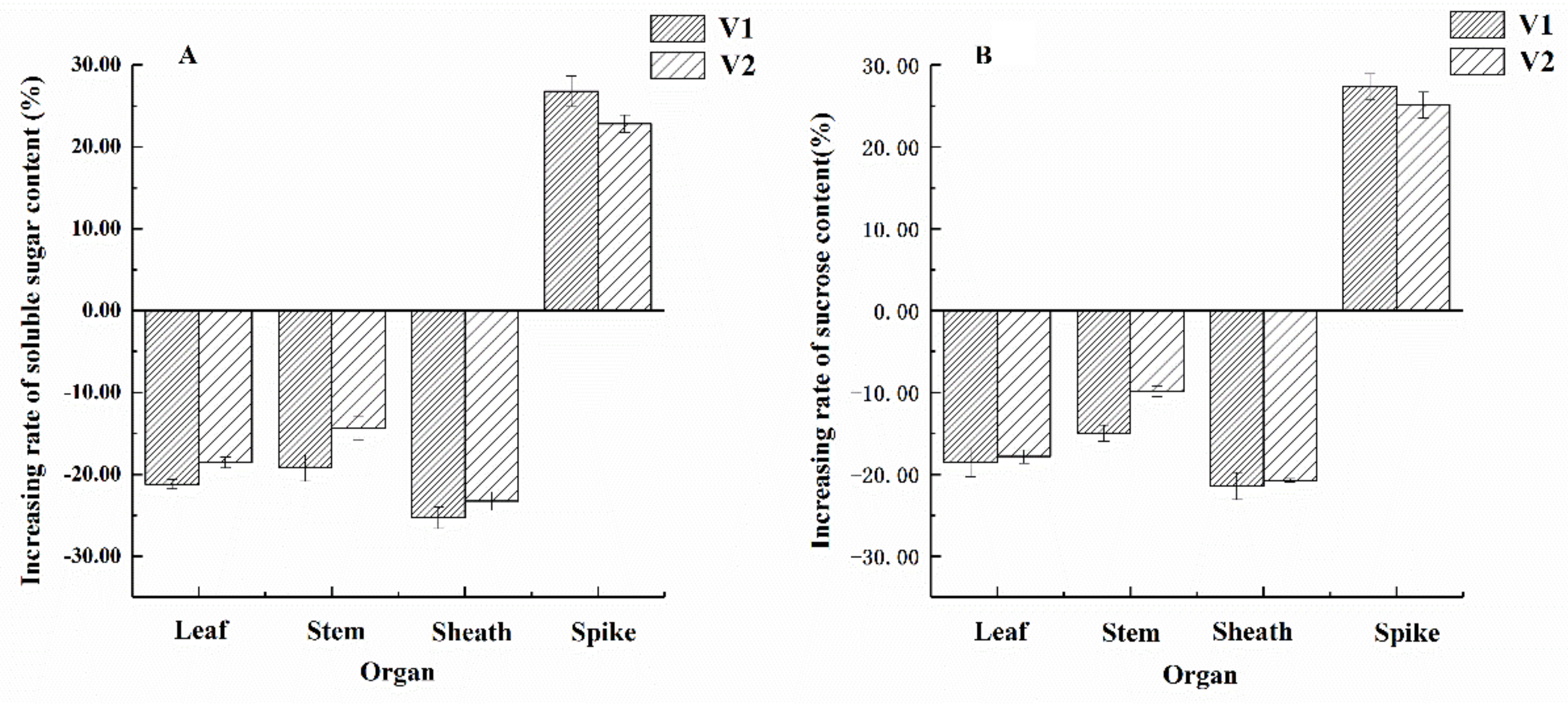
3.4. Effects of 6-BA Application on Sugar Content in Different Organs During Anthesis in Wheat
3.5. The Effects of 6-BA Application on Endogenous Hormone Content in Spikes
3.6. Relationships Among Spike Dry Matter, Sugars, Hormones, and the Number of Florets per Spike and Grain Number per Spike at Anthesis
3.7. The effects of 6-BA Application on the Developmental Morphological Characteristics of Fertile Florets and Grains in Wheat
3.8. Effects of 6-BA Foliar Application on Yield and Yield Components in Wheat
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jiao, X.-Q.; Mongol, N.; Zhang, F.-S. The transformation of agriculture in China: Looking back and looking forward. J. Integr. Agric. 2018, 17, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, J.; Daliakopoulos, I.N.; Del Moral, F.; Hueso, J.J.; Tsanis, I.K. A Review of Soil-Improving Cropping Systems for Soil Salinization. Agronomy 2019, 9, 295. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Toulmin, C.; Pretty, J.; Robinson, S.; Thomas, S.M. Food security: the challenge of feeding 9 billion People. Science 2012, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Wu, Y.; Zhou, Y.; Ren, X.; Shao, Y.; Feng, W.; Zhu, Y.; He, L.; Guo, T. Approach to Higher Wheat Yield in the Huang-Huai Plain: Improving Post-anthesis Productivity to Increase Harvest Index. Front. Plant Sci. 2018, 9, 1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuire, S. FAO, IFAD, and WFP. The State of Food Insecurity in the World 2015: Meeting the 2015 International Hunger Targets: Taking Stock of Uneven Progress; FAO: Rome, Italy, 2015. Adv. Nutr. 2015, 6, 623–624. [Google Scholar] [CrossRef] [PubMed]
- Tyczewska, A.; Woźniak, E.; Gracz, J.; Kuczyński, J.; Twardowski, T. Towards Food Security: Current State and Future Prospects of Agrobiotechnology. Trends Biotechnol. 2018, 36, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zhang, F.; Liu, C.; Yu, H.; Cao, B.; Tian, S.; Liao, Y.; Siddique, K.H. Wheat yield improvements in China: Past trends and future directions. Field Crop. Res. 2015, 177, 117–124. [Google Scholar] [CrossRef]
- Sadras, V.O.; Slafer, G.A. Environmental modulation of yield components in cereals: Heritabilities reveal a hierarchy of phenotypic plasticities. Field Crop. Res. 2012, 127, 215–224. [Google Scholar] [CrossRef]
- Duan, J.; Wu, Y.; Zhou, Y.; Ren, X.; Shao, Y.; Feng, W.; Zhu, Y.; Wang, Y.; Guo, T. Grain number responses to pre-anthesis dry matter and nitrogen in improving wheat yield in the Huang-Huai Plain. Sci. Rep. 2018, 8, 7126. [Google Scholar] [CrossRef]
- Acreche, M.M.; Briceño-Félix, G.; Sánchez, J.A.M.; Slafer, G.A. Physiological bases of genetic gains in Mediterranean bread wheat yield in Spain. Eur. J. Agron. 2008, 28, 162–170. [Google Scholar] [CrossRef]
- Reynolds, M.; Foulkes, M.J.; Slafer, G.A.; Berry, P.; Parry, M.A.J.; Snape, J.W.; Angus, W.J. Raising yield potential in wheat. J. Exp. Bot. 2009, 60, 1899–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, F.G.; Miralles, D.J.; Slafer, G.A. Wheat floret survival as related to pre-anthesis spike growth. J. Exp. Bot. 2011, 62, 4889–4901. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Slafer, G.A.; Schnurbusch, T. Genotypic variation in spike fertility traits and ovary size as determinants of floret and grain survival rate in wheat. J. Exp. Bot. 2016, 67, 4221–4230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.M.; Guo, T.C.; Zhu, Y.J.; Wang, C.Y.; Ma, X.M. Ear of Wheat, 1st ed.; China Agricultural Press: Beijing, China, 2008. (In Chinese) [Google Scholar]
- Zheng, C.F.; Zhu, Y.J.; Wang, C.Y.; Guo, T.C. Wheat grain yield increase in response to pre-anthesis foliar application of 6-benzylaminopurine is dependent on floret development. PLoS ONE 2016, 11, e0156627. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Navarro, O.E.; Griffiths, S.; Molero, G.; Reynolds, M.P.; Slafer, G.A. Dynamics of floret development determining differences in spike fertility in an elite population of wheat. Field Crop. Res. 2015, 172, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, A.; Savin, R.; Slafer, G.A. Floret development and grain setting differences between modern durum wheats under contrasting Nitrogen availability. J. Exp. Bot. 2013, 64, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Serrago, R.A.; Miralles, D.J.; Slafer, G.A. Floret fertility in wheat as affected by photoperiod during stem elongation and removal of spikelets at booting. Eur. J. Agron. 2008, 28, 301–308. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Wockner, K.B.; Palta, J.A.; McIntyre, C.L.; Borgognone, M.G.; Bourgault, M.; Reynolds, M.P.; Miralles, D.J. More fertile florets and grains per spike can be achieved at higher temperature in wheat lines with high spike biomass and sugar content at booting. Funct. Plant Boil. 2014, 41, 482–495. [Google Scholar] [CrossRef] [Green Version]
- Bartrina, I.; Otto, E.; Strnad, M.; Werner, T.; Schmülling, T. Cytokinin Regulates the Activity of Reproductive Meristems, Flower Organ Size, Ovule Formation, and Thus Seed Yield in Arabidopsis thaliana [C][W][OA]. Plant Cell 2011, 23, 69–80. [Google Scholar] [CrossRef]
- Jameson, P.E.; Song, J.C. Cytokinin: A key driver of seed yield. J. Exp. Bot. 2016, 67, 593–606. [Google Scholar] [CrossRef]
- Zhang, K.R.; Diederich, L.; John, P. The Cytokinin Requirement for Cell Division in Cultured Nicotiana plumbaginifolia Cells Can Be Satisfied by Yeast Cdc25 Protein Tyrosine Phosphatase. Implications for Mechanisms of Cytokinin Response and Plant Development. Plant Physiol. 2004, 137, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Meng, X.; Liu, X.; Liu, T.; Wang, H.; Jia, Z.; Yang, D.; Ren, X. Exogenous Hormonal Application Regulates the Occurrence of Wheat Tillers by Changing Endogenous Hormones. Front. Plant Sci. 2018, 9, 1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.Q.; Li, Y.L.; Ni, Y.L.; Yin, Y.P.; Yang, W.B.; Cui, Z.Y.; Zhang, Y.T.; Ma, R.Y.; Wang, Z.L. Effects of Exogenous ABA and 6-BA on Protein Content and Grain Filling Process in Different Types of Stay-Green Wheat. Acta Agron. Sin. 2016, 40, 301–312. (In Chinese) [Google Scholar] [CrossRef]
- Yang, D.Q.; Luo, Y.L.; Dong, W.H.; Yin, Y.P.; Li, Y.; Wang, Z.L. Response of photosystem II performance and antioxidant enzyme activities in stay-green wheat to cytokinin. Photosynthetica 2018, 56, 567–577. [Google Scholar] [CrossRef]
- Luo, Y.; Tang, Y.; Zhang, X.; Li, W.; Chang, Y.; Pang, D.; Xu, X.; Li, Y.; Wang, Z. Interactions between cytokinin and nitrogen contribute to grain mass in wheat cultivars by regulating the flag leaf senescence process. Crop. J. 2018, 6, 538–551. [Google Scholar] [CrossRef]
- Shi, H.; Wang, B.; Yang, P.; Li, Y.; Miao, F. Differences in Sugar Accumulation and Mobilization between Sequential and Non-Sequential Senescence Wheat Cultivars under Natural and Drought Conditions. PLoS ONE 2016, 11, 0166155. [Google Scholar] [CrossRef]
- Wang, W.; Hao, Q.; Tian, F.; Li, Q.; Wang, W. Cytokinin-Regulated Sucrose Metabolism in Stay-Green Wheat Phenotype. PLoS ONE 2016, 11, e0161351. [Google Scholar] [CrossRef]
- Wang, Z.-Q.; Zhang, W.-Y.; Yang, J.-C. Physiological mechanism underlying spikelet degeneration in rice. J. Integr. Agric. 2018, 17, 1475–1481. [Google Scholar] [CrossRef] [Green Version]
- Sreenivasulu, N.; Schnurbusch, T. A genetic playground for enhancing grain number in cereals. Trends Plant Sci. 2012, 17, 91–101. [Google Scholar] [CrossRef]
- Yang, D.; Li, Y.; Shi, Y.; Cui, Z.; Luo, Y.; Zheng, M.; Chen, J.; Li, Y.; Yin, Y.; Wang, Z. Exogenous Cytokinins Increase Grain Yield of Winter Wheat Cultivars by Improving Stay-Green Characteristics under Heat Stress. PLoS ONE 2016, 11, e0155437. [Google Scholar] [CrossRef] [PubMed]
- Miralles, D.J.; Richards, R.A.; Slafer, G.A. Duration of the stem elongation period influences the number of fertile florets in wheat and barley. Funct. Plant Boil. 2000, 27, 931. [Google Scholar] [CrossRef]
- Prystupa, P.; Savin, R.; Slafer, G.A. Grain number and its relationship with dry matter, N and P in the spikes at heading in response to N × P fertilization in barley. Field Crops Res. 2004, 90, 245–254. [Google Scholar] [CrossRef]
- Ghiglione, H.O.; Gonzalez, F.G.; Serrago, R.; Maldonado, S.B.; Chilcott, C.; Curá, J.A.; Miralles, D.J.; Zhu, T.; Casal, J.J. Autophagy regulated by day length determines the number of fertile florets in wheat. Plant J. 2008, 55, 1010–1024. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Gao, K.; Hu, W.; Snider, J.L.; Wang, S.; Chen, B.; Zhou, Z. Chemical priming of seed alters cotton floral bud differentiation by inducing changes in hormones, metabolites and gene expression. Plant Physiol. Biochem. 2018, 130, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Lebon, G.; Wojnarowiez, G.; Holzapfel, B.; Fontaine, F.; Vaillant-Gaveau, N.; Clément, C. Sugars and flowering in the grapevine (Vitis vinifera L.). J. Exp. Bot. 2008, 59, 2565–2578. [Google Scholar] [CrossRef] [PubMed]
- Emery, R.J.N.; Ma, Q.F.; Atkins, C.A. The Forms and Sources of Cytokinins in Developing White Lupine Seeds and Fruits. Plant Physiol. 2002, 123, 1593–1604. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.D. Essential Roles of Local Auxin Biosynthesis in Plant Development and in Adaptation to Environmental Changes. Annu. Rev. Plant Biol. 2018, 69, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Chika, N.; Yoshi, O.; Shusei, S.; Tomohiko, K.; Satoshi, T.; Chiharu, U. Histidine Kinase Homologs That Act as Cytokinin Receptors Possess Overlapping Functions in the Regulation of Shoot and Root Growth in Arabidopsis. Plant Cell 2004, 16, 1365–1377. [Google Scholar] [Green Version]
- Schaller, G.E.; Bishopp, A.; Kieber, J.J. The Yin-Yang of Hormones: Cytokinin and Auxin Interactions in Plant Development. Plant Cell 2015, 27, 44–63. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita-Tsujimura, K.; Kakimoto, T. Cytokinin receptors in sporophytes are essential for male and female functions in Arabidopsis thaliana. Plant Signal. Behav. 2011, 6, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.M.; Zheng, Y.L. The Influence of Phytohormone on the Duration of Spike Differentiation in Multispiklet Wheat. J. Triticeae Crop. 2000, 20, 35–38. (In Chinese) [Google Scholar]
- Roitsch, T.; Ehneß, R. Regulation of source/sink relations by cytokinins. Plant Growth Regul. 2000, 32, 359–367. [Google Scholar] [CrossRef]
- Yang, D.; Dong, W.; Luo, Y.; Song, W.; Cai, T.; Li, Y.; Yin, Y.; Wang, Z. Effects of exogenous 6-BA on photosynthetic characteristics and endogenous hormone content in wheat leaves under two nitrogen application levels at seedling stage. Sci. Agric. Sin. 2017, 50, 3871–3884. (In Chinese) [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Dry Weight (g) | Soluble Sugar (mg·g−1 DW) | Sucrose (mg·g−1 DW) | Auxin (ng·g−1 FW) | Cytokinin (ug·g−1 FW) |
|---|---|---|---|---|---|
| Number of fertile florets per spike | 0.63 ** | 0.50 ** | 0.59 ** | 0.34 | 0.65 ** |
| Grain number per spike | 0.60 ** | 0.47 * | 0.51 * | 0.17 | 0.55 ** |
| Year | Treatment | SN | GN | TGW | GY |
|---|---|---|---|---|---|
| 2016–2017 | V1S0 | 457.67 b | 35.33 b | 41.31 a | 6954.63 b |
| V1S1 | 464.84 b | 38.67 a | 42.74 a | 7214.88 a | |
| V2S0 | 545.70 a | 27.67 d | 36.85 b | 6344.48 d | |
| V2S1 | 551.38 a | 30.33 c | 38.11 b | 6541.70 c | |
| 2017–2018 | V1S0 | 513.66 b | 36.00 b | 47.35 a | 7355.59 b |
| V1S1 | 529.49 b | 39.67 a | 48.77 a | 7750.39 a | |
| V2S0 | 615.36 a | 28.67 d | 42.76 b | 6816.70 c | |
| V2S1 | 622.03 a | 31.67 c | 43.71 b | 7167.03 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Song, M.; Duan, J.; Yang, J.; Zhu, Y.; Zhou, S. Regulation of Spraying 6-BA in the Late Jointing Stage on the Fertile Floret Development and Grain Setting in Winter Wheat. Agronomy 2019, 9, 546. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090546
Li S, Song M, Duan J, Yang J, Zhu Y, Zhou S. Regulation of Spraying 6-BA in the Late Jointing Stage on the Fertile Floret Development and Grain Setting in Winter Wheat. Agronomy. 2019; 9(9):546. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090546
Chicago/Turabian StyleLi, Shengnan, Miao Song, Jianzhao Duan, Jiaheng Yang, Yunji Zhu, and Sumei Zhou. 2019. "Regulation of Spraying 6-BA in the Late Jointing Stage on the Fertile Floret Development and Grain Setting in Winter Wheat" Agronomy 9, no. 9: 546. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090546