1. Introduction
Staphylococcus aureus is a widely distributed bacterium in nature and is often considered a frequent host of skin [
1] and mucous, mainly in the nasal cavity [
2]. Studies have shown that about 20% of humans are persistent nasal carriers of
S. aureus, and around 30% are intermittent carriers [
2]. The percentage of nasal carriers reported in different animal species varies: 7.9% in horses [
3], 29% in ewes [
4] and 32.1–53% in rabbits [
5,
6]. Asymptomatic nasal carriers play a key role in the epidemiology and control of staphylococcal diseases, as the nasal cavity allows bacteria to persist over time and to multiply, which constitutes a source of infection [
2]. In human medicine, the main problem caused by
S. aureus occurs in hospitals, with the most important cause of not only nosocomial infections [
7], but also community-acquired infection [
8].
S. aureus is also a major pathogen in veterinary medicine that affects various animal species. In commercial rabbits, it has been signaled as one of the main causes of culling on farms [
9,
10]. In these cases, staphylococcal infections in rabbitries are caused by the international dissemination of the ST121 lineage of
S. aureus, and other less frequent lineages like ST96 [
11,
12].
The increase in bacterial resistance to antibiotics in recent years has become a serious health problem.
S. aureus is perfectly capable of acquiring multiple resistance mechanisms to several antimicrobial agents [
13,
14], which limits their therapeutic effectiveness. One of the most important ones for its clinical repercussions is resistance to methicillin. Methicillin-resistant
Staphylococcus aureus (MRSA) evolved from Methicillin-susceptible
Staphylococcus aureus (MSSA) by acquiring SSC
mec elements containing a
mec gene (
mecA,
mecC), which encodes a protein with a low affinity for β-lactam antibiotics [
15].
In animals, livestock-associated MRSA (LA-MRSA) emerged in pigs in 2005 [
16] and was later described in other animal species. The most widely related clonal complex (CC) to LA-MRSA is CC398. This CC was also isolated from farmers who had been in close contact with infected animals [
17], which indicates that this clone plays an important role as a reservoir of transmission to humans [
18]. Therefore, special attention has been paid to the colonization of animals with
S. aureus because they may potentially act as a reservoir to humans [
15,
19,
20].
Rabbits are among the animal species in which LA-MRSA CC398 strains have been isolated. The first case in rabbits was reported on a commercial farm in Italy, which also involved farm workers and their families [
21]. Recently, LA-MRSA strains have been described to belong to ST2855 (CC96), ST146 (CC5), ST398 (CC398) and ST4774 (CC130) from several rabbitries of the Iberian Peninsula [
22]. These CC have been related to illness in humans [
18,
23,
24,
25,
26].
MRSA strains have been isolated in human hospitals and on animal farms where antibiotics are regularly used. These antibiotic-resistant organisms can spread to communities and the environment [
27]. Therefore, free-living animals might be colonized or infected by human and livestock sources, and can be associated with contaminated environments, even though they do not directly come into contact with antimicrobial drugs [
28].
In recent studies, a high
S. aureus carriage rate has been detected in wild animals, including European wild rabbits. The most relevant finding was that all the isolates were MRSA [
29], which suggests a wildlife MRSA reservoir. Additionally MRSA, belonging to CC130, has been detected in diseased European brown hares (
Lepus europaeus) [
30,
31].
The wild rabbit (
Oryctolagus cuniculus) is an extremely abundant endemic species of the Iberian Peninsula in some areas, where conservation of endangered predators is essential, but where major crop damage is a problem [
32]. Rabbits and wild hares are also subject to hunting and are subsequently consumed domestically, often with no adequate veterinary control or sanitary management.
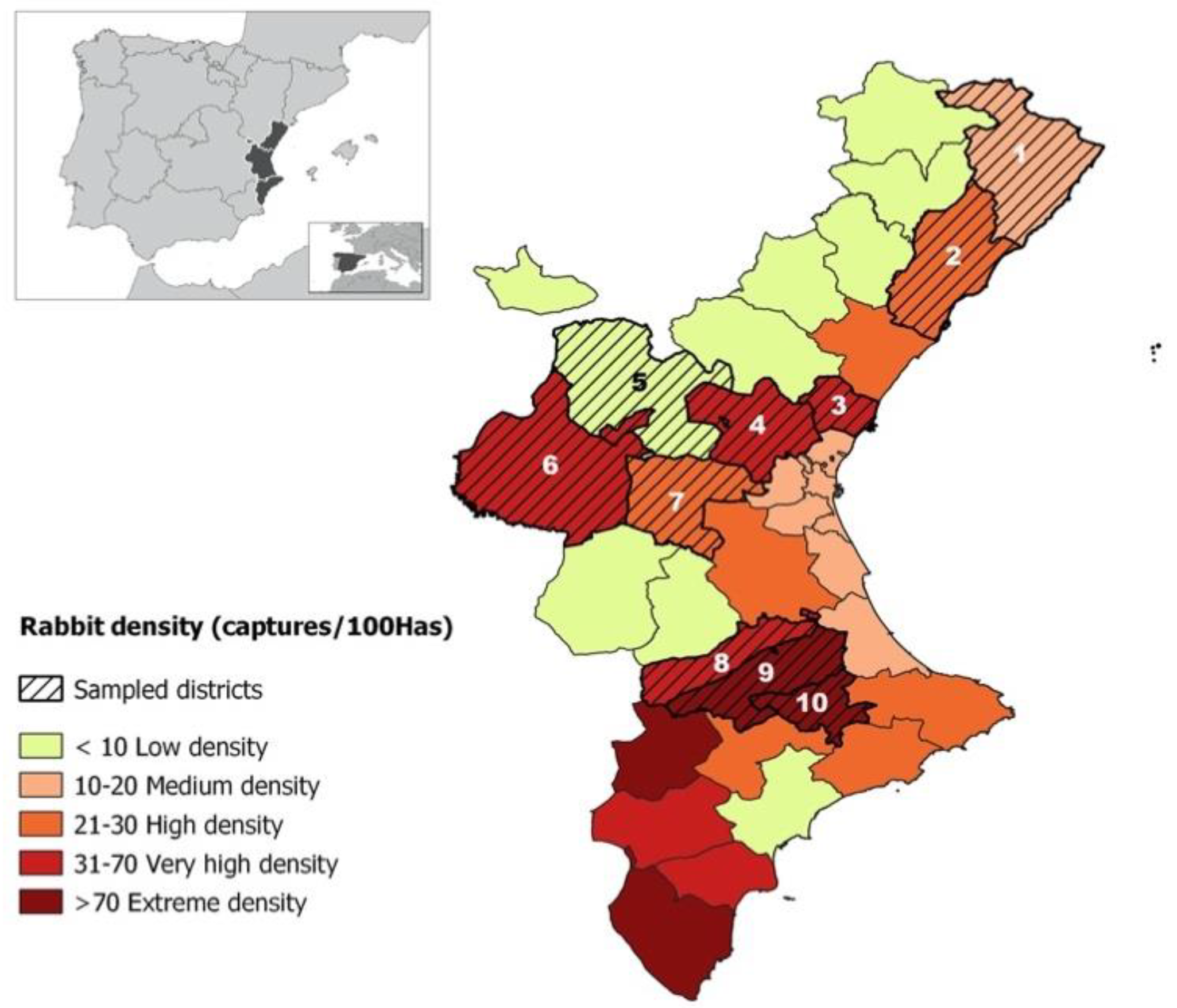
The role of S. aureus as a possible pathogen or colonizer in wild populations of lagomorphs has not yet been studied in a massive and systematic manner. In high-density areas, the interaction between rabbits and other animals is higher and, therefore, the probability of S. aureus transmission will also be higher. For these reasons, this study sets out the following objectives: (1) to know the role of wild rabbits and hares as a reservoir of S. aureus in wild lagomorph high-density areas; (2) to characterize the S. aureus strains isolated from wild rabbits and hares to compare them with those obtained previously on commercial farms; (3) to study the prevalence of MRSA strains in wild populations.
4. Discussion
In the present study, 244 S. aureus isolates obtained from 150 wild rabbits and hares, located in 10 high-density population areas (districts) of the Valencia Region in Spain, were analyzed. As far as the authors are aware, this is the first time that a study with such a large number of animals, conducted in high-density areas of animals, has been carried out. The reason why this study was performed in rabbit high-density areas was because the probability of detecting S. aureus carrier animals was believed to be higher in these areas. In total, 41.3% (150 of 363) of the animals carried S. aureus and positive animals were detected in all the studied geographical areas. Apparently, there was no clear geographical distribution of towns with more positive animals, although they were mainly located in the center and south (La Vall d’Albaida and El Comtat) of the Valencian region. Further studies in areas with lower rabbit density would be necessary to assess the relationship between S. aureus prevalence and rabbit density of animal.
Sixty-five animals carried
S. aureus in more than one location; and two different
S. aureus strains were identified in the same anatomical location in five rabbits. This agrees with previous studies which have reported how several different
S. aureus strains can simultaneously colonize individual rabbits [
42]. This is an important issue because it could affect the diagnosis of this condition if only one colony is selected during the microbiological identification of staphylococcal infections or only one anatomical site is selected for sampling
In humans, the nose is the main ecological niche where
S. aureus resides [
2]. However, in this study, the highest percentage of
S. aureus was detected in ears (49.6%). This finding has also been observed in commercial rabbitries where the presence of
S. aureus was isolated from mainly samples taken from the ears and perineum among nine different anatomical locations [
42]. The percentage of nasal carriers (28.7%) was lower than previously described in farm rabbits with staphylococcal problems (56%) [
6], but was higher than other wild mammals (22.3%), and was substantially higher than recently described for wild rabbits (8%) [
29].
In the present study, only eight animals (6 rabbits, 2 hares) showed lesions;
S. aureus was isolated from the lesions presented by the 6 rabbits. The high percentage of samples (8 of 11) obtained from the animals positive for
S. aureus (72.7%) was surprising, especially when considering the different characteristics of the observed lesions compared to commercial rabbits [
43]. Four of the six animals with positive
S. aureus lesions (66.7%) also carried this bacterium in their noses. In humans, it is reported that nasal carriers of
S. aureus are at increased risk of acquiring infection with this pathogen [
2]. This has also been found in animals, specifically rabbits, where the colonization capacity of this bacterium plays an important role in spreading the disease [
42]. There are reports that
S. aureus carriage in rabbits can be a risk for developing clinical infections [
6]. These results agree with this asseveration because the
S. aureus strains isolated from nostrils/ear/perineum and lesions were clonally related in 100% animals. Only in one case, corresponding to rabbit OC19006, was a strain different from those from rostril/perineum identified in the lesion.
In order to understand the diffusion and possible origin of
S. aureus (especially MRSA), correctly identifying the involved strain is vital. The most frequently detected CC herein was CC130, followed by CC425. The ST121 (CC121) lineage has been identified in most nasal carriers [
6] and chronic staphylococcal infections [
12,
22,
44] in commercial rabbits. Therefore, these results indicate that the
S. aureus strains which affect commercial and wild rabbits are different, which reinforces recently observed findings in wild rabbits in Aragón (a region in north Spain), where three strains were typed as t843 (ascribed to CC130). This finding has also been observed in samples from other wild species, such as reed deer (
Dama dama) and wild boar (
Sus scrofa) [
29]. Only one hare carried
S. aureus, isolated from its nostril (MSSA), and it differed (CC121) from the strains isolated in the rabbits from the same district (CC5, CC130, CC398 and CC425). However, this number was not big enough to collect data on possible species specificity.
In the present study, 138 of the 181 CC130 strains carried
mecC and the rest were methicillin-sensitive. The isolates reported to date carrying
mecC belong mainly to common lineages in cattle, namely CC130, CC1943, and CC425, which suggests a zoonotic reservoir [
19]. Besides cattle,
mecC has been found in other farm animals, with isolates ST130 in sheep [
45,
46], ST4774 (a novel tpi single locus variant of ST130) in rabbits [
22] and isolate ST425, which caused highly virulent infection, in one rabbit [
47]. ST425 is a lineage that has been found in both wild and domestic animals, and has been previously noted in wild boar from Germany [
48], red deer from Spain [
49] and also in humans. In this work, all the ST425 strains were methicillin-sensitive. The appearance of ST425-MRSA-XI, from cattle in the UK in 2011 [
50], in wildlife (fallow deer, wild boar), and in an environmental sample from Spain [
51], should put us on alert and to follow up these lineages in the future.
One interesting finding was that the only strain to carry the
mecA gene belonged to the ST398 lineage. The first LA-MRSA case reported in rabbits for meat production belonged to this lineage [
21]. Moreover, the first case of LA-MRSA in rabbits in Spain showed limited genetic diversity (ST2855, ST146, ST398, ST4774), with ST2855 being the predominant clone [
22]. It is notorious that this study found neither this lineage nor ST146 strains. This ST belongs to CC5, which has been previously described in
S. aureus isolates from rabbit carcasses, but were not resistant to methicillin [
52].
Of all the strains, 72.5% belonged to the ST1945 lineage, which is included in CC130. ST1945, a single-locus variant of ST130, has been reported as an
mecC-carrying MRSA in humans in the UK [
50], Germany [
53], France [
54] and Spain [
55]. This lineage has been described in fecal samples taken from free-ranging wild small mammals in southern Spain, specifically in two wood mice. MRSA were detected in 7% of the analyzed wood mice [
56]. The MRSA percentage herein observed was much higher, as 57.4% of
S. aureus isolates had MRSA.
Another technique used to characterize the MRSA strains included the identification of the
agr and staphylococcal cassette chromosome
mec (SCC
mec). In our study, all the isolates belonged to
agr type III, as reported in other series [
53], except for isolate ST398, which was
agrI. CC130 presented SCC
mecXI and ST398 SCC
mecV, as previously described [
50,
57,
58].
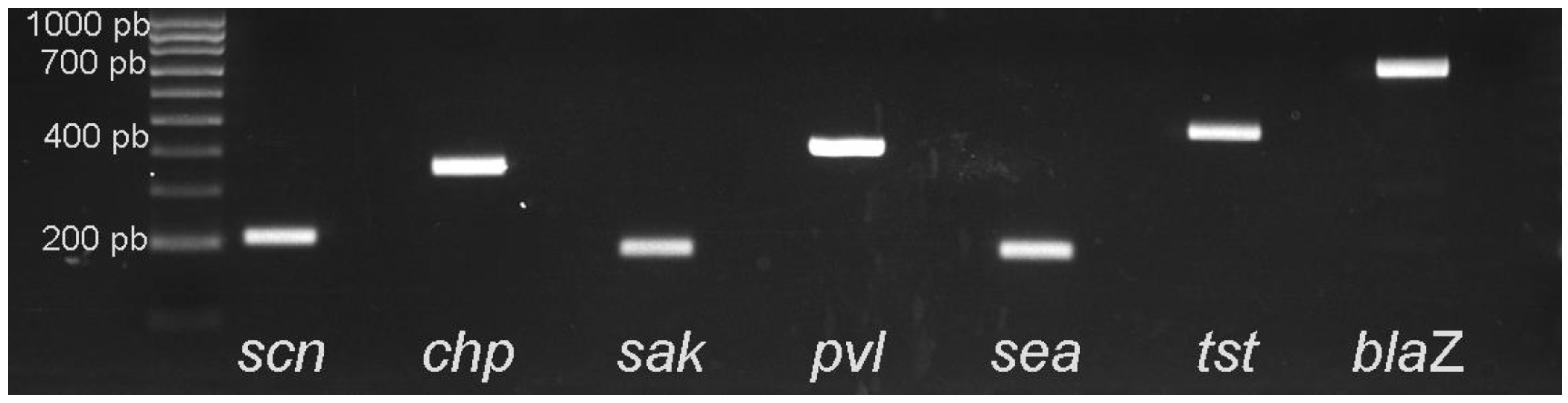
Two
mecC-positive MRSA isolates ST1945 carried genes
scn and
sak from the IEC system, and were consequently ascribed to IEC-type E. The origin of the
mecC gene is unclear, but it has been detected in staphylococci from humans and animals [
56]. Very little research has determined the presence of IEC genes in
mecC-positive isolates, and all the available research works found isolates lacking genes
sak,
chp and
scn [
31,
53,
59], which supports the hypothesis of the possible animal origin of these isolates. The detection of genes
sak and
scn in these two strains is relevant and poses questions about the potential origin of these isolates.
The possible origin of the MRSA strains remains an interesting enigma. In Spain, some wild animals (wild boars and fallow deer) located on a game estate had similar
mecC-MRSA isolates detected in river water. Therefore, it was proposed that water could be a source that disseminates this type of strain in nature [
51]. The colonization of wild animals with
S. aureus lineages from humans has been confirmed in a zoological park [
60]. The transmission of MRSA strains between animals and persons is relatively easy, because it has been observed how 22% of students who visited pork farms (30% prevalence of MRSA) became MRSA nasal carriers [
18]. It has also demonstrated that the most prevalent
S. aureus strain in commercial rabbits in Spain (ST121) has a human origin. This host adaptation took place 45 years ago when a single nucleotide mutation was sufficient to convert a human-specific
S. aureus strain into one that could infect rabbits [
61]. However, close contact between humans and animals has been assumed in all these cases, and the animals on commercial farms lived in high-density communities with interactions between animals. These conditions are not, a priori, those that take place in wild lagomorphs. However, it is worth considering that the present study entails the peculiarity of being conducted in rabbit high-density areas that could increase the probability of contact between animals and favor
S. aureus spreading through rabbit populations. Additionally, CC130 (the most isolated CC in this study) is a lineage associated mainly with cattle, with transmission patterns that might be assumed in which wild rabbits could be infected by ingesting contaminated feces from wild ruminants in the same area. Our study did not analyze water or other animals that could interact with wild rabbits (predators, ruminants, etc.). Therefore, it would be interesting to extend this study along this line in the future to know the epidemiological origin of these bacteria.
Most MRSA isolates show resistance to cefoxitin. As previously reported for
mecC-carrying MRSA, most isolates were susceptible to all the tested antibiotics, except for β-lactams [
55], and 10 strains were Bacitracin-resistant. However, the ST398-MRSA-
mecA strain also shows resistance to enrofloxacin and tetracycline, and carried the
blaZ gene. Resistance to tetracycline has also been associated with livestock-related ST398 [
62].
Food-producing animals, both livestock and wildlife, and derived products are considered potential sources of MRSA in humans [
63]. However in that study, which included 1365 wild animals,
S. aureus was detected in only 2.0% of wild animal carcasses and in 3.2% of wild boar lymph nodes, and none had MRSA. Therefore, the authors concluded that the risk of transmission to humans was limited. Conversely, in our study, the high presence of
S. aureus (41.3% of animals), of which 63.3% had MRSA, indicates an alarming situation for wild rabbits in our study area, because these animals can be consumed directly by hunters, or even be donated to charities, without undergoing adequate veterinary inspections.

 ,
, 



{kind=link}
{kind=link}