Transcriptomic Analyses of the Hypothalamic-Pituitary-Gonadal Axis Identify Candidate Genes Related to Egg Production in Xinjiang Yili Geese

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Feed and Feeding Management
2.3. Total RNA Extraction
2.4. Construction of cDNA Libraries and Illumina Sequencing
2.5. Sequencing Data Analyses
2.6. Real-Time PCR Validation of Sequencing Results
3. Results
3.1. Evaluation of RNA and Sequencing Data Quality
3.2. Identification of Differentially Expressed Genes
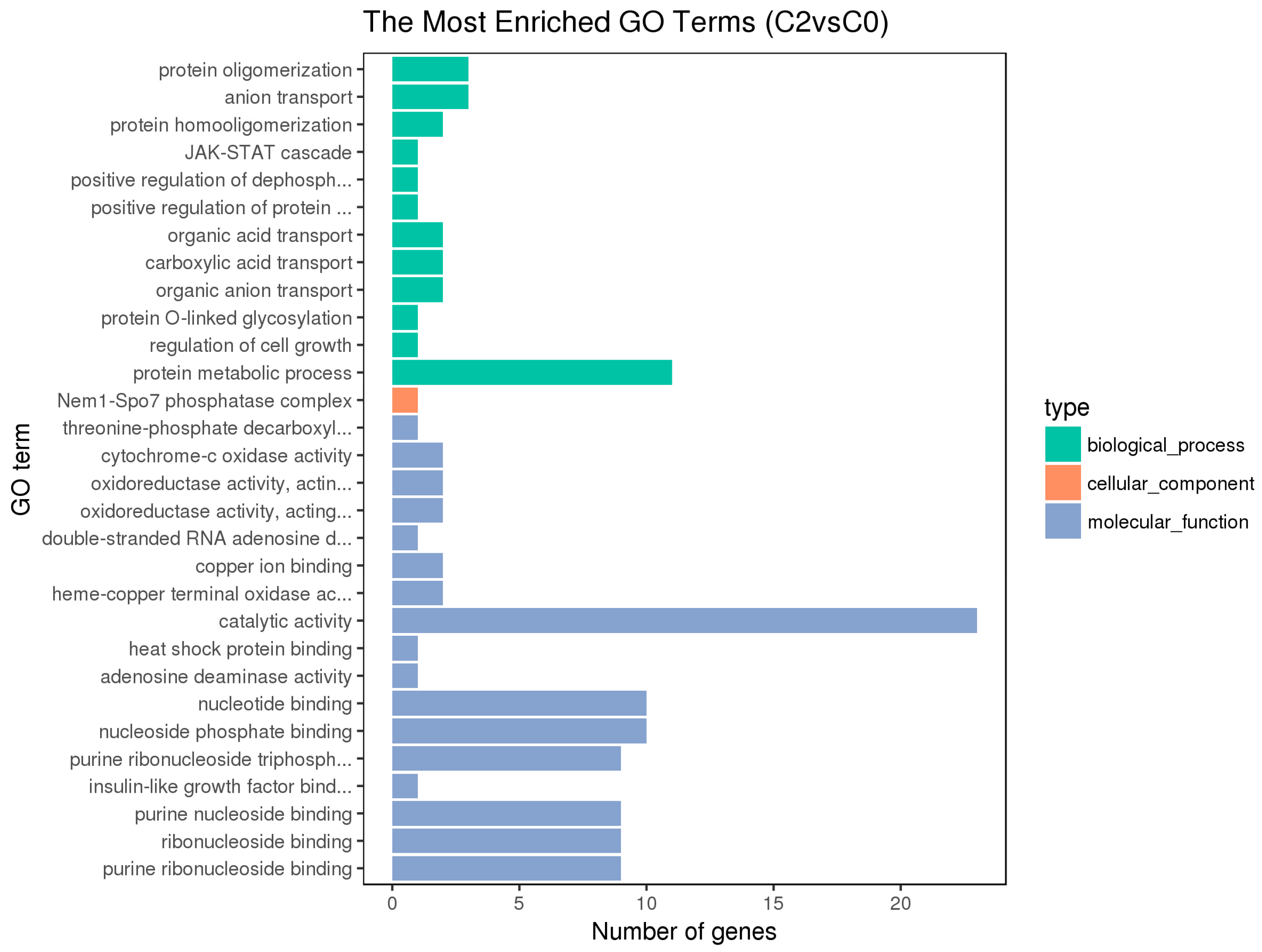
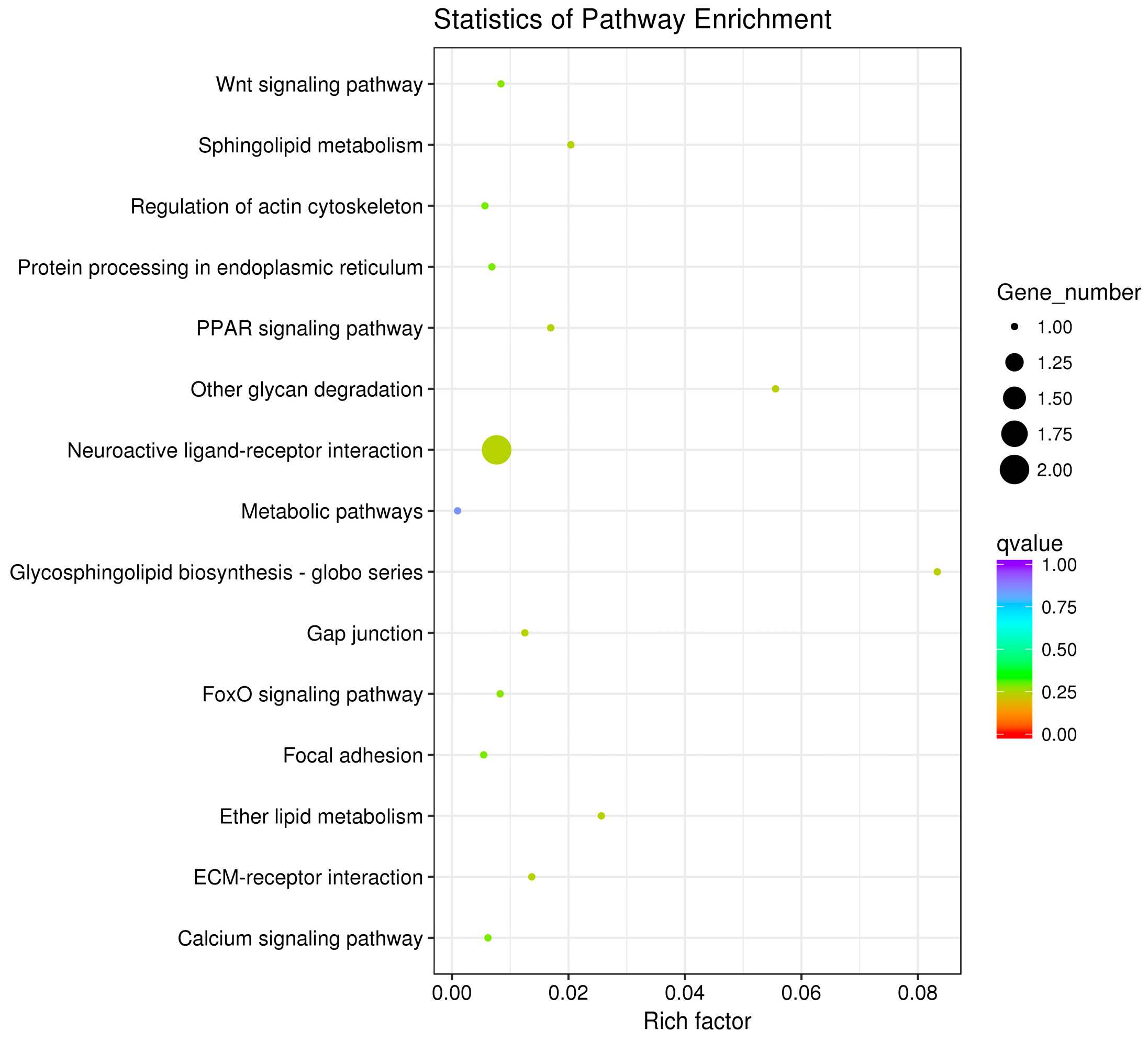
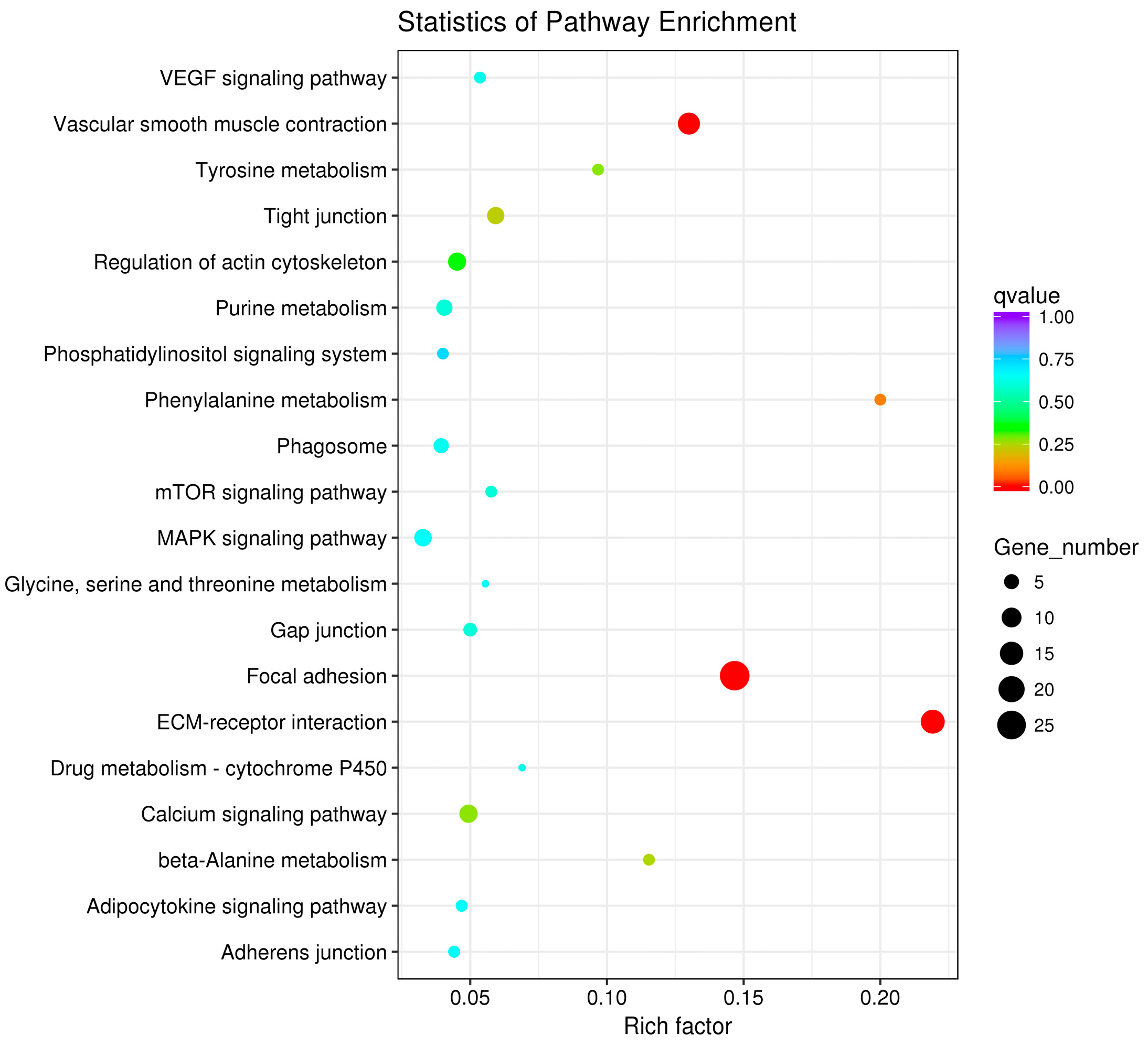
3.3. GO and KEGG Pathway Enrichment Analyses of Differentially Expressed Genes
3.4. Fluorescence Quantitative Polymerase Chain Reaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fanming, Z.; Dugan, N. Variety Characteristics of Yili Goose and Grassland Feeding and Management Techniques. China Anim. Health 2015, 17, 6–8. [Google Scholar]
- Huifang, L.; Weitao, S.; Wenjuan, X.; Wenqi, Z.; Qingping, T.; Kuanwei, C. Study on Genetic Structure and Differentiation of Ten Goose Breeds in China. Acta Ecol. Anim. Domast. 2009, 30, 10–14. [Google Scholar]
- Daixian, D.; Gang, Y.; Bing, W. Variety Characters and Development Prospect of Yili Goose. Xinjiang Agric. Sci. 1992, 1, 42–43. [Google Scholar]
- Xianxiao, B.; Zijing, Z.; Jing, W.; Zhaoxue, X. Application of Transcriptome Sequencing Technology in Genetic Breeding of Livestock. Henan Agric. Sci. 2017, 46, 6–9. [Google Scholar]
- Chunlan, Z.; Zijuan, Q.; Guizhi, W.; Zhibin, J.; Jianmin, W. Transcriptome and RNA-Seq Technology. Biotechnol. Bull. 2012, 12, 57–62. [Google Scholar]
- Huang, W.; Khatib, H. Comparison of transcriptiome assembly of RNA-Seq data in a sheep model for delayed hone healing. BMC Genom. 2011, 12, 158. [Google Scholar]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.K.; Mockler, T.C. Genome-wide mapping of alternative splicing in Arabidops thaliana. Genom. Res. 2010, 20, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.K.; Wang, X.W.; Wang, X.; Liang, Y.; Zhang, X. Observations on novel splice junctions from RNA sequencing data. Biochem. Biophys. Res. Commun. 2011, 409, 299–303. [Google Scholar] [CrossRef]
- Maher, C.A.; Kumar-Sinha, C.; Cao, X.H.; Kalyana-Sundaram, S.; Han, B.; Jing, X.; Sam, L.; Barrette, T.; Palanisamy, N.; Chinnaiyan, A.M.; et al. Transcriptome Sequencing to detect gene fusions in cancer. Nature 2009, 458, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Ren, S.C.; Peng, Z.Y.; Mao, J.H.; Yu, Y.; Yin, C.; Gao, X.; Cui, Z.; Zhang, J.; Yi, K.; Xu, W.; et al. RNA-Seq analysis of prostate cancer in the Chinese population identifies recurrent gene fusions, cancer-associated long noneoding RNAs and aberrant alternative splicing. Cell Res. 2012, 22, 806–821. [Google Scholar] [CrossRef]
- Wei, L.; Zhong, C. Research Progress of the Main Reproductive Hormone Receptors in Poultry Ovary. China Poult. 2016, 38, 48–56. [Google Scholar]
- Liu, H.; Wang, J.; Li, L.; Han, C.; He, H.; Xu, H. Transcriptome analysis revealed the possible regulatory pathways initiating female geese broodiness within the hypothalamic-pituitary-gonadal axis. PLoS ONE 2018, 13, e0191213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, G.; Li, Q.; Zhao, X.; Ding, N.; Han, Q.; Su, J.; Wang, Q. Transcriptome profiling of the hypothalamus during prelaying and laying periods in Sichuan white geese (Anser cygnoides). Anim. Sci. J. 2015, 86, 800–805. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Han, Q.; Zhao, X.Z.; Li, Q.; Li, J.; Zhang, H.F.; Gao, G.L.; Luo, Y.; Xie, Y.H.; Su, J. Differential gene expression in pre-laying and laying period ovaries of Sichuan White geese (Anser cygnoides). GMR 2015, 14, 6773–6785. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression analysis for sequence countdata. Genom. Biol. 2010, 11, 106. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genom. Boil. 2010, 11, 14. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Liangtao, W.; Min, Z.; Min, H. Screening of Differentially Expressed Immune related Genes from Duck Spleenvwith Duck Enteritis Virus Infection Based on RNA- Seq Technology. Acta Vet. Zootech. Sin. 2017, 48, 297–306. [Google Scholar]
- Mustard, J.A.; Beggs, K.T.; Mercer, A.R. Molecular biology of the invertebrate dopamine receptors. Arch. Insect. Biochem. 2005, 59, 103–117. [Google Scholar] [CrossRef]
- Shulin, N.; Zhenyou, W.; Chenzhen, C.; Jingbo, L.; Meng, H.; Xiuwu, W. Effect of exogenous cAMP on production performance of Hailan commercial laying hens. Anim. Husb. Vet. Med. 2000, 5, 17–18. [Google Scholar]
- Youngren, O.M.; Chaiseha, Y.; Halawani, M.E. Regulation of prolactin secretion by dopamine and vasoactive instinal peptide at theievel of the pituitary in the turkey. Neuroendocriolgy 2008, 12, 319–325. [Google Scholar]
- Xu, H.; Shen, X.; Zhou, M.; Fang, M.; Zeng, H.; Nie, Q.; Zhang, X. The genetic effects of the dopa-mine D1 receptor gene on chicken egg production and broodiness traits. BMC Genet. 2010, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, S.; Li, C.; Feng, Y.; Peng, X.; Gong, Y. Molecular cloning, expression profile, polymorphism and the genetic effects of the dopamine D1 receptor gene on duck reproductive traits. Mol. Biol. Rep. 2012, 39, 9239–9246. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Shijun, L.; Chuang, L.; Yanping, F.; Xiuli, P.; Yanchang, G. Cloning and polymorphism detection of duck dopamine receptor 1 (DRD1) gene. Natl. Poult. Symp. 2011, 226–230. [Google Scholar]
- Scanes, C.G.P.; Proudman, J.A.; Radecki, S.V. Influence of continuous growth hormone insulin-like growth factor Iadministration in adult female chickens. Gen. Comp. Endocrinol. 1999, 114, 315–323. [Google Scholar] [CrossRef]
- Hao, Z.; Fang, C.; Zhenhua, L.; Jinping, D. Advances in follicular atresia in poultry. Hubei J. Anim. Vet. Sci. 2014, 35, 61–65. [Google Scholar]
- Jia, Y.D.; Yan, F.F.; Zeng, W.D.; Zhang, C.Q. Promoting Effect of IGF-I on Prehierarchical Follicle Development in Laying Chickens. Sci. Agric. Sin. 2011, 44, 4295–4301. [Google Scholar]
- Zhu, W.; Li, H.; Wu, X.; Qian, K.; Zhang, J.; Ge, H. The genetic effects of N PY and IGF-1 genes on reproduction traits of Wenchang chicken. J. Yangzhou Univ. (Agric. Life Sci. Ed.) 2007, 02, 20–23. [Google Scholar]
- Mingyue, Y. Research on Gene Markers Associated with the Egg Production Traits of Shiqiza Chicken; Nanjing Normal University: Nanjing, China, 2006. [Google Scholar]
- Tong, T.; Shen, Y.; Lee, H.W.; Yu, R.; Park, T. Adenylyl cyclase 3 haploinsufficiency confers susceptibility to dietinduced obesity and insulin resistance in mice. Sci. Rep. 2016, 6, 34179. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.B.; Robert, J.R. Interactions of gonadotropins with corpus luteum membranes. I. Properties and distributions of same marker enzyme activities after subcellular fractionation of the superovulated rat ovary. Endocrinology 1978, 103, 778. [Google Scholar]
- Lee, C.Y.; Tateishi, K.; Ryan, R.J.; Jiang, N.S. Binding of human chorionic gonadotropin by rat ovarian slices: Dependance on the functional state of the ovary. Proc. Soc. Exp. Biol. Med. 1975, 1148, 505. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Guan, Y.; Zhang, W.; Wu, Y.; Jia, H.; Zhang, Y.; Zhang, X.; Du, H.; Wang, X. RNA-seq analysis of the hypothalamic transcriptome reveals the networks regulating physiopathological progress in the diabetic GK rat. Sci. Rep. 2016, 6, 34138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Han, S.; Shu, L.; Zhang, J.; Liu, M.; Shen, L.; Wang, Z. Adenylyl Cyclase 3 Deletion Influences the Related Factors andSignaling Pathways in the Main Olfactory Epithelium of Mice. Chin. J. Biochem. Mol. Biol. 2016, 32, 816–822. [Google Scholar]
- Livera, G.; Xie, F.; Garcia, M.A.; Jaiswal, B.; Chen, J.; Law, E.; Storm, D.R. Conti M. Inactivation of the mouse adenylyl cyclase 3 gene disrupts male fertility and spermatozoon function. Mol. Endocrinol. 2005, 19, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Heng, L. Studies on Expression Characteristics and Genetic Effects of Poultry Reproductive Genes; Yangzhou University: Yangzhou, China, 2011. [Google Scholar]
- Liu, Y.; Wang, D.K.; Chen, L.M. The physiology of bicarbonate transporters in mammalian reproduction. Biol. Reprod. 2012, 86, 1–13. [Google Scholar] [CrossRef]
- Yin, L.; Qunwei, L.; Liming, C. Physiology and pathophysiology of Na+/HCO3 cotransporter NBCe1. Acta Physiol. Sin. 2012, 64, 729–740. [Google Scholar]
- Majumdar, D.; Maunsbach, A.B.; Shacka, J.J.; Williams, J.B.; Berger, U.V.; Schultz, K.P.; Harkins, L.E.; Boron, W.F.; Roth, K.A.; Bevensee, M.O. Localization of electrogenic Na/bicarbonate cotransporter NBCe1 variants in rat brain. Neuroscience 2008, 155, 818–832. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, J.Y.; Wang, D.K.; Wang, L.; Chen, L.M. Cloning and identification of two novel NBCe1 splice variants from mouse reproductive tract tissues: A comparative study of NCBT genes. Genomics 2011, 98, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.; Schmitt, B.M.; Brown, D.; Berger, U.V.; Hediger, M.A.; Boron, W.F.; Breton, S. Localizati on of sodi umn bicarbonate co-transporter (NB C) protein and mRNA in rat epi didymis. Biol. Reprod. 1999, 60, 573–579. [Google Scholar] [CrossRef] [Green Version]
- Gholami, K.; Muniandy, S.; Salleh, N. Modulation of sodium–bicarbonate co-transporter (SLC4A4/NBCe1) protein and mRNA expression in rat’s uteri by sex-steroids and at different phases of the oestrous cycle. Res. Vet. Sci. 2014, 96, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Yu, M.K.; Leung, K.M.; Yip, C.Y.; Ko, W.H.; Liu, C.Q.; Chan, HC. Involvement of Na+-HCO3–cotransporter in medi-ating cyclic adenosine 3’, 5-monophosphate-dependent HCO3–secretion by mouse endometrial epithelium. Biol. Reprod. 2002, 66, 1846–1852. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Ba, C.; Zhang, Y.; Zhao, X. Progress on Transcription Factors on the Regulation of Expression of Melanocortin2-repeptor Gene. Chinese J. Comp. Med. 2007, 01, 46–50. [Google Scholar]
- Kai, C.; Jun, X.; Liguo, L.; Binwen, X. Advances in research on melanocortin 2 receptors. Jiangsu Agric. Sci. 2015, 43, 16–20. [Google Scholar]
- Roy, S.; Perron, B.; Gallo-Payet, N. Role of asparagine-linked glycosylation in cell surface expression and function of the human adrenocorticotropin receptor (melanocortin 2 receptor) in 293/FRT cells. Endocrinology 2010, 151, 660–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S.; RoyS, J.; Pinard, S.; Agulleiro, M.J.; Cerdá-Reverter, J.M.; Parent, J.L.; Gallo-Payet, N. The C-terminal domains of melanocortin-2 receptor (MC2R) accessory proteins (MRAP1) influencetheir localization and ACTH induced cA MP prodduction. Gen. Comp. Endocrinol. 2012, 176, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, A.A.J.; Lee, A.A.; Sebag, J.A. Regions of MRAP2 required for the inhibition of orexin and prokineticin receptor signaling. BBA Mol. Cell Res. 2017, 1864, 2322–2329. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | X04 | C04 | L04 | X23 | C23 | L23 |
|---|---|---|---|---|---|---|
| Total reads | 52,544,012 | 47,471,474 | 43,311,034 | 49,243,474 | 43,885,970 | 44,825,988 |
| Total mapped | 40,355,619 (76.8%) | 36,619,486 (77.14%) | 31,406,234 (72.51%) | 38,401,158 (77.98%) | 33,704,381 (76.8%) | 33,641,532 (75.05%) |
| Multiple mapped | 388,703 (0.74%) | 349,245 (0.74%) | 620,638 (1.43%) | 374,387 (0.76%) | 313,471 (0.71%) | 640,850 (1.43%) |
| Uniquely mapped | 39,966,916 (76.06%) | 36,270,241 (76.4%) | 30,785,596 (71.08%) | 38,026,771 (77.22%) | 33,390,910 (76.09%) | 33,000,682 (73.62%) |
| Organ | DEG | |
|---|---|---|
| Hypothalamus | Up-regulated | SFXN1, SGMS1, THAP5, SPRY1, COG2, LOC106031011, CARHSP1, Novel02312, LOC106031299, DMAP1, LOC106030886, MTERF4, SNRNP35, LOC106036439, Novel00604, Novel00813, LOC106048860, LOC106033299, PMP22, LOC106038373, CENPP, LOC106043368, TMEM255B, Novel02772, LOC106034340, LOC106032431, RBP1, STOML3, Novel02183, Novel00159, GRAP2, Novel00353, LOC106035108, CPS1, ANKRD1, Novel03069, IGFBP1, LOC106033972, LOC106035461, SPTBN5, FAM167B, Novel01797, GALNT8, LOC106037026, Novel00680, Novel00440, GPR55, Novel02518, LOC106044451, Novel02028, KIF18B, Novel02623, LOC106039706, ARMC12, LOC106046673, LOC106035746, SLC22A18, Novel00332, Novel01473, Novel02483, LOC106037068, LOC106049124, AGR2, Novel00634, Novel00776, Novel02517, LOC106030701, LOC106034203, LOC106032232, LOC106046797, LOC106038395, LOC106033669, LOC106039869, Novel00497, LOC106047183, Novel00809, LOC106047182, LOC106049953, LOC106046796. |
| Down-regulated | LOC106043121, Novel00149, LOC106048725, NR4A3, EGR4, Novel00772, LOC106049845, SLA, LOC106033075, LOC106043342, 106032369, LOC106029709, Novel02265, Novel02348, LOC106036764, RUNX3, Novel01461, Novel00460, LOC106037894, Novel00595, BARD1, Novel02665, ANTXR1, Novel03067, OTOL1, LOC106032087, C1QTNF3, Novel01396, DDR2, MXRA5, LOC106049821, LOC106036261, LOC106039154, LOC106045306, FAM227B, LOC106040733, Novel01364, Novel02473, RFWD3, CGN, FILIP1L, CEMIP, LOC106032402, F13A1, Novel00314, SCRT2, LOC106033600, RNF213, LOC106032129, Novel02666, AQP9, Novel01931, LOC106042177, MAP7D3, NTRK3, TIE1. | |
| Pituitary | Up-regulated | LOC106031011, CRACR2B, SOX9, LOC106036438, LOC106036439, LOC106036974, TNC, NSMF, LOC106037323, ATP13A3, LOC106039850, DNAJA1, PRKG2, SPARCL1, P2RY2, LOC106042825, SLC4A4, TMEM201, SFRP4, LOC106044451, LOC106049066, Novel00497, Novel01686, Novel02339, Novel02920. |
| Down-regulated | VIT, SLC15A2, LOC106031282, LOC106031667, FABP7, DRD5, NDE1, LOC106034326, AFAP1L1, LOC106034774, KCTD4, ENPP6, CTHRC1, LOC106036254, LOC106036743, SCIN, LOC106037776, SYNPO2, LOC106038998, NEU4, LOC106039845, LOC106042210, STK17A, MRAP, DNAJC28, SGK2, SPTSSB, SLAIN1, GBGT1, LOC106047046, LOC106049843. | |
| Ovary | Up-regulated | LOC106029456, LOC106029515, LOC106029546, LOC106029652, LOC106029721, LOC106029781, SPEG, MYLK, HEG1, LOC106029988, SORBS1, LOC106030002, HOXA3, HOXA5, PRKG1, NR1D2, LRRC3B, LOC106030346, LOC106030347, CD109, KCNQ5, RIMS1, PSD, BAG2, LOC106030416, GEM, LOC106030538, PI15, COLEC11, GABRA2, RERG, LOC106030915, MICAL3, KIAA1671, PROX1, CDC42BPA, ACTN1, MEIS2, CACNA1H, MPRIP, LOC106032203, RASD1, KIAA0922, NPY2R, LOC106032455, LAMA2, FAM26E, LAMA4, ZCCHC24, PPP2R2B, KCNMA1, RASGEF1C, ADAMTS2, ACOT11, CDKN2C, ROR1, MAST2, ZSWIM5, VCL, PRKAA2, LOC106033331, RGS11, MYH11, CPN1, EEF2K, PRKAR1B, FBXL22, HIPK3, SYDE2, ADGRL4, KCNMB1, LOC106034392, LOC106034406, COL6A3, COL3A1, PRKCA, AFAP1, EMILIN2, LOC106035048, DGKH, SLC25A4, NCALD, SYBU, LOC106035506, SEMA3C, MAST4, PDE4D, RAB3C, INPP4A, RBFOX2, ALDH1L2, IGF1, PVALB, LOC106035922, GALR1, EML1, KCNE4, ADAMTS12, TESK1, RAI14, TPM2, RXFP3, LOC106036401, SYNM, NR2F2, LOC106036475, RGMA, ALDH1A3, CILP, RASL12, LOC106036832, LOC106036842, ZBTB47, LOC106037166, AKAP5, ADSSL1, RHOJ, DACT1, SETBP1, POPDC2, ARL13A, IL13RA1, PON2, GREB1L, LOC106037870, SYNC, LOC106037900, LDLRAP1, MGLL, ADCY3, ABCC9, LOC106038240, KCNJ8, LIMD1, FYCO1, ZHX1, LOC106038471, LOC106038505, IRS4, CHRDL1, ABI3BP, COL4A6, COL4A5, CAPN6, SYNPO2, TGFBR3, CDK6, CLMP, GAB2, LOC106039897, SDPR, LOC106040026, LOC106040036, SLC6A1, FBLN2, LACTBL1, LIMS2, GPR17, MYO7B, LOC106040291, CRYL1, LOC106040422, ADA, JPH2, JAM3, ADAMTS8, MORN5, LOC106040733, SCUBE2, NRIP3, MICAL2, MRVI1, HSPB1, LOC106041406, MYO1C, P2RX1, FMO3, PRRX1, RGS5, LOC106041605, FHL2, CAP2, SUSD5, LAMB1, CAV1, LOC106042166, GPR20, CCDC102B, SLC24A3, LOC106042361, ARHGEF17, CHRDL2, RGS6, ANGPTL1, ZBTB37, ZEB1, KIAA1462, LOC106042668, PDLIM7, NEURL1B, DRD1, LOC106042807, MUSTN1, STARD9, INHBA, PGM5, SPTBN5, GNAO1, LOC106043213, LOC106043342, CEMIP, MRAP, LOC106043529, PDE4B, RAVER2, TEK, PITPNM3, MN1, SLC4A4, FERMT2, LOC106044084, ADAMTSL2, CSPG4, PEAK1, LINGO1, LOC106044229, KIF1B, ACTA2, ADGRA2, COL4A1, COL4A2, LOC106044472, LOC106044704, MEIS1, MUC4, MASP1, CDH13, LOC106045192, OTOGL, MYL9, LOC106045455, MED13L, LOC106045561, LOC106045776, LOC106045852, LOC106045870, CSRP1, TNS1, MYL4, LOC106046005, IP6K3, TEAD3, LMOD1, PRDM6, LOC106046391, PPP1R12A, 106046568, LOC106046709, TRHDE, LOC106047053, MSRB3, MYOCD, NCS1, PRR16, SMTN, CCDC50, UTS2B, ACTG2, SCNN1A, PIANP, LOC106048001, LOC106048002, PRSS23, MFAP5, CPT1B, HOXB3, THBS3, DNAJB5, IL11RA, HSPG2, KCNK1, ACTA1, NID1, LOC106048884, FLNC, NPR1, ECM2, OGN, MYO18A, COL6A1, PCBP3, COL6A2, GLI1, Novel00206, Novel00225, Novel00283, Novel00349, Novel00428, Novel00498, Novel00508, Novel00568, Novel00676, Novel00688, Novel00911, Novel01277, Novel01584, Novel01601, Novel02245, Novel02466, Novel02966, Novel03036, Novel03063. |
| Down-regulated | KCNB2, SAMD3, FSTL4, LOC106033036, PRRG4, CA10, LOC106035557, LOC106036685, LOC106042047, LOC106042103, TSPAN8, LOC106046958, LOC106048733, LOC106048993, GAL3ST1, Novel00233, Novel02085, Novel02711, Novel02751. |
| Terms/Pathways | Gene Name | log2FoldChange | Padj | Up-Down Regulation (HEP/LEP) |
|---|---|---|---|---|
| steroid biosynthetic process | LOC106035461 | 1.247 | 0.73322 | Up |
| steroid hormone-mediated signaling pathway | NR4A3 | −1.5699 | 0.34746 | Down |
| developmental process | LOC106040733 | −1.2066 | 0.71683 | Down |
| reproduction | Novel01396 | −1.2601 | 0.72516 | Down |
| G-protein coupled receptor signaling pathway | GPR55 | 1.3013 | 0.7477 | Up |
| growth | LOC106034326 | −1.0064 | 0.028997 | Down |
| G-protein coupled receptor signaling pathway | P2RY2 | 1.7694 | 0.031867 | Up |
| DRD5 | −1.8187 | 0.049623 | Down | |
| calcium ion binding | CRACR2B | 1.7139 | 0.0090239 | Up |
| SPARCL1 | 2.7338 | 0.012128 | Up | |
| reproduction | HOXB3 | 1.1354 | 0.028618 | Up |
| DRD1 | 1.9867 | 0.028259 | Up | |
| developmental process | LAMA2 | 1.8842 | 0.0060556 | Up |
| LAMA4 | 1.4628 | 0.0014504 | Up | |
| COL4A1 | 2.388 | 0.039715 | Up | |
| COL4A2 | 2.2348 | 0.033559 | Up | |
| COL4A5 | 1.0104 | 0.018881 | Up | |
| P2RX1 | 2.6862 | 0.0054992 | Up | |
| ADCY3 | 1.6102 | 0.0060191 | Up | |
| LOC106040733 | 1.3839 | 0.026521 | Up | |
| Neuroactive ligand-receptor interaction | DRD5 | −1.8187 | 0.049623 | Down |
| P2RY2 | 1.7694 | 0.031867 | Up | |
| Sphingolipid metabolism | NEU4 | −2.237 | 0.0000000023607 | Down |
| Focal adhesion | IGF1 | 3.0141 | 0.00029768 | Up |
| Oocyte meiosis | ||||
| Progesterone-mediated oocyte maturation | ||||
| mTOR signaling pathway | ||||
| FoxO signaling pathway | ||||
| p53 signaling pathway | ||||
| Vascular smooth muscle contraction | PRKCA | 1.0063 | 0.023648 | Up |
| Calcium signaling pathway | ||||
| MAPK signaling pathway | ||||
| Tight junction | ||||
| Gap junction | ||||
| mTOR signaling pathway | ||||
| VEGF signaling pathway | ||||
| Phosphatidylinositol signaling system | ||||
| Focal adhesion | ||||
| Melanogenesis | ||||
| Vascular smooth muscle contraction | ADCY3 | 1.6102 | 0.0060191 | Up |
| Calcium signaling pathway | ||||
| GnRH signaling pathway | ||||
| Oocyte meiosis | ||||
| Progesterone-mediated oocyte maturation | ||||
| Tight junction | ||||
| Gap junction | ||||
| Purine metabolism | ||||
| Adrenergic signaling in cardiomyocytes | ||||
| Melanogenesis | ||||
| Gap junction | PRKG1 | 1.3621 | 0.0018268 | Up |
| Focal adhesion | THBS3 | 2.4123 | 0.000087642 | Up |
| ECM-receptor interaction | ||||
| Phagosome | ||||
| Vascular smooth muscle contraction | MYLK | 2.8068 | 0.00081465 | Up |
| Calcium signaling pathway | ||||
| Tight junction | ||||
| Focal adhesion | ||||
| Regulation of actin cytoskeleton | ||||
| Focal adhesion | MYL9 | 3.1427 | 0.0018211 | Up |
| Vascular smooth muscle contraction | ||||
| Regulation of actin cytoskeleton | ||||
| Calcium signaling pathway | LOC106045455 | 2.4292 | 0.00036177 | Up |
| Tight junction | ||||
| Tight junction | SLC25A4 | 2.7157 | 0.00087784 | Up |
| Calcium signaling pathway | ||||
| Calcium signaling pathway | CACNA1H | 1.4128 | 0.0014504 | Up |
| MAPK signaling pathway | ||||
| Tight junction |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhao, X.; Chen, L.; Wang, J.; Duan, Y.; Li, H.; Lu, L. Transcriptomic Analyses of the Hypothalamic-Pituitary-Gonadal Axis Identify Candidate Genes Related to Egg Production in Xinjiang Yili Geese. Animals 2020, 10, 90. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10010090
Wu Y, Zhao X, Chen L, Wang J, Duan Y, Li H, Lu L. Transcriptomic Analyses of the Hypothalamic-Pituitary-Gonadal Axis Identify Candidate Genes Related to Egg Production in Xinjiang Yili Geese. Animals. 2020; 10(1):90. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10010090
Chicago/Turabian StyleWu, Yingping, Xiaoyu Zhao, Li Chen, Junhua Wang, Yuqing Duan, Haiying Li, and Lizhi Lu. 2020. "Transcriptomic Analyses of the Hypothalamic-Pituitary-Gonadal Axis Identify Candidate Genes Related to Egg Production in Xinjiang Yili Geese" Animals 10, no. 1: 90. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10010090