Infrared Thermography to Estimate Vine Water Status: Optimizing Canopy Measurements and Thermal Indices for the Varieties Merlot and Moscato in Northern Italy

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set-Up
2.2. Water Potential Measurements
2.3. Gas Exchange Measurements
2.4. Thermal Image Acquisition
2.5. Thermal Indices Calculation
2.6. Statistical Analysis
3. Results and Discussion
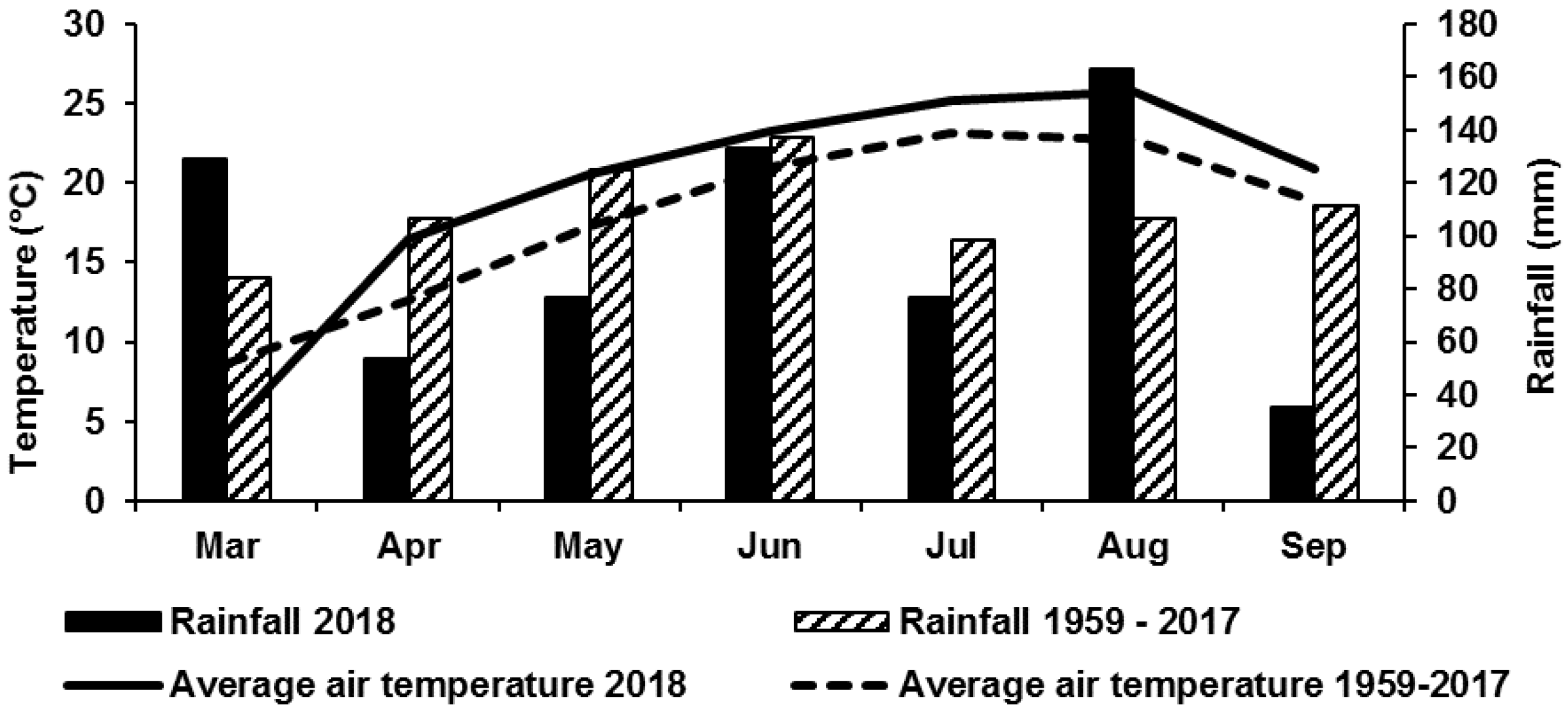
3.1. Weather Conditions During the Trial Period
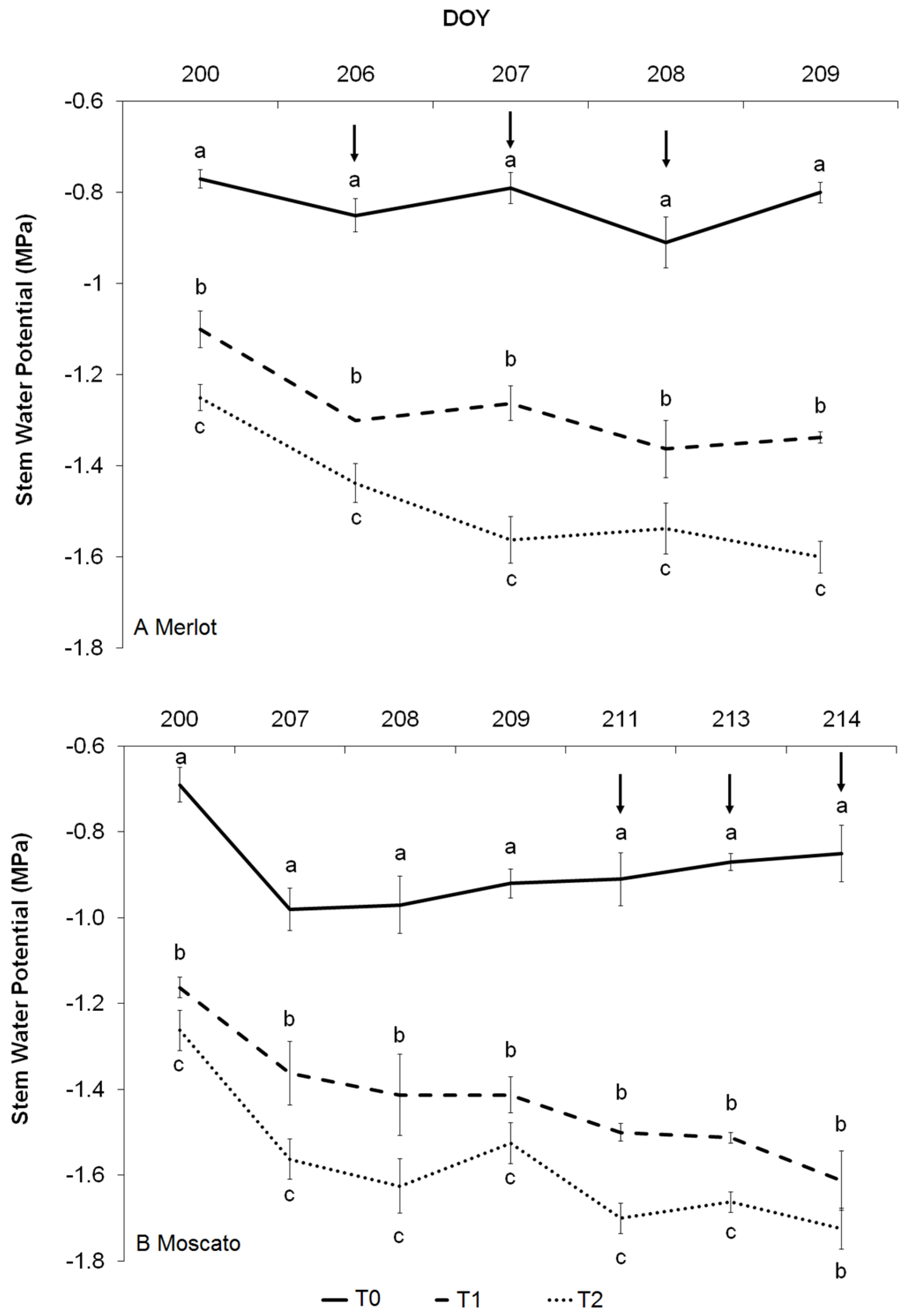
3.2. Water Potential Measurements
3.3. Gas Exchange Measurements
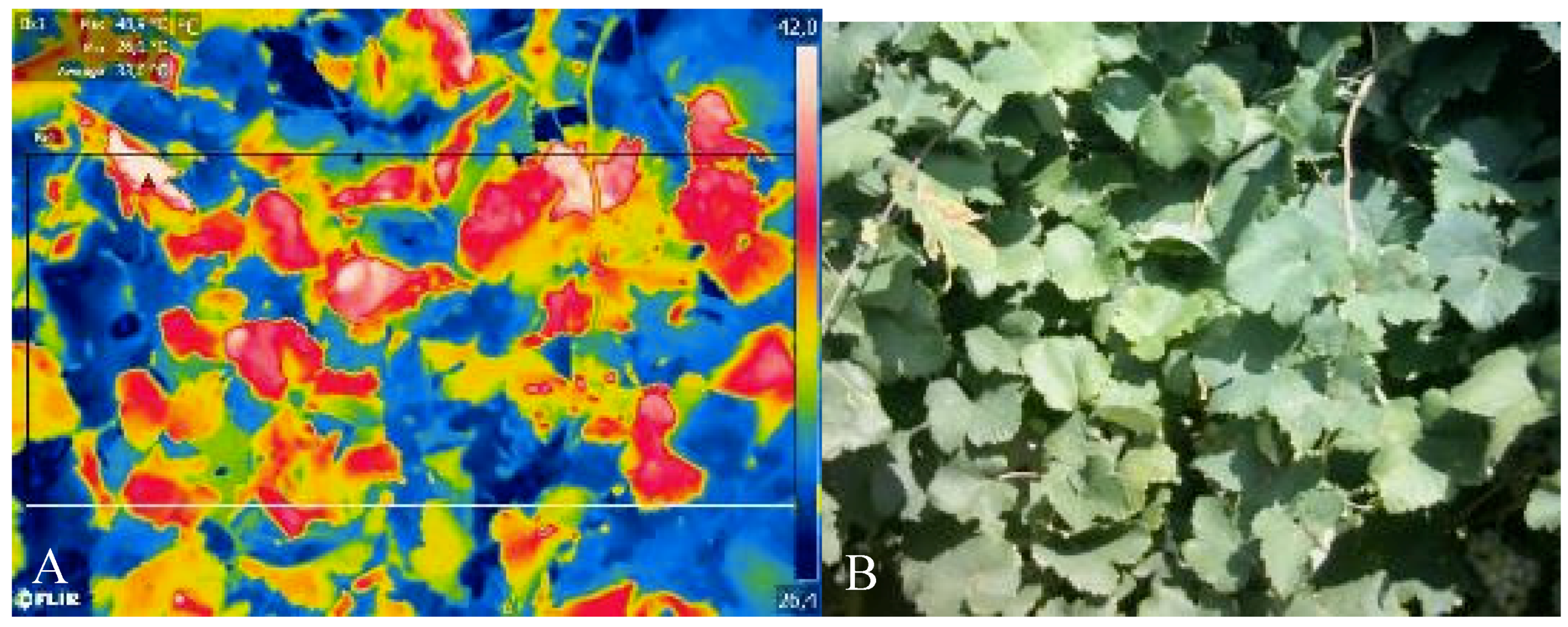
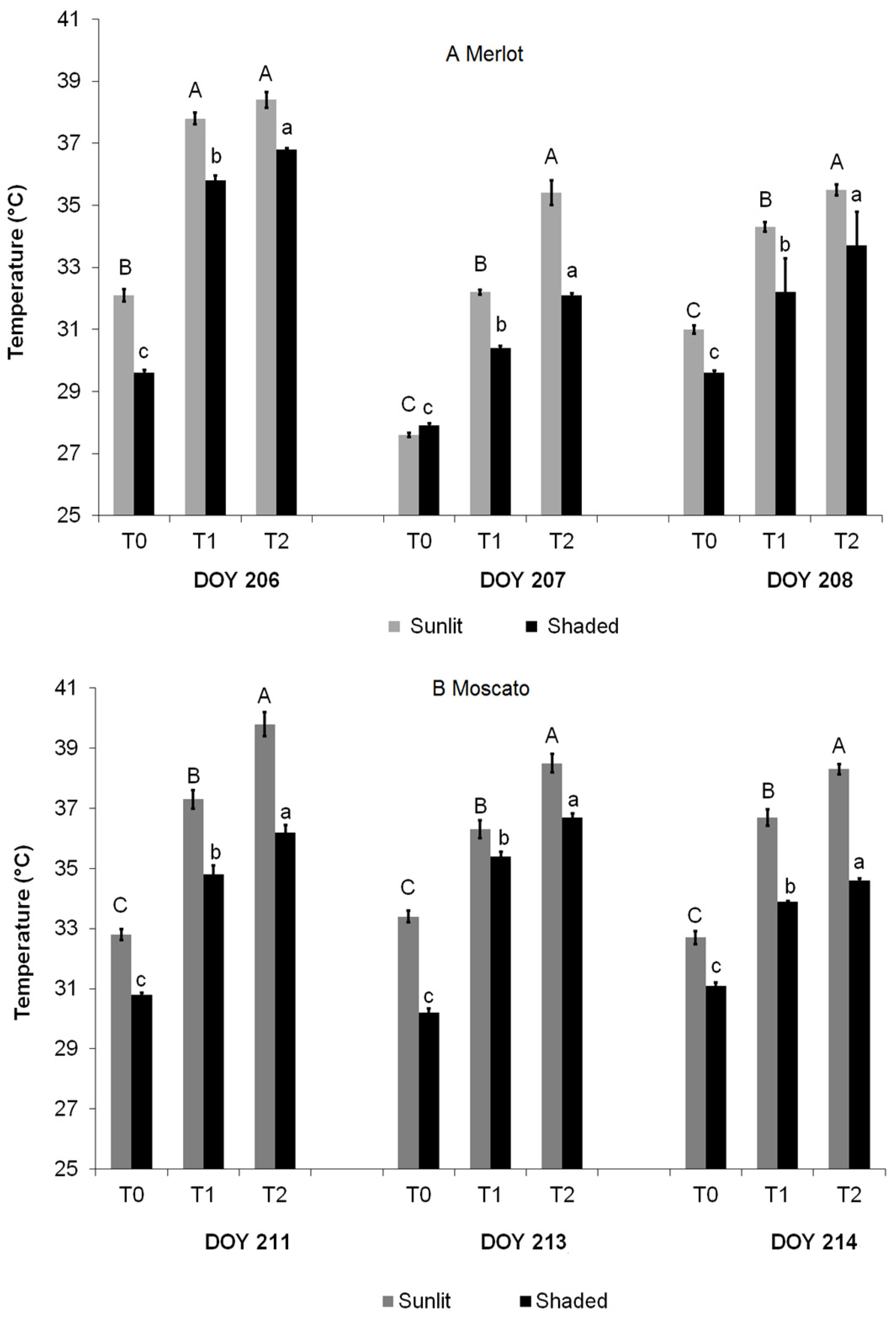
3.4. Infrared Thermography
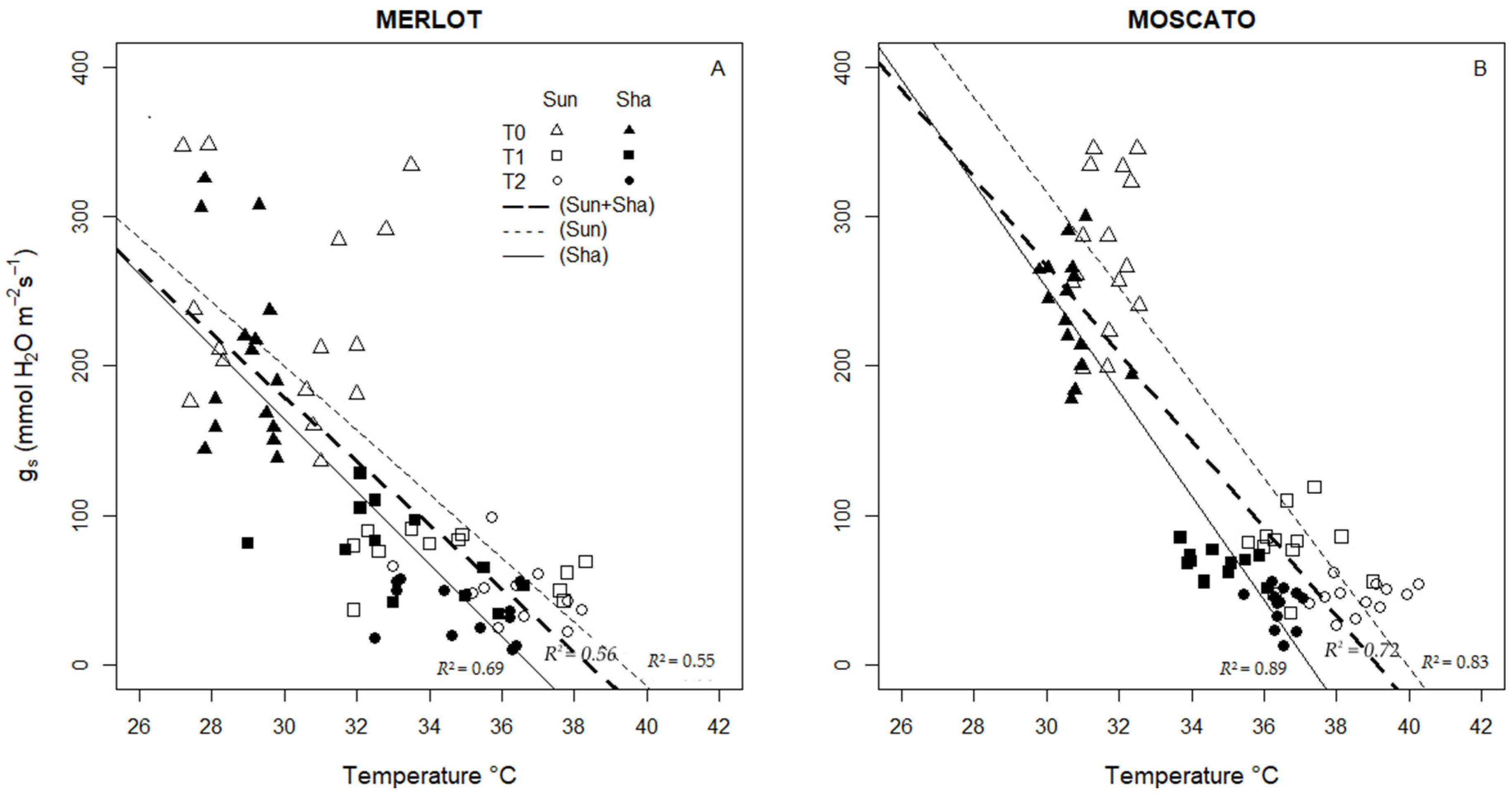
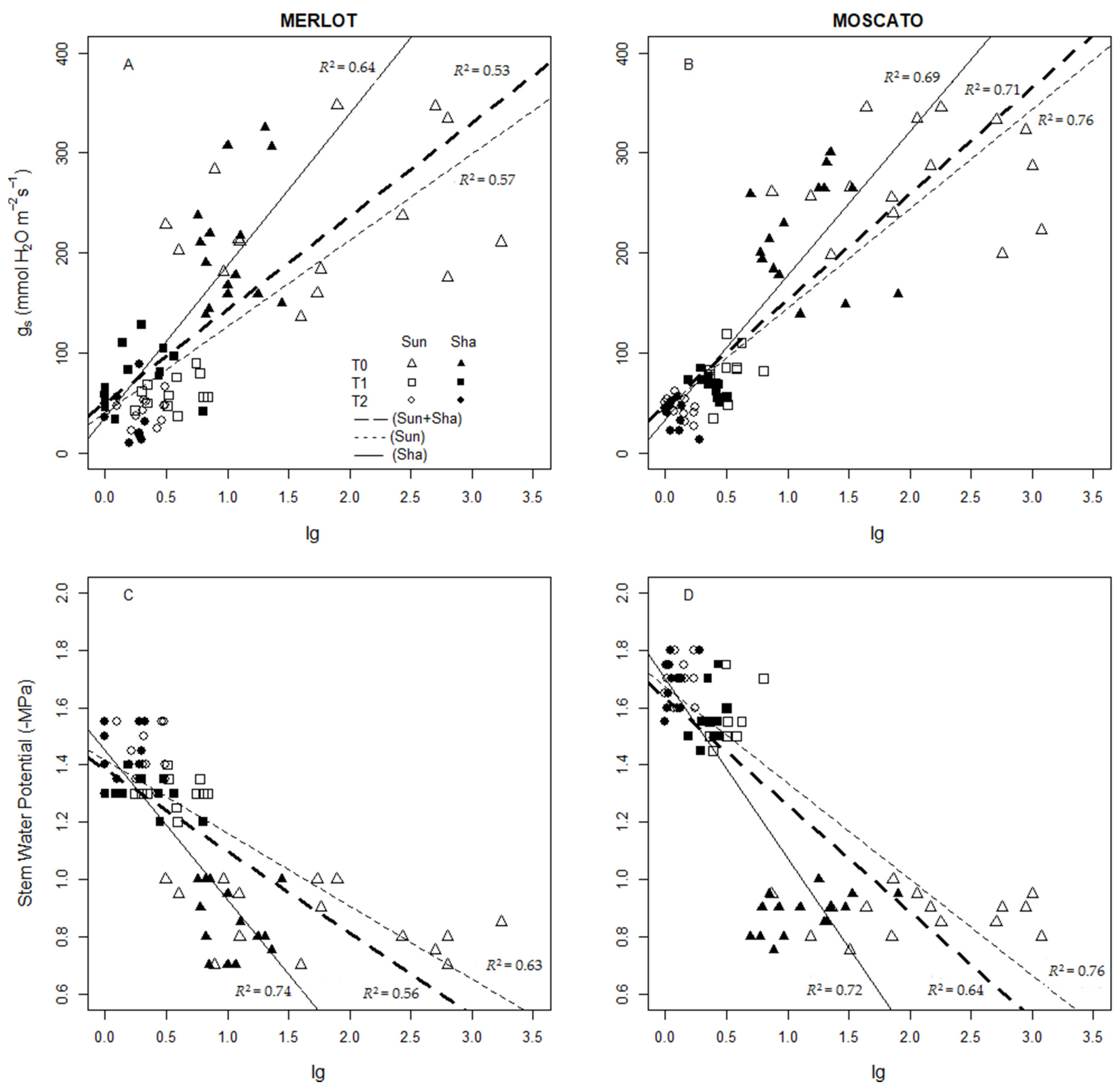
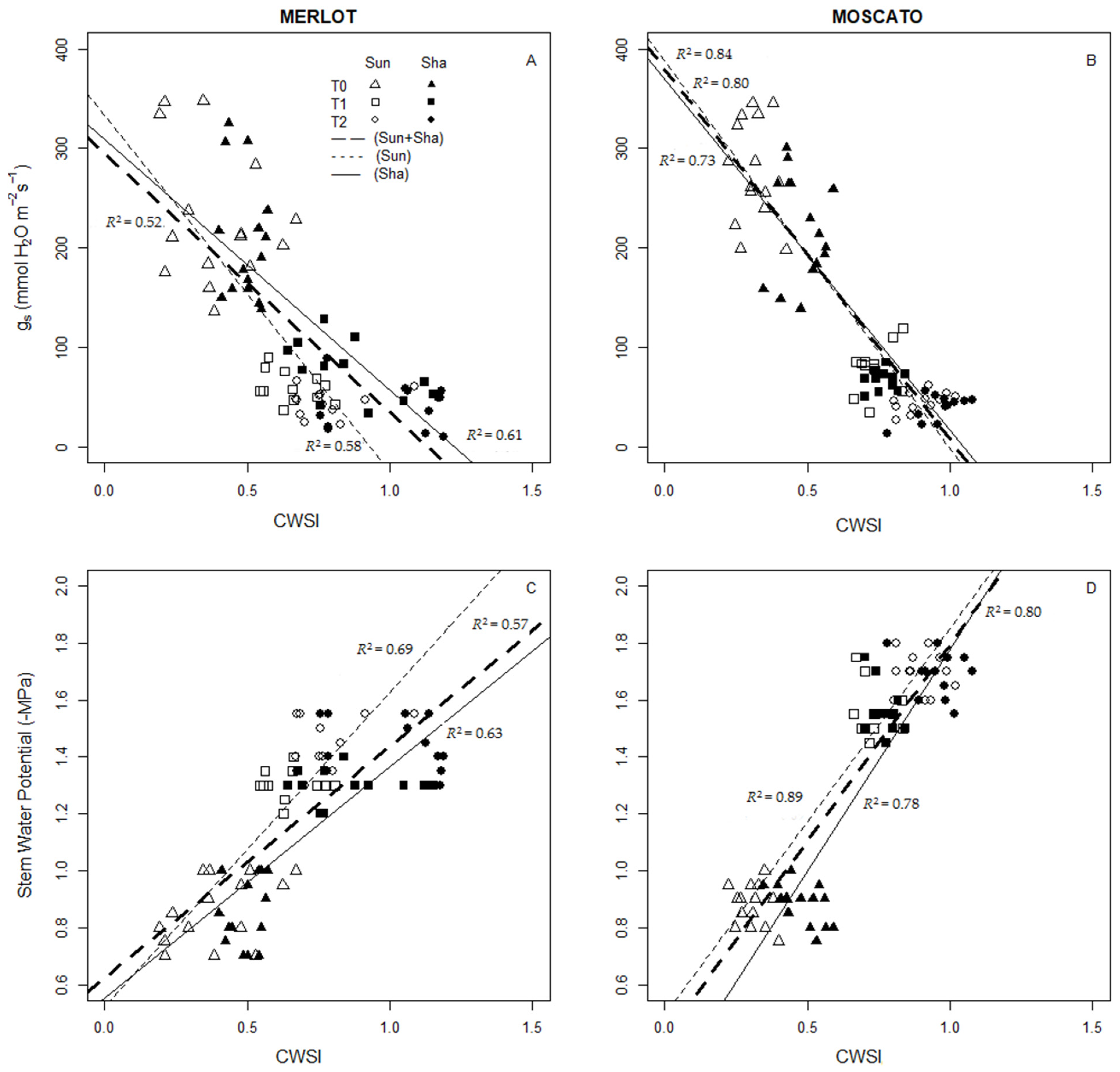
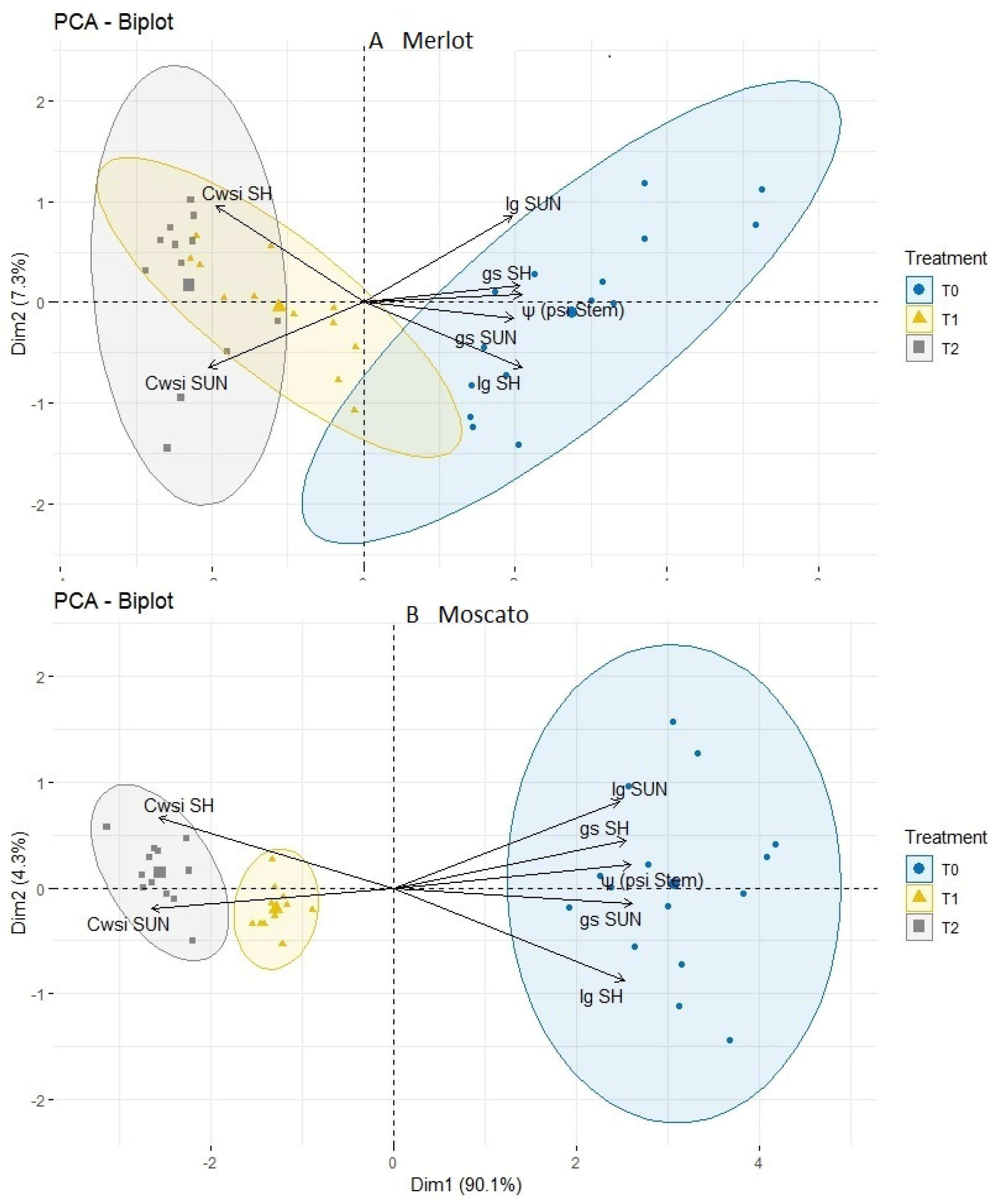
3.5. Thermal Indices: Values and Interaction with Physiological Measurements
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. 2013: Allegato III: Glossario. In Climate Change 2013: The Physical Science Basis; Planton, S., Ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; ISBN 978-92-9169-138-8. [Google Scholar]
- Maracchi, G.; Sirotenko, O.; Bindi, M. Impacts of present and future Climate Variability on Agricolture and Forestry in the Temperate Regions: Europe. Clim. Chang. 2005, 70, 117–135. [Google Scholar] [CrossRef]
- Luterbacher, J.; Xoplaki, E.; Casty, C.; Wanner, H.; Pauling, A.; Küttel, M.; Brönnimann, S.; Fischer, E.; Fleitmann, D.; Gonzalez-Rouco, F.J.; et al. Mediterranean climate variability over the last centuries: A review. In The Mediterranean Climate: An Overview of the Main Characteristics and Issues; Lionello, P., Malanotte-Rizzoli, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 27–148. [Google Scholar]
- Petit, J.R.; Jouzel, J.; Raynaud, D.; Barkov, N.I.; Barnola, J.M.; Basile, I.; Bender, M.; Chappellaz, J.; Davis, M.; Delaygue, G.; et al. Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 1999, 399, 429–436. [Google Scholar] [CrossRef]
- Miranda, P.M.A.; Valente, M.A.; Tomé, A.R.; Trigo, R.; Coelho, M.F.E.S.; Aguiar, A.; Azevedo, E.B. O clima de Portugal nos se´culos XX e XXI. In Alteracxao es Clima´ticasem Portugal; Cena´rios, Impactes e Medidas de Adaptacxao; Santos, F.D., Miranda, P., Eds.; Gradiva: Lisboa, Portugal, 2006; pp. 45–113. [Google Scholar]
- Moriondo, M.; Jones, G.; Bois, B.; Dibari, C.; Ferrise, R.; Trombi, G.; Bindi, M. Projected shifts of wine regions in response to climate change. Clim. Chang. 2013, 119, 825–839. [Google Scholar] [CrossRef]
- Hannah, L.; Roehrdanz, P.; Ikegami, M.; Shepard, A.; Shaw, M.; Tabor, G.; Zhi, L.; Marquet, P.A.; Hijmans, R. Climate change, wine, and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 6907–6912. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.R. Climate change and viticulture: A European perspective on climatology, carbon dioxide and UV B effects. Aust. J. Grape Wine Res. 2000, 6, 2–12. [Google Scholar] [CrossRef]
- Schultz, H.; Jones, G. Climate Induced Historic and Future Changes in Viticulture. J. Wine Res. 2010, 21, 137–145. [Google Scholar] [CrossRef]
- Jones, G.V.; Davis, R.E. Climate influences on grapevine phenology, grape composition, and wine production and quality for Bordeaux, France. Am. J. Enol. Vitic. 2000, 51, 249–261. [Google Scholar]
- Meier, N.; Rutishauer, T.; Pfister, C.; Wanner, H.; Luterbacher, J. Grape harvest dates as a proxy for Swiss April to August temperature reconstruction back to AD 1480. Geophys. Res. Lett. 2007, 34, l20705. [Google Scholar] [CrossRef]
- Chuine, I.; Yiou, P.; Viovy, N.; Seguin, B.; Daux, V.; Le Roy Ladurie, E. Historical phenology: Grape ripening as a past climate indicator. Nature 2004, 432, 289–290. [Google Scholar] [CrossRef]
- Fila, G.; Tomasi, D.; Gaiotti, F.; Jones, G.V. The Book of Vinesprouts of Kőszeg (Hungary): A documentary source for reconstructing spring temperatures back to the eighteenth century. Int. J. Biometeorol. 2016, 60, 207–219. [Google Scholar] [CrossRef]
- Tomasi, D.; Jones, G.V.; Giust, M.; Lovat, L.; Gaiotti, F. Grapevine Phenology and Climate Change: Relationships and Trends in the Veneto Region of Italy for 1964–2009. Am. J. Enol. Vitic. 2011, 62, 329–339. [Google Scholar] [CrossRef]
- Fraga, H.; García de Cortázar Atauri, I.; Malheiro, A.; Santos, J. Modelling climate change impacts on viticultural yield, phenology and stress conditions in Europe. Glob. Chang. Biol. 2016, 22, 3774–3788. [Google Scholar] [CrossRef] [PubMed]
- Fraga, H.; Santos, J.A.; Moutinho–Pereira, J.; Carlos, C.; Silvestre, J.; Eiras-Dias, J.; Mota, T.; Malheiro, A.C. Statistical modelling of grapevine phenology in Portuguese wine regions: Observed trends and climate change projections. J. Agric. Sci. 2016, 154, 795–811. [Google Scholar] [CrossRef]
- Schultz, H.R.; Stoll, M. Some critical issues in environmental physiology of grapevines: Future challenges and current limitations. Aust. J. Grape Wine Res. 2010, 16, 4–24. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulías, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef]
- Medrano, H.; Escalona, J.M.; Cifre, J.; Bota, J.; Flexas, J. A ten-year study on the physiology of two Spanish grapevine varieties under field conditions: Effects of water availability from leaf photosynthesis to grape yield and quality. Funct. Plant Biol. 2003, 30, 607–619. [Google Scholar] [CrossRef]
- Palliotti, A.; Tombesi, S.; Silvestroni, O.; Lanari, V.; Gatti, M.; Poni, S. Changes in vineyard establishment and canopy management urged by earlier climate-related grape ripening. Sci. Hortic. 2014, 178, 43–54. [Google Scholar] [CrossRef]
- Chonè, X.; Van Leeuwen, C.; Dubourdieu, D.; Gaugaudillere, J.P. Stem Water Potential is a Sensitive Indicator of Grapevine Water Status. Ann. Bot. 2001, 87, 477–483. [Google Scholar] [CrossRef]
- Pisciotta, A.; Di Lorenzo, R.; Santalucia, G.; Barbagallo, M. Response of grapevine (Cabernet Sauvignon cv) to above ground and subsurface drip irrigation under arid conditions. Agric. Water Manag. 2018, 197, 122–131. [Google Scholar] [CrossRef]
- Dry, P.R.; Loveys, B.R.; McCarthy, M.G.; Stoll, M. Strategic irrigation management in Australian vineyards. J. Int. Sci. Vigne Vin 2001, 35, 129–139. [Google Scholar] [CrossRef]
- Escalona, J.; Flexas, J. Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines. Aust. J. Plant Physiol. 2000, 26, 421–433. [Google Scholar] [CrossRef]
- Tanner, C.B. Plant Temperatures. Agron. J. 1963, 55, 210–211. [Google Scholar] [CrossRef]
- Cifre, J.; Bota, J.; Escalona, J.M.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.). An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Jones, H.G. Irrigation scheduling: Advantages and pitfalls of plant-based methods. J. Exp. Bot. 2004, 55, 2427–2436. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Stoll, M.; Santos, T.; De Sousa, C.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Idso, S.B.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J.; Hatfield, J.L. Normalizing thestress-degree-day parameter for environmental variability. Agric. Meteorol. 1981, 24, 45–55. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J. Canopy Temperature as a Crop Water Stress Indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Jones, H.G. Use of thermography for quantitative studies of spatial and temporal variation of stomatal conductance over leaf surfaces. Plant Cell Environ. 1999, 22, 1043–1055. [Google Scholar] [CrossRef]
- Jones, H.G.; Leinonen, I. Thermal imaging for the study of plant water relations. J. Agric. Meteorol. 2003, 59, 205–217. [Google Scholar] [CrossRef]
- Grant, O.M.; Tronina, L.; Jones, H.G.; Chaves, M.M. Optimizing thermal imaging as a technique for detecting stomatal closure induced by drought stress under green-house conditions. Physiol. Plant 2006, 127, 507–518. [Google Scholar] [CrossRef]
- Grant, O.M.; Tronina, L.; Jones, H.G.; Chaves, M.M. Exploring thermal imaging variables for the detection of stress responses in grapevine under different irrigation regimes. J. Exp. Bot. 2007, 58, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, S.; De Bei, R.; Pech, J.; Tyerman, S. Computational water stress indices obtained from thermal image analysis of grapevine canopies. Irrig. Sci. 2012, 30, 523–536. [Google Scholar] [CrossRef]
- Costa, J.M.; Ortuno, M.F.; Lopes, C.M.; Chaves, M.M. Grapevine variety exhibiting differences in stomatal response to water deficit. Funct. Plant Biol. 2012, 39, 179–189. [Google Scholar] [CrossRef]
- Chaerle, L.; Van der Straeten, D. Imaging techniques and the early detection of plant stress. Trends Plant Sci. 2000, 5, 495–501. [Google Scholar] [CrossRef]
- Oerke, E.C.; Steiner, U.; Dehne, H.W.; Lindenthal, M. Thermal imaging of cucumber leaves affected by downy mildew and environmental conditions. J. Exp. Bot. 2006, 57, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Winkel, T.; Rambal, S. Influence of water stress on grapevines growing in the field: From leaf to whole-plant response. Aust. J. Plant Physiol. 1993, 20, 143–157. [Google Scholar] [CrossRef]
- Hetherington, A.M.; Woodward, I. The role of stomata in sensing and driving environmental change. Nature 2003, 424, 901–908. [Google Scholar] [CrossRef]
- Gates, D.M. Transpiration and leaf temperature. Ann. Rev. Plant Physiol. 1964, 19, 211–238. [Google Scholar] [CrossRef]
- Downton, W.J.S.; Loveys, B.R.; Grant, W.J.R. Non-uniformstomatal closure induced by water stress causes putative non-stomatal inhibition of photosynthesis. New Phytol. 1988, 110, 503–509. [Google Scholar] [CrossRef]
- Jones, H.G. Use of infrared thermometry for estimation of stomatal conductance as a possible aid to irrigation scheduling. Agric. For. Meteorol. 1999, 95, 139–149. [Google Scholar] [CrossRef]
- Jones, H.G. Plants and Microclimate, 2th ed.; Cambridge University Press: Cambridge, UK, 1992; p. 428. [Google Scholar]
- Guilioni, L.; Jones, H.G.; Leinonen, I.; Lhomme, J.P. On the relationships between stomatal resistance and leaf temperatures in thermography. Agric. Meteorol. 2008, 148, 1908–1912. [Google Scholar] [CrossRef]
- Idso, S.B.; Roselyne, I.; Abutaleb, K.; Ahmed, F. Non-water-stressed baselines: A key to measuring and interpreting plant water stress. Agric. Meteorol. 1982, 27, 59–70. [Google Scholar] [CrossRef]
- Leinonen, I.; Jones, H.G. Combining thermal and visible imagery for estimating canopy temperature and identifying plant stress. J. Exp. Bot. 2004, 55, 1423–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zia, S.; Spohrer, K.; Merkt, N.; Wenyong, D.; Xiongkui, H.; Muller, J. Non invasive water status detection in grapevine. Int. J. Agric. Biol. Eng. 2009, 2, 46–54. [Google Scholar]
- Costa, J.M.; Grant, O.M.; Chaves, M.M. Use of thermal imaging in viticulture: Current application and future prospects. In Methodologies and Results in Grapevine Research; Delrot, S., Medrano, H., Or, E., Bavaresco, L., Grando, S., Eds.; Springer: Berlin, Germany, 2010; pp. 135–150. [Google Scholar]
- Pou, A.; Diago, M.P.; Medrano, H.; Baluja, J.; Tardaguila, J. Validation of thermal indices for water status identification in grapevine. Agric. Water Manag. 2014, 134, 60–72. [Google Scholar] [CrossRef]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 2003, 26, 1393–1405. [Google Scholar] [CrossRef]
- Vandeleur, R.K.; Mayo, G.; Shelden, M.C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: Diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol. 2009, 149, 445–460. [Google Scholar] [CrossRef] [Green Version]
- Scholander, P.F.; Hammel, H.T.; Hemmingsen, E.A.; Bradstreet, E.D. Hydrostschultzatic pressure and osmotic potential in leaves of mangroves and some other plants. Proc. Natl. Acad. Sci. USA 1964, 52, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Begg, J.E.; Turner, N.C. Water potential gradients in field tobacco. Plant Physiol. 1970, 46, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Jones, H.G. Application of thermal imaging and infrared sensing in plant physiology and ecophysiology. Adv. Bot. Res. 2004, 41, 107–163. [Google Scholar]
- Qiu, G.Y.; Momi, K.; Yano, T. Estimation of plant transpiration by imitation leaf temperature (I). Theoretical consideration and field verification. Trans. Jpn. Soc. Irrig. 1996, 63, 401–410. [Google Scholar]
- Jones, H.G.; Aikman, D.; McBurney, T.A. Improvements to infrared thermometry for irrigation scheduling in humid climates. Acta Hortic. 1997, 449, 259–266. [Google Scholar] [CrossRef]
- Williams, L.E.; Arujo, F.J. Correlations among predawn leaf, midday leaf, and midday stem water potential and their correlations with other measures of soil and plant water status in Vitis vinifera. J. Am. Soc. Hortic. Sci. 2002, 127, 448–454. [Google Scholar] [CrossRef] [Green Version]
- Deloire, A.; Heyns, D. The Leaf Water Potentials: Principles, Method and Thresholds. Available online: https://www.researchgate.net/publication/259589941 (accessed on 20 December 2011).
- Flexas, J.; Escalona, J.M.; Evain, S.; Gulías, J.; Moya, I.; Osmond, C.B.; Medrano, H. Steady-state chlorophyll fluorescence (Fs) measurements as a tool to follow variations of net CO2 assimilation and stomatal conductance during water-stress in C3 plants. Physiol. Plant. 2002, 114, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardieu, F.; Simonneau, T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Lovisolo, C.; Perrone, I.A.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non-hydraulic interactions at the whole-plant level: A physiological and molecular update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- Soar, C.J.; Speirs, J.; Maffei, S.M.; Penrose, A.B.; McCarthy, M.G.; Loveys, B.R. Grape vine arieties Shiraz and Grenache differ in their stomatal response to VPD: Apparent links with ABA physiology and gene expression in leaf tissue. Aust. J. Grape Wine Res. 2006, 12, 2–12. [Google Scholar] [CrossRef]
- Williams, L.E.; Baeza, P. Relationships among ambient temperature and vapor pressure deficit and leaf and stem water potentials of fully irrigated, field-grown grapevines. Am. J. Enol. Vitic. 2007, 58, 173–181. [Google Scholar]
- Shellie, K.; Glenn, D.M. Wine grape response to kaolin particle film under deficit and well-watered conditions. Acta Hortic. 2008, 792, 587–591. [Google Scholar] [CrossRef]
- Zia, S.; Wenyong, D.; Spreer, W.; Spohrer, K.; Xiongkui, H.; Müller, J. Assessing crop water stress of winter wheat by thermography under different irrigation regimes in North China Plain. Int. J. Agric. Biol. Eng. 2012, 5, 24–34. [Google Scholar]
- Leinonen, I.; Grant, O.M.; Tagliavia, C.P.P.; Chaves, M.M.; Jones, H.G. Estimating stomatal conductance with thermal imagery. Plant Cell Environ. 2006, 29, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Moller, M.; Alchanatis, V.; Cohen, Y.; Meron, M.; Tsipris, J.; Naor, A.; Ostrovsky, V.; Sprintsin, M.; Cohen, S. Use of thermal and visible imagery for estimating crop water status of irrigated grapevine. J. Exp. Bot. 2006, 58, 827–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, Y.; Kimball, B.A.; Jackson, R.D.; Pinter, P.J.; Reginato, R.J. Remote estimation of leaf transpiration rate and stomatal resistance based on infrared thermometry. Agric. For. Meteorol. 1990, 51, 21–33. [Google Scholar] [CrossRef]
- Inoue, Y.; Sakuratani, T.; Shibayama, M.; Morinaga, S. Remote and real-time sensing of canopy transpiration and conductance-comparison of remote and stem flow gauge methods in soybean canopies as affected by soil water status. Jpn. J. Crop Sci. 1994, 63, 664–670. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | DOY | Solar Radiation (W m−2) | Relative Humidity (%) | Wind Speed (km h−1) | Average Air Temperature (°C) | VPD (KPa) |
|---|---|---|---|---|---|---|
| Merlot | 206 | 831 | 31 | 4.5 | 27.5 | 2.5 |
| Merlot | 207 | 731 | 29 | 4.5 | 24.6 | 1.3 |
| Merlot | 208 | 851 | 42 | 5.0 | 26.8 | 2.1 |
| Moscato | 211 | 772 | 33 | 4.9 | 29.0 | 2.5 |
| Moscato | 213 | 820 | 34 | 7.0 | 29.9 | 2.8 |
| Moscato | 214 | 795 | 32 | 5.0 | 27.7 | 2.7 |
| Merlot (gs) (mmol H2O m−2 s−1) | Moscato (gs) (mmol H2O m−2 s−1) | |||||
|---|---|---|---|---|---|---|
| Treatment | DOY 206 | DOY 207 | DOY 208 | DOY 211 | DOY 213 | DOY 214 |
| T0 | 198.1 a | 232.2 a | 204.6 a | 286.7 a | 258.6 a | 219.2 a |
| T1 | 52.7 b | 81.3 b | 76.8 b | 77.8 b | 71.1 b | 73.8 b |
| T2 | 31.3 c | 48.1 c | 48.3 c | 47.6 c | 49.3 c | 31.6 c |
| Merlot | Moscato | ||||||
|---|---|---|---|---|---|---|---|
| Canopy Portion | Target | DOY 206 | DOY 207 | DOY 208 | DOY 211 | DOY 213 | DOY 214 |
| Sunlit | Tdry | 40.5 a | 36.3 a | 38.7 a | 39.7 a | 40.3 a | 40.3 a |
| Twet | 26.8 b | 25.6 b | 26.8 b | 29.2 b | 29.7 b | 29.4 b | |
| Shaded | Tdry | 34.1 a | 33.9 a | 33.6 a | 36.1 a | 36.1 a | 34.7 a |
| Twet | 25.9 b | 24.4 b | 23.9 b | 28.3 b | 28.3 b | 27.8 b | |
| Merlot | DOY 206 | DOY 207 | DOY 208 | |||
| Treatment | Sunlit | Shaded | Sunlit | Shaded | Sunlit | Shaded |
| T0 | 0.5 b | 0.5 b | 0.2 c | 0.4 c | 0.4 c | 0.5 c |
| T1 | 0.8 a | 1.0 a | 0.6 b | 0.6 b | 0.6 b | 0.7 b |
| T2 | 0.8 a | 1.1 a | 0.9 a | 0.8 a | 0.8 a | 1.0 a |
| Moscato | DOY 211 | DOY 213 | DOY 214 | |||
| Treatment | Sunlit | Shaded | Sunlit | Shaded | Sunlit | Shaded |
| T0 | 0.3 b | 0.4 c | 0.3 c | 0.5 c | 0.3 c | 0.5 c |
| T1 | 0.8 a | 0.8 b | 0.7 b | 0.7 b | 0.7 b | 0.7 b |
| T2 | 0.9 a | 1.1 a | 0.9 a | 1.0 a | 0.9 a | 0.9 a |
| Merlot | DOY 206 | DOY 207 | DOY 208 | |||
| Treatment | Sunlit | Shaded | Sunlit | Shaded | Sunlit | Shaded |
| T0 | 0.8 a | 1.0 a | 3.5 a | 1.2 a | 1.6 a | 0.9 a |
| T1 | 0.3 b | 0.4 b | 0.7 b | 0.6 b | 0.5 b | 0.3 b |
| T2 | 0.3 b | 0.01 c | 0.2 c | 0.3 c | 0.4 b | 0.01 c |
| Moscato | DOY 211 | DOY 213 | DOY 214 | |||
| Treatment | Sunlit | Shaded | Sunlit | Shaded | Sunlit | Shaded |
| T0 | 2.7 a | 1.3 a | 1.6 a | 1.2 a | 2.0 a | 0.9 a |
| T1 | 0.5 b | 0.4 b | 0.5 b | 0.4 b | 0.5 b | 0.4 b |
| T2 | 0.2 c | 0.04 c | 0.08 c | 0.04 c | 0.1 c | 0.1 c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belfiore, N.; Vinti, R.; Lovat, L.; Chitarra, W.; Tomasi, D.; de Bei, R.; Meggio, F.; Gaiotti, F. Infrared Thermography to Estimate Vine Water Status: Optimizing Canopy Measurements and Thermal Indices for the Varieties Merlot and Moscato in Northern Italy. Agronomy 2019, 9, 821. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120821
Belfiore N, Vinti R, Lovat L, Chitarra W, Tomasi D, de Bei R, Meggio F, Gaiotti F. Infrared Thermography to Estimate Vine Water Status: Optimizing Canopy Measurements and Thermal Indices for the Varieties Merlot and Moscato in Northern Italy. Agronomy. 2019; 9(12):821. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120821
Chicago/Turabian StyleBelfiore, Nicola, Rosanna Vinti, Lorenzo Lovat, Walter Chitarra, Diego Tomasi, Roberta de Bei, Franco Meggio, and Federica Gaiotti. 2019. "Infrared Thermography to Estimate Vine Water Status: Optimizing Canopy Measurements and Thermal Indices for the Varieties Merlot and Moscato in Northern Italy" Agronomy 9, no. 12: 821. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9120821