Sheep and Goats Respond Differently to Feeding Strategies Directed to Improve the Fatty Acid Profile of Milk Fat


 , , , ,
, , , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Feeding Strategy to Improve the Fatty Acid Content in Sheep and Goat Milk
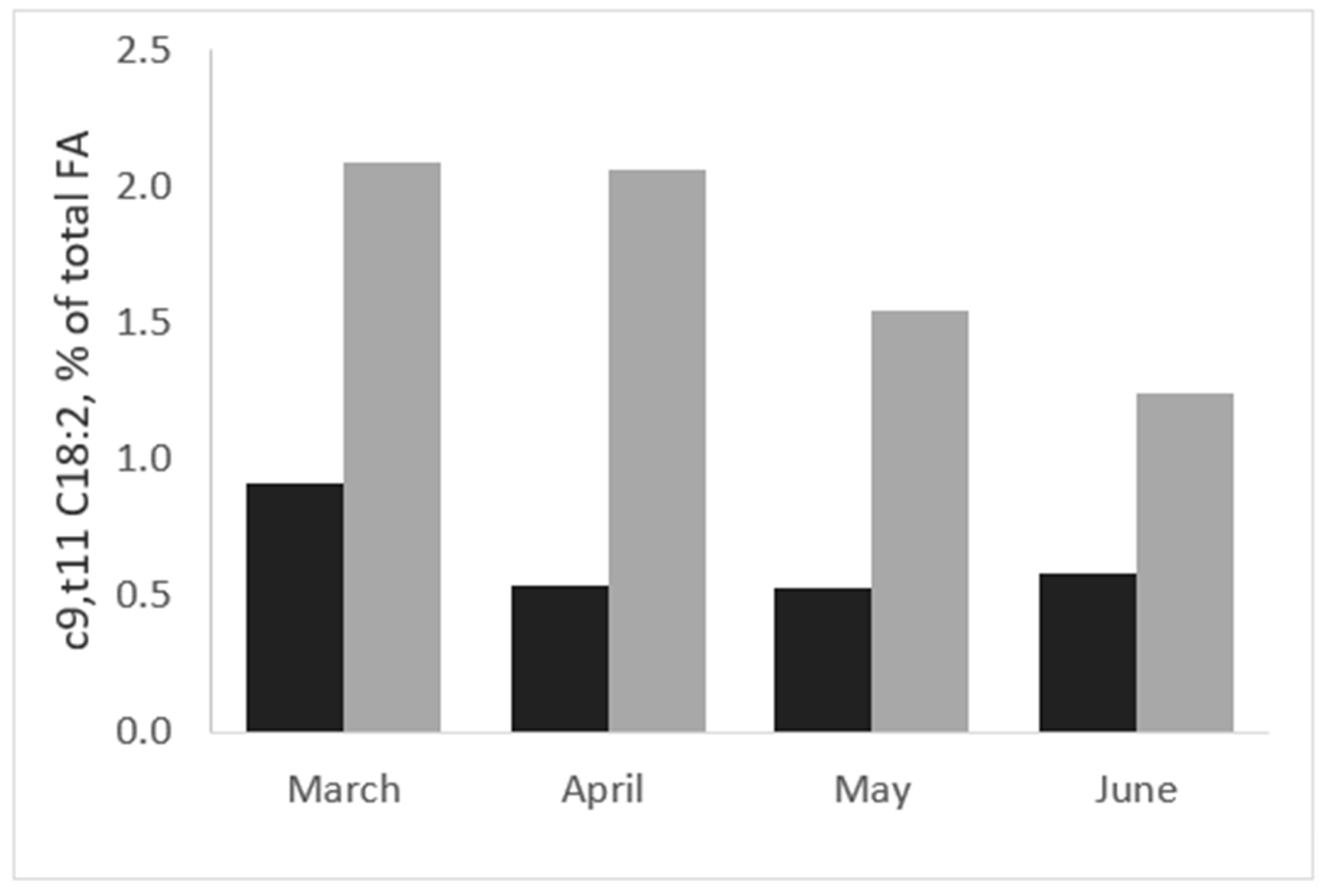
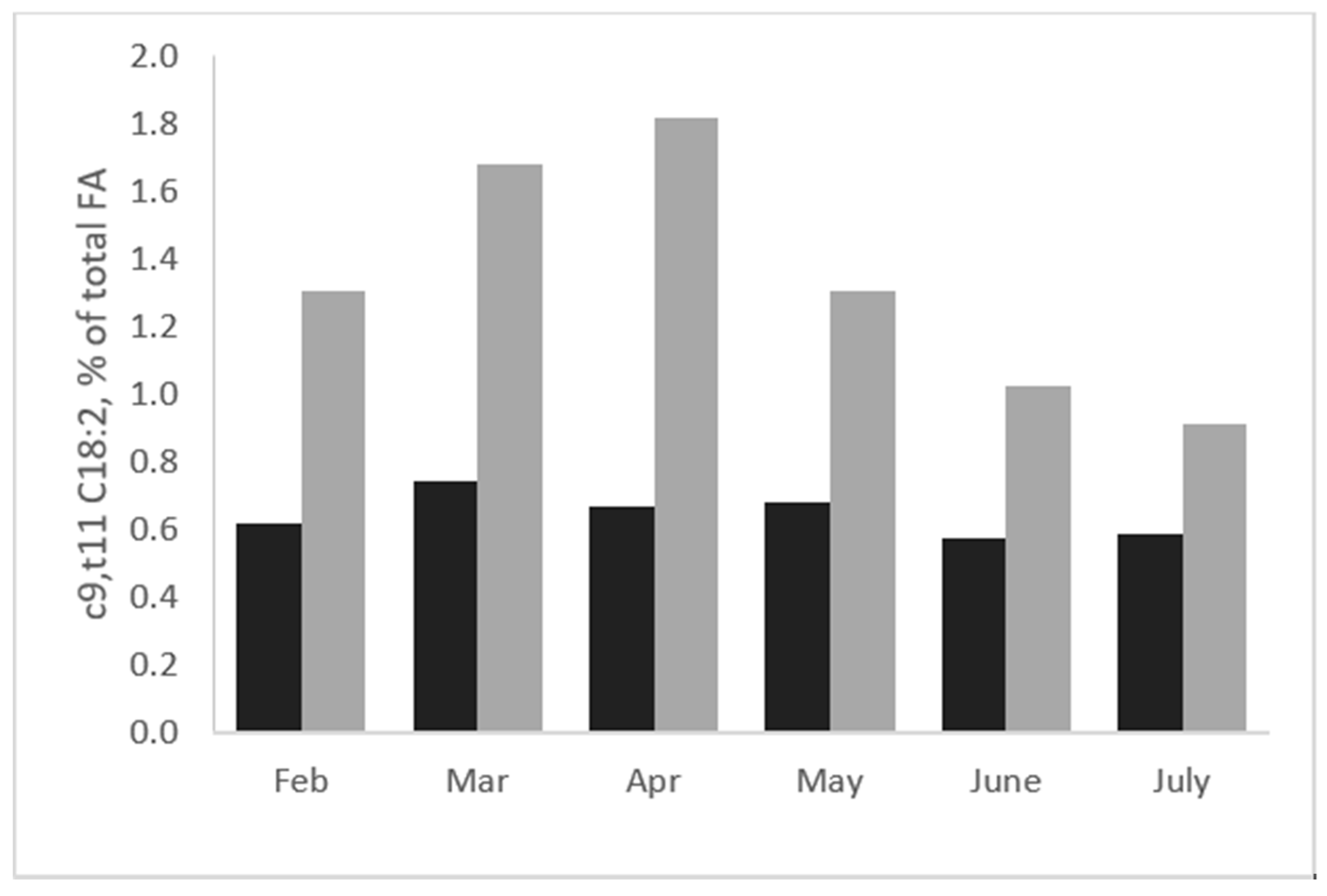
2.1. Effects of Grass and Forage on Milk Fat Quality
2.2. Effects of Vegetable Oil on Milk Fat Quality
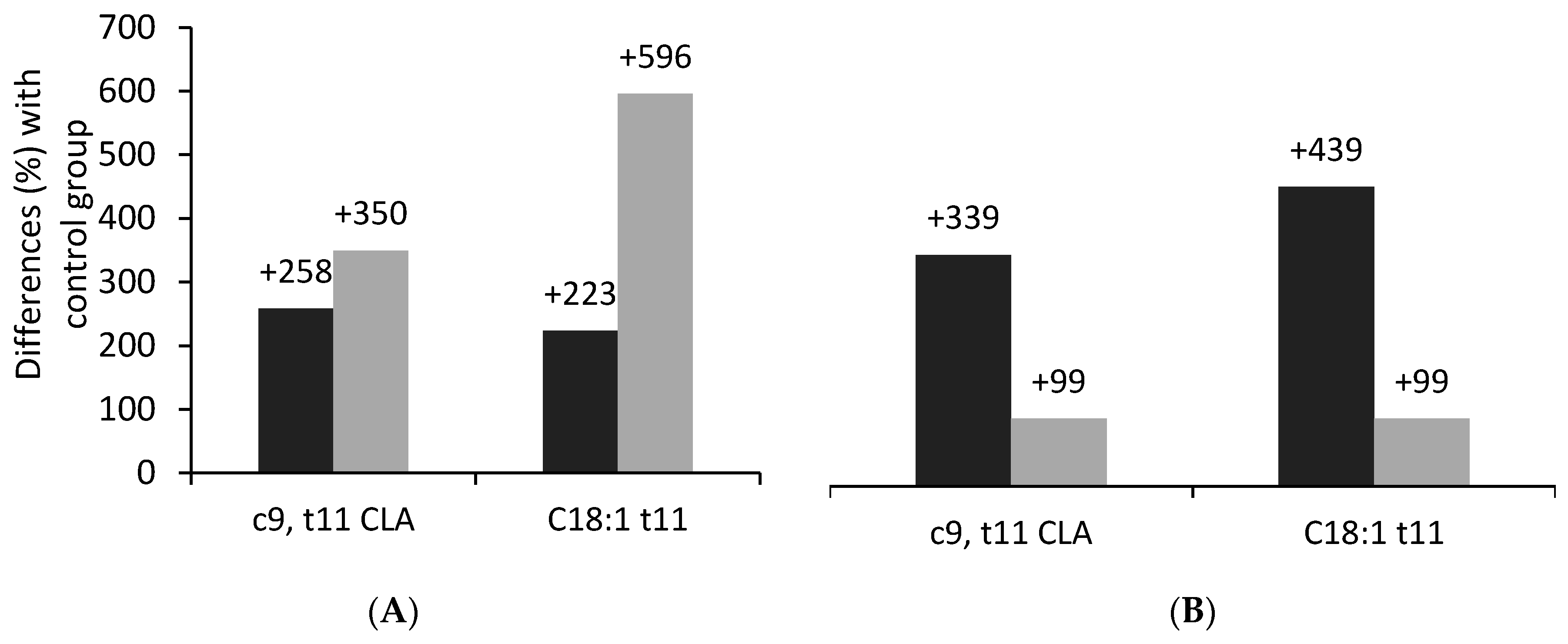
2.3. Effects of Fish Oil and Algae Supplementation on the Fatty Acid Profile of Sheep and Goat Milk

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Diet 2 | Milk Fatty Acids (%) 1 | References | |||
|---|---|---|---|---|---|---|
| RA | ALA | EPA + DHA | n6:n3 | |||
| Goats | 55.5 g/day SO + 11.1 g/day FO | 609 | 13 | 260 | 61 | [73] |
| 27.0 g/day Chlorella kessleri microalga | 28 | 33 | 120 | −18 | [90] | |
| 10.0 g/day Chlorella vulgaris microalgae | −11 | −6 | 7 | 18 | [94] | |
| 10.0 g/day Japonochytrium sp. microalgae | −1 | −12 | 21 | −11 | [94] | |
| 50.0 g/day FO | −9 | −71 | [93] | |||
| 20.0 g/day FO | 1377 | −74 | [86] | |||
| 40.0 g/day MA | 18 | −71 | [85] | |||
| Sheep | 30.0 g/day FO | 146 | 18 | 500 | −21 | [95] |
| 45.0 g/day FO | 322 | 8 | 960 | −15 | [95] | |
| 6.0 g/day DHA | 2863 | −60 | [88] | |||
| 25.0 g SUN + 8.0 g MA | 126 | −10 | 200 | 9 | [91] | |
| 25.0 g SUN + 16.0 g MA | 110 | −12 | 556 | −30 | [91] | |
| 25.0 g SUN + 24.0 g MA | 162 | −17 | 700 | −31 | [91] | |
| 0.25% FO + 3.75% SO | 10 | 0 | 400 | [96] | ||
| 0.50% FO + 3.50% SO | 22 | 0 | 500 | [96] | ||
| 0.75% FO + 3.25% SO | 7 | −8 | 600 | [96] | ||
| 25.0 g SO + 8.0 g MA/kg DM | 65 | −2 | 389 | −23 | [97] | |
2.4. Effects of Essential Oils on the Fatty Acid Profile of Sheep and Goat Milk
| Species | Source | Dietary EO 2 | Milk Fatty Acids (%) 1 | References | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SFA | MUFA | PUFA | SA | VA | RA | LA | ALA | ||||
| Goats | Rosmarinus officinalis spp. | 10% of control diet | −2 | 1 | 38 | −4 | 36 | 0 | [103] | ||
| Rosmarinus officinalis spp. | 20% of control diet | −2 | −1 | 54 | 4 | 55 | 23 | [103] | |||
| Cuminum cyminum | 1.27 g/kg DMI | −2 | 2 | 22 | −4 | 7 | 25 | 23 | 23 | [104] | |
| Cuminum cyminum | 2.53 g/kg DMI | −3 | 1 | 25 | −7 | 7 | 24 | 25 | 29 | [104] | |
| Garlic oil | 0.57 g/kg DM | 0 | 1 | 4 | 0 | 1 | 7 | −1 | 10 | [105] | |
| Garlic oil | 1.14 g/kg DM | 0 | 0 | 3 | 1 | 3 | 12 | −1 | 5 | [105] | |
| Garlic oil | 1.71 g/kg DM | −1 | 1 | 6 | 2 | 4 | 12 | −1 | 10 | [105] | |
| Citral oil | 0.08 mL of EO/kg BW | 3 | −16 | −15 | −3 | [106] | |||||
| Citral oil | 0.16 mL of EO/kg BW | 2 | −3 | −4 | −3 | [106] | |||||
| Sheep | Citral oil | 0.24 mL of EO/kg BW | 3 | −9 | −4 | −5 | [106] | ||||
| α-pinene, limonene, β-caryophyllene | 1 mL of each in a 10-mL soya bean oil mixture | 0 | 6 | 5 | −3 | −8 | 37 | [107] | |||
| Carum seeds | High dose | −7 | 38 | 5 | 71 | 86 | 10 | 6 | −12 | [100] | |
| Coriandrum seeds | High dose | −7 | 45 | −15 | 97 | 152 | −17 | −16 | −21 | [100] | |
| Satureja leaves | High dose | 0 | 0 | −5 | 4 | −9 | −15 | −13 | 0 | [100] | |
| Orange peel | 150 mg/kg | 5 | −11 | −10 | −13 | 3 | 1 | −12 | −17 | [102] | |
| Orange peel | 300 mg/kg | 5 | −12 | −15 | −12 | −10 | −10 | −15 | −17 | [102] | |
| Orange peel | 450 mg/kg | 1 | −3 | −7 | −1 | −8 | −3 | −8 | −9 | [102] | |
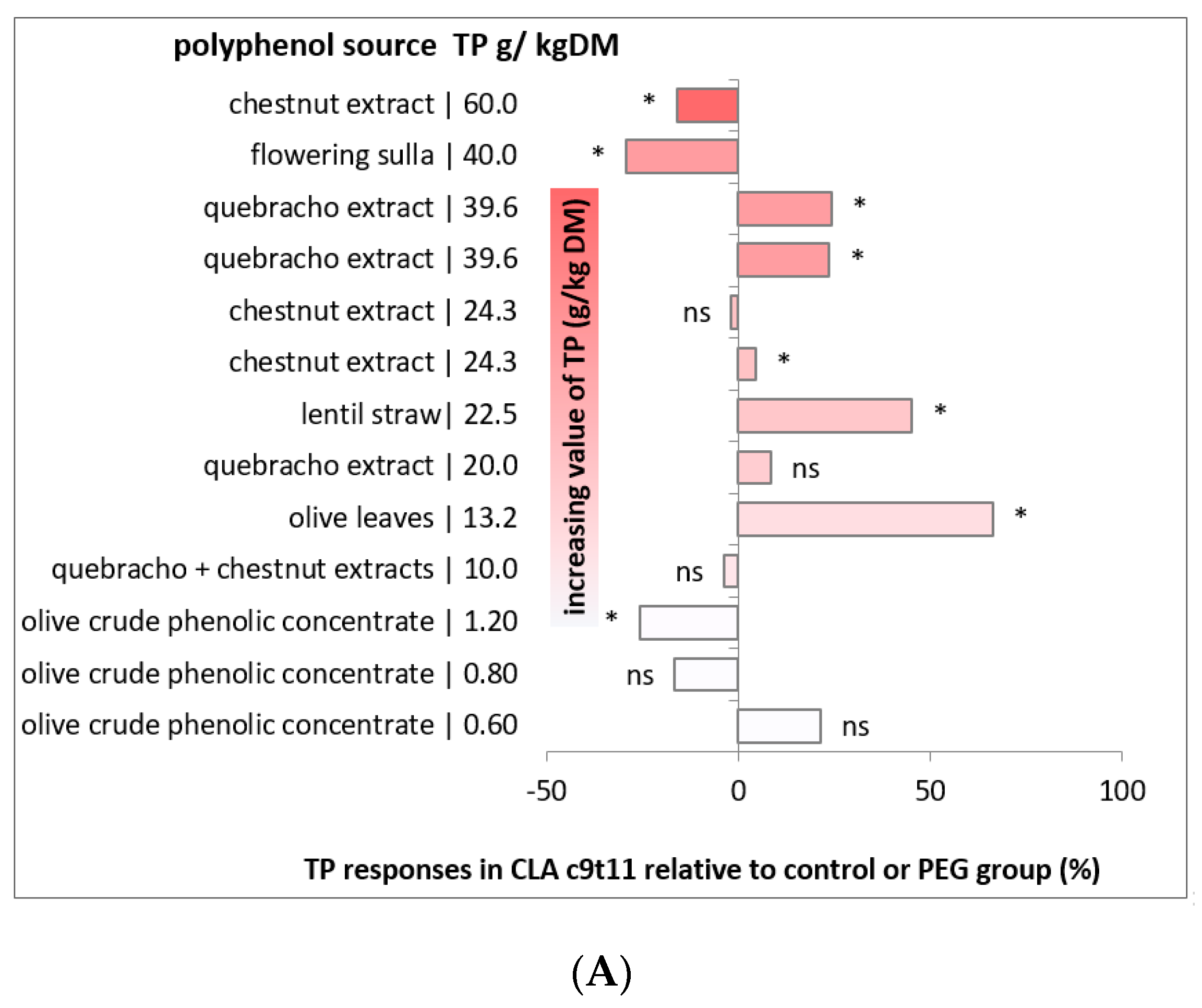
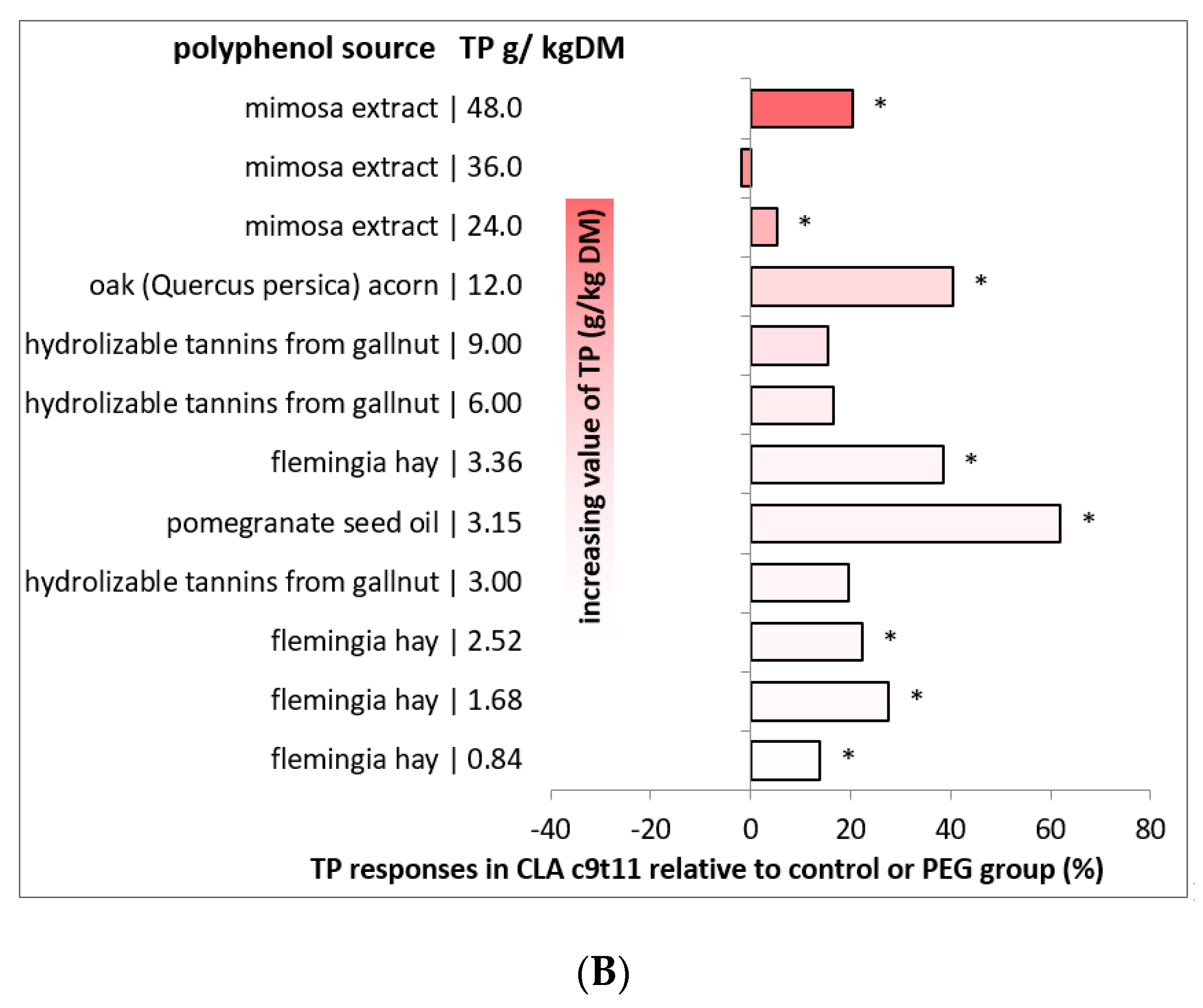
2.5. Effects of Dietary Polyphenols on the Fatty Acid Profile of Sheep and Goat Milk
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pulina, G.; Milán, M.J.; Lavín, M.P.; Theodoridis, A.; Morin, E.; Capote, J.; Thomas, D.L.; Francesconi, A.H.D.; Caja, G. Invited review: Current production trends, farm structures, and economics of the dairy sheep and goat sectors. J. Dairy Sci. 2018, 101, 6715–6729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAOSTAT (Food and Agriculture Organization of the United Nations). 2018 Statistics Database. Available online: http://www.fao.org/faostat/en/#data (accessed on 9 July 2020).
- Sepe, L.; Argüello, A. Recent advances in dairy goat products. Asian-Australas. J. Anim. Sci. 2019, 32, 1306. [Google Scholar] [CrossRef] [Green Version]
- Pfeuffer, M.; Jaudszus, A. Pentadecanoic and Heptadecanoicheptadecanoic acids: Multifaceted odd–chain fatty acids. Adv. Nutr. 2016, 7, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, F.; Fretts, A.; Marklund, M.; Ardisson Korat, A.V.A.; Yang, W.S.; Lankinen, M.; Qureshi, W.; Helmer, C.; Chen, T.A.; Wong, K.; et al. Fatty acids and outcomes research consortium (FORCE). Fatty acid biomarkers of dairy fat consumption and incidence of type 2 diabetes: A pooled analysis of prospective cohort studies. PLoS Med. 2018, 15, e1002670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlaeminck, B.; Fie vez, V.; Tamminga, S.; Dewhurst, R.J.; van Vuuren, A.; De Brabander, D.; Demeyer, D. Milk odd—And branched—Chain fatty acids in relation to the rumen fermentation pattern. J. Dairy Sci. 2006, 89, 3954–3964. [Google Scholar] [CrossRef] [Green Version]
- Prandini, A.; Sigolo, S.; Piva, G. A comparative study of fatty acid composition and CLA concentration in commercial cheeses. J. Food Comp. Anal. 2011, 24, 55–61. [Google Scholar] [CrossRef]
- Lobos-Ortega, I.; Revilla, I.; González-Martín, M.I.; Hernández-Hierro, J.M.; Vivar-Quintana, A.; González-Pérez, C. Conjugated linoleic acid contents in cheeses of different compositions during six months of ripening. Czech J. Food Sci. 2012, 30, 220–226. [Google Scholar] [CrossRef] [Green Version]
- Kepler, C.R.; Hirons, K.P.; McNeill, J.J.; Tove, S.B. Intermediates and products of the biohydrogenation of linoleic acid by Butyrivibrio fibrisolvens. J. Biol. Chem. 1966, 241, 1350–1354. [Google Scholar]
- Szumacher-Strabel, M.; Cieslak, A.; Nowakowska, A. Effect of oils rich in linoleic acid on in vitro rumen fermentation parameters of sheep, goats and dairy cows. J. Anim. Feed Sci. 2009, 18, 440–452. [Google Scholar] [CrossRef]
- Yang, B.; Chen, H.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, Y.Q.; Chen, W. Review of the roles of conjugated linoleic acid in health and disease. J. Funct. Foods 2015, 15, 314–325. [Google Scholar] [CrossRef]
- Ferlay, A.; Bernard, L.; Meynadier, A.; Malpuech-Brugère, C. Production of trans and conjugated fatty acids in dairy ruminants and their putative effects on human health: A review. Biochimie 2017, 141, 107–120. [Google Scholar] [CrossRef]
- Bergamo, P.; Palmieri, G.; Cocca, E.; Ferrandino, I.; Gogliettino, M.; Monaco, A.; Maurano, F.; Rossi, M. Adaptive response activated by dietary cis9, trans11 conjugated linoleic acid prevents distinct signs of gliadin-induced enteropathy in mice. Eur. J. Nutr. 2016, 55, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Bachmair, E.M.; Bots, M.L.; Mennen, L.I.; Kelder, T.; Evelo, C.T.; Horgan, G.W.; Ford, I.; de Roos, B. Effect of supplementation with an 80:20 cis9, trans11 conjugated linoleic acid blend on the human platelet proteome. Mol. Nutr. Food Res. 2012, 56, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, X.; Zhang, X.; Liu, J.; Ye, S.; Xiao, S.; Chen, H.; Wang, H. Induction of apoptosis by c9, t11–CLA in human endometrial cancer RL 95–2 cells via ERα-mediated pathway. Chem. Phys. Lipids 2013, 175, 27–32. [Google Scholar] [CrossRef]
- Lu, G.; Zhang, G.; Zheng, X.; Zeng, Y.; Xu, Z.; Zeng, W.; Wang, K. c9, t11-conjugated linoleic acid induces HCC cell apoptosis and correlation with PPAR-γ signaling pathway. Am. J. Transl. Res. 2015, 7, 2752–2763. [Google Scholar]
- Penedo, L.A.; Nunes, J.C.; Gama, M.A.S.; Leite, P.E.C.; Quirico-Santos, T.F.; Torres, A.G. Intake of butter naturally enriched with cis9, trans11 conjugated linoleic acid reduces systemic inflammatory mediators in healthy young adults. J. Nutr. Biochem. 2013, 24, 2144–2151. [Google Scholar] [CrossRef] [PubMed]
- Mollica, M.P.; Trinchese, G.; Cavaliere, G.; De Filippo, C.; Cocca, E.; Gaita, M.; Della-Gatta, A.; Marano, A.; Mazzarella, G.; Bergamo, P. c9, t11-Conjugated linoleic acid ameliorates steatosis by modulating mitochondrial uncoupling and Nrf2 pathway. J. Lipid Res. 2014, 55, 837–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pintus, S.; Murru, E.; Carta, G.; Cordeddu, L.; Batetta, B.; Accossu, S.; Pistis, D.; Uda, S.; Ghiani, M.E.; Mele, M.; et al. Sheep cheese naturally enriched in α-linolenic, conjugated linoleic and vaccenic acids improves the lipid profile and reduces anandamide in the plasma of hypercholesterolaemic subjects. Brit. J. Nutr. 2013, 109, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Derakhshande-Rishehri, S.M.; Mansourian, M.; Kelishadi, R.; Heidari-Beni, M. Association of foods enriched in conjugated linoleic acid (CLA) and CLA supplements with lipid profile in human studies: A systematic review and meta-analysis. Public Health Nutr. 2015, 18, 2041–2054. [Google Scholar] [CrossRef] [Green Version]
- Sofi, F.; Buccioni, A.; Cesari, F.; Gori, A.M.; Minieri, S.; Mannini, L.; Casini, A.; Gensini, G.F.; Abbate, R.; Antongiovanni, M. Effects of a dairy product (pecorino cheese) naturally rich in cis9, trans-11 conjugated linoleic acid on lipid, inflammatory and haemorheological variables: A dietary intervention study. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Krogager, T.P.; Nielsen, L.V.; Kahveci, D.; Dyrlund, T.F.; Scavenius, C.; Sanggaard, K.W.; Enghild, J.J. Hepatocytes respond differently to major dietary trans fatty acid isomers, elaidic acid and trans–vaccenic acid. Proteome Sci. 2015, 13, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacome-Sosa, M.; Vacca, C.; Mangat, R.; Diane, A.; Nelson, R.C.; Reaney, M.J.; Shen, J.; Curtis, J.M.; Vine, D.F.; Field, C.J.; et al. Vaccenic acid suppresses intestinal inflammation by increasing anandamide and related N–acylethanolamines in the JCR:LA-cp rat. J. Lipid Res. 2016, 57, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, M.S.; Bilodeau, J.F.; Larose, J.; Greffard, K.; Julien, P.; Barbier, O.; Rudkowska, I. Modulation of the biomarkers of inflammation and oxidative stress by ruminant trans fatty acids and dairy proteins in vascular endothelial cells (HUVEC). Prostaglandins Leukot. Essent. Fatty Acids 2017, 126, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Lin, L.; Shi, B.; Jing, J.; Cai, L. The effects of early life polyunsaturated fatty acids and ruminant trans fatty acids on allergic diseases: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2018, 59, 18021–181514. [Google Scholar] [CrossRef] [PubMed]
- Del Gobbo, L.C.; Imamura, F.; Aslibekyan, S.; Marklund, M.; Virtanen, J.K.; Wennberg, M.; Yakoob, M.Y.; Chiuve, S.E.; dela Cruz, L.; Frazier-Wood, A.C.; et al. ω-3 polyunsaturated fatty acid biomarkers and coronary heart disease: Pooling project of 19 cohort studies. JAMA Intern. Med. 2016, 176, 1155–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbeau, P.A.; Holloway, T.M.; Whitfield, J.; Baechler, B.L.; Quadrilatero, J.; van Loon, L.J.C.; Chabowski, A.; Holloway, G.P. α-Linolenic acid and exercise training independently, and additively, decrease blood pressure and prevent diastolic dysfunction in obese Zucker rats. J. Physiol. 2017, 595, 4351–4364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, R.; Hasanally, D.; Stamenkovic, A.; Maddaford, T.G.; Chaudhary, R.; Pierce, G.N.; Ravandi, A. Alpha linolenic acid decreases apoptosis and oxidized phospholipids in cardiomyocytes during ischemia/reperfusion. Mol. Cell. Biochem. 2018, 437, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, K.; Ikeda, A.; Chei, C.L.; Noda, H.; Umesawa, M.; Cui, R.; Muraki, I.; Ohira, T.; Imano, H.; Sankai, T.; et al. Serum α-linolenic and other ω-3 fatty acids, and risk of disabling dementia: Community-based nested case-control study. Clin. Nutr. 2017, 36, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Chilliard, Y.; Ferlay, A.; Rouel, J.; Lamberet, G. A review of nutritional and physiological factors affecting goat milk lipid synthesis and lipolysis. J. Dairy Sci. 2003, 86, 1751–1770. [Google Scholar] [CrossRef] [Green Version]
- Nudda, A.; Battacone, G.; Boaventura Neto, O.; Cannas, A.; Francesconi, A.H.D.; Atzori, A.S.; Pulina, G. Feeding strategies to design the fatty acid profile of sheep milk and cheese. Rev. Bras. Zootec. 2014, 43, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Tsiplakou, E.; Mountzouris, K.C.; Zervas, G. Concentration of conjugated linoleic acid in grazing sheep and goat milk fat. Livest. Sci. 2006, 103, 74–84. [Google Scholar] [CrossRef]
- Nudda, A.; Manca, M.G.; Rubattu, R.; Boe, R.; Battacone, G. Fatty acids content with nutraceutical properties in Sardinian ovine and caprine cheese: A survey. In Proceedings of the 43th International Symposium SIPZOO on Nutripharm and Biosecurity, Alghero, Italy, 30 May 2008. [Google Scholar]
- Tudisco, R.; Cutrignelli, M.I.; Calabrò, S.; Piccolo, G.; Bovera, F.; Guglielmelli, A.; Moniello, G.; Infascelli, F. Influence of organic systems on milk fatty acid profile and CLA in goats. Small Rumin. Res. 2010, 88, 151–155. [Google Scholar] [CrossRef]
- Nudda, A.; Battacone, G.; Usai, M.G.; Fancellu, S.; Pulina, G. Supplementation with extruded linseed cake affects concentrations of conjugated linolenic acid and vaccenic acid in goat milk. J. Dairy Sci. 2006, 89, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Nudda, A.; Battacone, G.; Atzori, A.S.; Dimauro, C.; Rassu, S.P.G.; Nicolussi, P.; Bonelli, P.; Pulina, G. Effect of extruded linseed supplementation on blood metabolic profile and milk performance of Saanen goats. Animal 2013, 7, 1464–1471. [Google Scholar] [CrossRef]
- Valenti, B.; Pagano, R.I.; Pennisi, P.; Lanza, M.; Avondo, M. Polymorphism at αs1-casein locus. Effect of genotype × diet interaction on milk fatty acid composition in Girgentana goat. Small Rumin. Res. 2010, 94, 210–213. [Google Scholar] [CrossRef]
- Chilliard, Y.; Toral, P.G.; Shingfield, K.J.; Rouel, J.; Leroux, C.; Bernard, L. Effects of diet and physiological factors on milk fat synthesis, milk fat composition and lipolysis in the goat: A short review. Small Rumin. Res. 2014, 122, 31–37. [Google Scholar] [CrossRef]
- Jahreis, G.; Fritsche, J.; Kraft, J. Species-dependent, seasonal, and dietary variation of conjugated linoleic acid in milk. In Advances in Conjugated Linoleic Acid Research; Yurawecz, M.P., Mossoba, M.M., Kramer, J.K.G., Pariza, M.W., Nelson, G.J., Eds.; AOCS Press: Champaign, IL, USA, 1999; Volume 1, pp. 215–225. [Google Scholar]
- Nudda, A.; Mele, M.; Battacone, G.; Usai, M.G.; Macciotta, N.P.P. Comparison of conjugated linoleic acid (CLA) content in milk of ewes and goats with the same dietary regimen. Ital. J. Anim. Sci. 2003, 2, 515–517. [Google Scholar] [CrossRef]
- Nudda, A.; McGuire, M.A.; Battacone, G.; Pulina, G. Seasonal variation in conjugated linoleic acid and vaccenic acid in milk fat of sheep and its transfer to cheese and ricotta. J. Dairy Sci. 2005, 88, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Nudda, A.; Battacone, G.; Testone, S.; Pulina, G. Seasonal variation of conjugated linoleic acid (CLA) and n-3 fatty acids of goat milk fat and its transfer into cheese. J. Dairy Sci. 2007, 90, 483. [Google Scholar]
- deRenobales, M.; Amores, G.; Arranz, J.; Virto, M.; Barrón, L.J.R.; Bustamante, M.A.; Ruiz De Gordoa, J.C.; Nájera, A.I.; Valdivielso, I.; Abilleira, E.; et al. Part-time grazing improves sheep milk production and its nutritional characteristics. Food Chem. 2012, 130, 90–96. [Google Scholar] [CrossRef]
- Renna, M.; Lussiana, C.; Cornale, P.; Fortina, R.; Mimosi, A. Changes in goat milk fatty acids during abrupt transition from indoor to pasture diet. Small Rumin. Res. 2012, 108, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Addis, M.; Cabiddu, A.; Pinna, G.; Decandia, M.; Piredda, G.; Pirisi, A.; Molle, G. Milk and cheese fatty acid composition in sheep fed Mediterranean forages with reference to conjugated linoleic acid cis-9, trans-11. J. Dairy Sci. 2005, 88, 3443–3454. [Google Scholar] [CrossRef] [Green Version]
- Cabiddu, A.; Decandia, M.; Addis, M.; Piredda, G.; Pirisi, A.; Molle, G. Managing Mediterranean pastures in order to enhance the level of beneficial fatty acids in sheep milk. Small Rumin. Res. 2005, 59, 169–180. [Google Scholar] [CrossRef]
- Fedele, V.; Pizzillo, M.; Claps, S.; Morand-Fehr, P.; Rubino, R. Grazing behavior and diet selection of goats on native pasture in Southern Italy. Small Rumin. Res. 1993, 11, 305–322. [Google Scholar] [CrossRef]
- De Rosa, G.; Moio, L.; Napolitano, F.; Grasso, F.; Gubitosi, L.; Bordi, A. Influence of flavor on goat feeding preferences. J. Chem. Ecol. 2002, 28, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.M.M.; Celaya, R.; Benavides, R.; Jáuregui, B.M.; García, U.; Santos, A.S.; García, R.R.; Rodrigues, M.A.M.; Osoro, K. Foraging behaviour of domestic herbivore species grazing on heathlands associated with improved pasture areas. Livest. Sci. 2013, 155, 373–383. [Google Scholar] [CrossRef]
- Abijaoudé, J.A.; Morand-Fehr, P.; Tessier, J.; Schmidely, P.; Sauvant, D. Contribution of main and secondary meals to the daily intake of stall-housed dairy goats in mid lactation. Sheep Goats Nutr. Intake Dig. Qual. Prod. Rangel. Cah. Options Méditerranéennes 2000, 52, 33–37. [Google Scholar]
- Gómez-Cortés, P.; Toral, P.G.; Frutos, P.; Juárez, M.; De La Fuente, M.A.; Hervás, G. Effect of the supplementation of dairy sheep diet with incremental amounts of sunflower oil on animal performance and milk fatty acid profile. Food Chem. 2011, 125, 644–651. [Google Scholar] [CrossRef]
- Albenzio, M.; Santillo, A.; Avondo, M.; Nudda, A.; Chessa, S.; Pirisi, A.; Banni, S. Nutritional properties of small ruminant food products and their role on human health. Small Rumin. Res. 2016, 135, 3–12. [Google Scholar] [CrossRef]
- Cieslak, A.; Kowalczyk, J.; Czauderna, M.; Potkanski, A.; Szumacher-Strabel, M. Enhancing unsaturated fatty acids in ewe’s milk by feeding rapeseed or linseed oil. Czech J. Anim. Sci. 2010, 55, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Sanz Sampelayo, M.R.; Chilliard, Y.; Schmidely, P.; Boza, J. Influence of type of diet on the fat constituents of goat and sheep milk. Small Rumin. Res. 2007, 68, 42–63. [Google Scholar] [CrossRef]
- Nguyen, Q.V.; Malau-Aduli, B.S.; Cavalieri, J.; Malau-Aduli, A.E.; Nichols, P.D. Enhancing Omega-3 long-chain polyunsaturated fatty acid content of dairy- derived foods for human consumption. Nutrients 2019, 11, 743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nudda, A.; Palmquist, D.L.; Battacone, G.; Fancellu, S.; Rassu, S.P.G.; Pulina, G. Relationships between the contents of vaccenic acid, CLA and n− 3 fatty acids of goat milk and the muscle of their suckling kids. Livest. Sci. 2008, 118, 195–203. [Google Scholar] [CrossRef]
- Emami, A.; Fathi Nasri, M.H.; Ganjkhanlou, M.; Rashidi, L.; Zali, A. Effect of pomegranate seed oil as a source of conjugated linolenic acid on performance and milk fatty acid profile of dairy goats. Livest. Sci. 2016, 193, 1–7. [Google Scholar] [CrossRef]
- Bernard, L.; Toral, P.; Rouel, J.; Chilliard, Y. Effects of extruded linseed and level and type of starchy concentrate in a diet containing fish oil on dairy goat performance and milk fatty acid composition. Anim. Feed Sci. Technol. 2016, 222, 31–42. [Google Scholar] [CrossRef]
- Bernard, L.; Shingfield, K.J.; Rouel, J.; Ferlay, A.; Chilliard, Y. Effect of plant oils in the diet on performance and milk fatty acid composition in goats fed diets based on grass hay or maize silage. Br. J. Nutr. 2009, 101, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Chilliard, Y.; Rouel, J.; Guillouet, P. Goat alpha-s1 casein genotype interacts with the effect of extruded linseed feeding on milk fat yield, fatty acid composition and post-milking lipolysis. Anim. Feed Sci. Technol. 2013, 185, 140–149. [Google Scholar] [CrossRef]
- Gómez-Cortés, P.; Cívico, A.; de la Fuente, M.A.; Sánchez, N.N.; Blanco, F.P.; Marín, A.M. Effects of dietary concentrate composition and linseed oil supplementation on the milk fatty acid profile of goats. Animal 2018, 12, 2310–2317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.H.; Mustafa, A.F.; Zhao, X. Effects of feeding oil seeds rich in linoleic and linolenic fatty acids to lactating ewes on cheese yield and on fatty acid composition of milk and cheese. Anim. Feed Sci. Technol. 2006, 127, 220–233. [Google Scholar] [CrossRef]
- Correddu, F.; Gaspa, G.; Pulina, G.; Nudda, A. Grapeseed and linseed, alone and in combination, enhance unsaturated fatty acids in the milk of Sarda dairy sheep. J. Dairy Sci. 2016, 99, 1725–1735. [Google Scholar] [CrossRef]
- Cabiddu, A.; Addis, M.; Fiori, M.; Spada, S.; Decandia, M.; Molle, G. Pros and cons of the supplementation with oil seed enriched concentrates on milk fatty acid profile of dairy sheep grazing Mediterranean pastures. Small Rumin. Res. 2017, 147, 63–72. [Google Scholar] [CrossRef]
- Bodas, R.; Manso, T.; Mantecón, A.R.; Juárez, M.; De La Fuente, M.A.; Gómez-Cortés, P. Comparison of the fatty acid profiles in cheeses from ewes fed diets supplemented with different plant oils. J. Agric. Food Chem. 2010, 58, 10493–10502. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Mustafa, A.F.; Zhao, X. Effects of flaxseed supplementation to lactating ewes on milk composition, cheese yield, and fatty acid composition of milk and cheese. Small Rumin. Res. 2006, 63, 233–241. [Google Scholar] [CrossRef]
- Mele, M.; Contarini, G.; Cercaci, L.; Serra, A.; Buccioni, A.; Povolo, M.; Conte, G.; Funaro, A.; Banni, S.; Lercker, G.; et al. Enrichment of Pecorino cheese with conjugated linoleic acid by feeding dairy ewes with extruded linseed: Effect on fatty acid and triglycerides composition and on oxidative stability. Int. Dairy J. 2011, 21, 365–372. [Google Scholar] [CrossRef]
- Manso, T.; Bodas, R.; Vieira, C.; Mantecón, A.R.; Castro, T. Feeding vegetable oils to lactating ewes modifies the fatty acid profile of sucking lambs. Animal 2011, 5, 1659–1667. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Cortés, P.; Gallardo, B.; Mantecón, A.R.; Juárez, M.; De La Fuente, M.A.; Manso, T. Effects of different sources of fat (calcium soap of palm oil vs. extruded linseed) in lactating ewes’ diet on the fatty acid profile of their suckling lambs. Meat Sci. 2014, 96, 1304–1312. [Google Scholar] [CrossRef] [Green Version]
- Serra, A.; Conte, G.; Ciucci, F.; Bulleri, E.; Corrales-Retana, L.; Cappucci, A.; Buccioni, A.; Mele, M. Dietary linseed supplementation affects the fatty acid composition of the sn-2 position of triglycerides in sheep milk. J. Dairy Sci. 2018, 101, 6742–6751. [Google Scholar] [CrossRef] [Green Version]
- Sinanoglou, V.J.; Koutsouli, P.; Fotakis, C.; Sotiropoulou, G.; Cavouras, D.; Bizelis, I. Assessment of lactation stage and breed effect on sheep milk fatty acid profile and lipid quality indices. Dairy Sci. Technol. 2015, 95, 509–531. [Google Scholar] [CrossRef] [Green Version]
- Bouattour, M.A.; Casals, R.; Albanell, E.; Such, X.; Caja, G. Feeding soybean oil to dairy goats increases conjugated linoleic acid in milk. J. Dairy Sci. 2008, 91, 2399–2407. [Google Scholar] [CrossRef] [Green Version]
- Tsiplakou, E.; Zervas, G. The effect of fish and soybean oil inclusion in goat diet on their milk and plasma fatty acid profile. Livest. Sci. 2013, 155, 236–243. [Google Scholar] [CrossRef]
- Ollier, S.; Leroux, C.; de la Foye, A.; Bernard, L.; Rouel, J.; Chilliard, Y. Whole intact rapeseeds or sunflower oil in high-forage or high-concentrate diets affects milk yield, milk composition, and mammary gene expression profile in goats. J. Dairy Sci. 2009, 92, 5544–5560. [Google Scholar] [CrossRef] [PubMed]
- Mele, M.; Buccioni, A.; Petacchi, F.; Serra, A.; Banni, S.; Antongiovanni, M.; Secchiari, P. Effect of forage/concentrate ratio and soybean oil supplementation on milk yield, and composition from Sarda ewes. Anim. Res. 2006, 55, 273–285. [Google Scholar] [CrossRef]
- Maia, M.O.; Susin, I.; Ferreira, E.M.; Nolli, C.P.; Gentil, R.S.; Petrini, J. Perfil de ácidos graxos do leite de ovelhas Santa Inês alimentadas com óleos vegetaisin. In Proceedings of the Anais da 48a Reunião Anual da Sociedade Brasileira de Zootecnia, Socieda de Brasileira de Zootecnia, Belém, Brazil, 18–21 July 2011. [Google Scholar]
- Castro, T.; Manso, T.; Jimeno, V.; Del Alamo, M.; Mantecón, A.R. Effects of dietary sources of vegetable fats on performance of dairy ewes and conjugated linoleic acid (CLA) in milk. Small Rumin. Res. 2009, 84, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Mandal, G.P.; Patra, A.K. Effects of different vegetable oils on rumen fermentation and conjugated linoleic acid concentration in vitro. Vet. World 2017, 10, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Luo, J.; Zhang, W.; Sheng, H. Using safflower supplementation to improve the fatty acid profile in milk of dairy goat. Small Rumin. Res. 2015, 127, 68–73. [Google Scholar] [CrossRef]
- Mierliță, D. Effects of diets containing hemp seeds or hemp cake on fatty acid composition and oxidative stability of sheep milk. S. Afr. J. Anim. Sci. 2018, 48, 504–515. [Google Scholar] [CrossRef]
- Buccioni, A.; Serra, A.; Minieri, S.; Mannelli, F.; Cappucci, A.; Benvenuti, D.; Rapaccini, S.; Conte, G.; Mele, M. Milk production, composition, and milk fatty acid profile from grazing sheep fed diets supplemented with chestnut tannin extract and extruded linseed. Small Rumin. Res. 2015, 130, 200–207. [Google Scholar] [CrossRef]
- Buccioni, A.; Pauselli, M.; Minieri, S.; Roscini, V.; Mannelli, F.; Rapaccini, S.; Lupi, P.; Conte, G.; Serra, A.; Cappucci, A.; et al. Chestnut or quebracho tannins in the diet of grazing ewes supplemented with soybean oil: Effects on animal performances, blood parameters and fatty acid composition of plasma and milk lipids. Small Rumin. Res. 2017, 153, 23–30. [Google Scholar] [CrossRef]
- Lee, J.; Lee, H.; Kang, S.; Park, W. Fatty acid desaturases, polyunsaturated fatty acid regulation, and biotechnological advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Kitessa, S.M.; Gulati, S.K.; Ashes, J.R.; Fleck, E.; Scott, T.W.; Nichols, P.D. Utilization of fish oil in ruminants: II. Transfer of fish oil fatty acids into goats’ milk. Anim. Feed Sci. Technol. 2001, 89, 201–208. [Google Scholar] [CrossRef]
- Fougère, H.; Bernard, L. Effect of diets supplemented with starch and corn oil, marine algae, or hydrogenated palm oil on mammary lipogenic gene expression in cows and goats: A comparative study. J. Dairy Sci. 2019, 102, 768–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, O.C.; Ferraz, M.V.C., Jr.; Susin, I.; Gentil, R.S.; Polizel, D.M.; Ferreira, E.M.; Barroso, J.P.R.; Pires, A.V. Plasma and milk fatty acid profiles in goats fed diets supplemented with oils from soybean, linseed or fish. Small Rumin. Res. 2019, 170, 125–130. [Google Scholar] [CrossRef]
- Pajor, F.; Egerszegi, I.; Steiber, O.; Bodnár, Á.; Póti, P. Effect of marine algae supplementation on the fatty acid profile of milk of dairy goats kept indoor and on pasture. J. Anim. Feed Sci 2019, 28, 169–176. [Google Scholar] [CrossRef]
- Kitessa, S.M.; Peake, D.; Bencini, R.; Williams, A.J. Fish oil metabolism in ruminants: III. Transfer of n− 3 polyunsaturated fatty acids (PUFA) from tuna oil into sheep’s milk. Anim. Feed Sci. Technol. 2003, 108, 1–14. [Google Scholar] [CrossRef]
- Wachira, A.M.; Sinclair, L.A.; Wilkinson, R.G.; Hallett, K.; Enser, M.; Wood, J.D. Rumen biohydrogenation of n-3 polyunsaturated fatty acids and their effects on microbial efficiency and nutrient digestibility in sheep. J. Agric. Sci. 2000, 135, 419–428. [Google Scholar] [CrossRef]
- Póti, P.; Pajor, F.; Bodnár, Á.; Penksza, K.; Köles, P. Effect of micro-alga supplementation on goat and cow milk fatty acid composition. Chilean J. Agric. Res. 2015, 75, 259–263. [Google Scholar] [CrossRef]
- Toral, P.G.; Hervás, G.; Gómez-Cortés, P.; Frutos, P.; Juárez, M.; de La Fuente, M.A. Milk fatty acid profile and dairy sheep performance in response to diet supplementation with sunflower oil plus incremental levels of marine algae. J. Dairy Sci. 2010, 93, 1655–1667. [Google Scholar] [CrossRef]
- Toral, P.G.; Monahan, F.J.; Hervás, G.; Frutos, P.; Moloney, A.P. Modulating ruminal lipid metabolism to improve the fatty acid composition of meat and milk. Challenges and opportunities. Animal 2018, 12, s272–s281. [Google Scholar] [CrossRef] [Green Version]
- Bernard, L.; Toral, P.G.; Chilliard, Y. Comparison of mammary lipid metabolism in dairy cows and goats fed diets supplemented with starch, plant oil, or fish oil. J. Dairy Sci. 2017, 100, 9338–9351. [Google Scholar] [CrossRef] [Green Version]
- Novotná, K.; Fantová, M.; Nohejlová, L.; Borková, M.; Stádník, L.; Ducháček, J. Effect of Chlorella Vulgaris and Japonochytrium sp. microalgae supplementation on composition and fatty Acid acid profile of goat milk. Acta Univ. Agric. Silvic. Mendel. Brun. 2017, 65, 1585–1593. [Google Scholar] [CrossRef] [Green Version]
- Mozzon, M.; Frega, N.G.; Fronte, B.; Tocchini, M. Effect of dietary fish oil supplements on levels of n-3 polyunsaturated fatty acids, trans acids and conjugated linoleic acid in ewe milk. Food Technol. Biotechnol. 2002, 40, 213–220. [Google Scholar]
- Ferreira, E.M.; Pires, A.V.; Susin, I.; Maia, M.O.; Gentil, R.S.; Nolli, C.P. Perfil de ácidos graxos do leite de ovelhasrecebendoóleo de soja e óleo de peixecomo fontes de gordura suplementar. In Proceedings of the Anais da 48a Reunião Anual da Sociedade Brasileira de Zootecnia, Sociedade Brasileira de Zootecnia, Belém, Brazil, 18–21 July 2011. [Google Scholar]
- Bichi, E.; Hervás, G.; Toral, P.G.; Loor, J.J.; Frutos, P. Milk fat depression induced by dietary marine algae in dairy ewes: Persistency of milk fatty acid composition and animal performance responses. J. Dairy Sci. 2013, 96, 524–532. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K. Effects of essential oils on rumen fermentation, microbial ecology and ruminant production. Asian J. Anim. Vet. Adv 2011, 6, 416–428. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza-Marinucci, M.; Yu, Z. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci. Total Environ. 2016, 545, 556–568. [Google Scholar] [CrossRef]
- Boaventura Neto, O. Effect of the utilization of aromatic plants on diet utilization, milk production, parasitic load, and health status of dairy ewes. Ph.D. Thesis, University of Sassari, Sassari, Italy, 2014. Available online: https://core.ac.uk/download/pdf/19979891.pdf (accessed on 25 February 2020).
- Manca, M.G.; Boe, R.; Manca, R.; Decandia, M.; Acciaro, M.; Cannas, A. Profilo acidico di latte e liquido ruminale di pecore alimentate con piante aromatiche. In Proceedings of the 20th Congresso Nazionale Società Italiana di Patologia ed Allevamento degli Ovini e dei Caprini, Siracusa, Italy, 26–29 September 2012; pp. 26–29. [Google Scholar]
- Kotsampasi, B.; Tsiplakou, E.; Christodoulou, C.; Mavrommatis, A.; Mitsiopoulou, C.; Karaiskou, C.; Sossidou, E.; Fragioudakis, N.; Kpsomenos, I.; Bampidis, V.A.; et al. Effects of dietary orange peel essential oil supplementation on milk yield and composition, and blood and milk antioxidant status of dairy ewes. Anim. Feed Sci. Technol. 2018, 245, 20–31. [Google Scholar] [CrossRef]
- Boutoial, K.; Ferrandini, E.; Rovira, S.; García, V.; López, M.B. Effect of feeding goats with rosemary (Rosmarinus officinalis spp.) by-product on milk and cheese properties. Small Rumin. Res. 2013, 112, 147–153. [Google Scholar] [CrossRef]
- Miri, V.H.; Tyagi, A.K.; Ebrahimi, S.H.; Mohini, M. Effect of cumin (Cuminumcyminum) seed extract on milk fatty acid profile and methane emission in lactating goat. Small Rumin. Res. 2013, 113, 66–72. [Google Scholar] [CrossRef]
- Zhu, Z.; Hang, S.; Zhu, H.; Zhong, S.; Mao, S.; Zhu, W. Effects of garlic oil on milk fatty acid profile and lipogenesis-related gene expression in mammary gland of dairy goats. J. Sci. Food Agric. 2013, 93, 560–567. [Google Scholar] [CrossRef]
- Canaes, T.S.; Zanferari, F.; Maganhe, B.L.; Takiya, C.S.; Silva, T.H.; Del Valle, T.A.; Rennó, F.P. Increasing dietary levels of citral oil on nutrient total tract digestibility, ruminal fermentation, and milk composition in Saanen goats. Anim. Feed Sci. Technol. 2017, 229, 47–56. [Google Scholar] [CrossRef]
- Zoidis, E.; Poulopoulou, I.; Tsoufi, V.; Massouras, T.; Hadjigeorgiou, I. Effects of terpene administration on goats’ milk fatty acid profile and coagulation properties. Int. J. Dairy Technol. 2018, 71, 992–996. [Google Scholar] [CrossRef]
- Kholif, S.M.; Morsy, T.A.; Abdo, M.M.; Matloup, O.H.; Abu El-Ella, A.A. Effect of supplementing lactating goats rations with garlic, cinnamon or ginger oils on milk yield, milk composition and milk fatty acids profile. J. Life Sci. 2012, 4, 27–34. [Google Scholar] [CrossRef]
- Bravo, L. Polyphenols: Chemistry, dietary sources, metabolism, and nutritional significance. Nutr. Rev. 1998, 56, 317–333. [Google Scholar] [CrossRef]
- Jerónimo, E.; Pinheiro, C.; Lamy, E.; Dentinho, M.T.; Sales-Baptista, E.; Lopes, O.; Capela e Silva, F. Tannins in ruminant nutrition: Impact on animal performance and quality of edible products. In Tannins: Biochemistry, Food Sources and Nutritional Properties; Combs, C.A., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2016; pp. 121–168. [Google Scholar]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Jöbstl, E.; O’Connell, J.; Fairclough, J.P.A.; Williamson, M.P. Molecular model for astringency produced by polyphenol/protein interactions. Biomacromolecules 2004, 5, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Correddu, F.; Lunesu, M.F.; Buffa, G.; Atzori, A.S.; Nudda, A.; Battacone, G.; Pulina, G. Can agro-industrial by-products rich in polyphenols be advantageously used in the feeding and nutrition of dairy small ruminants? Animals 2020, 10, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Vaithiyanathan, S. Occurrence, nutritional significance and effect on animal productivity of tannins in tree leaves. Anim. Feed Sci. Technol. 1990, 30, 21–38. [Google Scholar] [CrossRef]
- Min, B.R.; Barry, T.N.; Attwood, G.T.; McNabb, W.C. The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: A review. Anim. Feed Sci. Technol. 2003, 106, 3–19. [Google Scholar] [CrossRef]
- Nudda, A.; Correddu, F.; Atzori, A.S.; Marzano, A.; Battacone, G.; Nicolussi, P.; Bonelli, P.; Pulina, G. Whole exhausted berries of Myrtus communis L. supplied to dairy ewes: Effects on milk production traits and blood metabolites. Small Rumin. Res. 2017, 155, 33–38. [Google Scholar] [CrossRef]
- Correddu, F.; Fancello, F.; Chessa, L.; Atzori, A.S.; Pulina, G.; Nudda, A. Effects of supplementation with exhausted myrtle berries on rumen function of dairy sheep. Small Rumin. Res. 2019, 170, 51–61. [Google Scholar] [CrossRef]
- Cabiddu, A.; Molle, G.; Decandia, M.; Spada, S.; Fiori, M.; Piredda, G.; Addis, M. Responses to condensed tannins of flowering sulla (Hedysarumcoronarium L.) grazed by dairy sheep: Part 2: Effects on milk fatty acid profile. Livest. Sci. 2009, 123, 230–240. [Google Scholar] [CrossRef]
- Vasta, V.; Yáñez-Ruiz, D.R.; Mele, M.; Serra, A.; Luciano, G.; Lanza, M.; Biondi, L.; Priolo, A. Bacterial and protozoal communities and fatty acid profile in the rumen of sheep fed a diet containing added tannins. Appl. Environ. Microbiol. 2010, 76, 2549–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buccioni, A.; Pauselli, M.; Viti, C.; Minieri, S.; Pallara, G.; Roscini, V.; Rapaccini, S.; Trabalza Marinucci, M.; Lupi, P.; Conte, G.; et al. Milk fatty acid composition, rumen microbial population, and animal performances in response to diets rich in linoleic acid supplemented with chestnut or quebracho tannins in dairy ewes. J. Dairy Sci. 2015, 98, 1145–1156. [Google Scholar] [CrossRef] [Green Version]
- Abbeddou, S.; Rischkowsky, B.; Richter, E.K.; Hess, H.D.; Kreuzer, M. Modification of milk fatty acid composition by feeding forages and agro-industrial by products from dry areas to Awassi sheep. J. Dairy Sci. 2011, 94, 4657–4668. [Google Scholar] [CrossRef]
- Toral, P.G.; Hervás, G.; Bichi, E.; Belenguer, Á.; Frutos, P. Tannins as feed additives to modulate ruminal biohydrogenation: Effects on animal performance, milk fatty acid composition and ruminal fermentation in dairy ewes fed a diet containing sunflower oil. Anim. Feed Sci. Technol. 2011, 164, 199–206. [Google Scholar] [CrossRef]
- Toral, P.G.; Hervás, G.; Belenguer, A.; Bichi, E.; Frutos, P. Effect of the inclusion of quebracho tannins in a diet rich in linoleic acid on milk fatty acid composition in dairy ewes. J. Dairy Sci. 2013, 96, 431–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappucci, A.; Alves, S.P.; Bessa, R.J.B.; Buccioni, A.; Mannelli, F.; Pauselli, M.; Viti, C.; Pastorelli, R.; Roscini, V.; Serra, A.; et al. Effect of increasing amounts of olive crude phenolic concentrate in the diet of dairy ewes on rumen liquor and milk fatty acid composition. J. Dairy Sci. 2018, 101, 4992–5005. [Google Scholar] [CrossRef]
- Abo-Donia, F.M.; Yang, L.Y.; Hristov, A.N.; Wang, M.; Tang, S.X.; Zhou, C.S.; Han, X.F.; Kang, J.H.; Tan, Z.L.; He, Z.X. Effects of tannins on the fatty acid profiles of rumen fluids and milk from lactating goats fed a total mixed ration containing rape seed oil. Livest Sci 2017, 204, 16–24. [Google Scholar] [CrossRef]
- Alipanahi, Z.; Fatahnia, F.; Jafari, H.; Taasoli, G.; Mirzaei-Alamouti, H.; Barrett, D.; Pormalekshahi, A. Effect of oak acorn with or without polyethylene glycol in diets containing extruded soybean on milk fatty acid profile, ruminal fermentation and plasma metabolites of lactating goats. Livest. Sci. 2019, 221, 57–62. [Google Scholar] [CrossRef]
- de Lucena, A.R.F.; Menezes, D.R.; de Carvalho, D.T.Q.; Matos, J.C.; Antonelli, A.C.; de Moraes, S.A.; Dias, F.S.; Queiroz, M.A.A.; Rodrigues, R.T.S. Effect of commercial tannin and a pornunça (Manihot spp.) silage-based diet on the fatty acid profile of Saanen goats’ milk. Int. J. Dairy Technol. 2018, 71, 613–620. [Google Scholar] [CrossRef]
- de Carvalho, I.D.N.O.; da Fonseca, C.E.M.; Lopes, F.C.F.; Morenz, M.J.F.; da Gama, M.A.S.; de Souza, V.C.; da Silva, A.B. Milk fatty acid composition of dairy goats fed increasing levels of Flemingia macrophylla hay. Semin. Cienc. Agrar. 2019, 40, 293–310. [Google Scholar] [CrossRef] [Green Version]
- Emami, A.; Ganjkhanlou, M.; Nasri, M.F.; Zali, A.; Rashidi, L.; Sharifi, M. Antioxidant status of dairy goats fed diets containing pomegranate seed oil or linseed oil. Small Rumin. Res. 2017, 153, 175–179. [Google Scholar] [CrossRef]
- Razzaghi, A.; Naserian, A.A.; Valizadeh, R.; Ebrahimi, S.H.; Khorrami, B.; Malekkhahi, M.; Khiaosa-ard, R. Pomegranate seed pulp, pistachio hulls, and tomato pomace as replacement of wheat bran increased milk conjugated linoleic acid concentrations without adverse effects on ruminal fermentation and performance of Saanen dairy goats. Anim. Feed Sci. Technol. 2015, 210, 46–55. [Google Scholar] [CrossRef]
- Priolo, A.; Bella, M.; Lanza, M.; Galofaro, V.; Biondi, L.; Barbagallo, D.; Ben Salem, H.; Pennisi, P. Carcass and meat quality of lambs fed fresh sulla (Hedysarumcoronarium L.) with or without polyethylene glycol or concentrate. Small Rumin. Res. 2005, 59, 281–288. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Matitashvili, E.; Corl, B.A.; Dwyer, D.A.; Bauman, D.E. Trans-10, cis-12 conjugated linoleic acid decreases lipogenic rates and expression of genes involved in milk lipid synthesis in dairy cows. J. Dairy Sci. 2002, 85, 2155–2163. [Google Scholar] [CrossRef] [Green Version]
- Shingfield, K.J.; Bonnet, M.; Scollan, N.D. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7, 132–162. [Google Scholar] [CrossRef]
- Bauman, D.E.; Davis, C.L. Biosynthesis of milk fat. In Lactation: A Comprehensive Treatise; Larson, B.L., Ed.; Academic Press: New York, NY, USA, 1974; Volume 2, pp. 31–75. [Google Scholar]
- Sessler, A.M.; Ntambi, J.M. Polyunsaturated fatty acid regulation of gene expression. J. Nutr. 1998, 128, 923–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasta, V.; Priolo, A.; Scerra, M.; Hallett, K.G.; Wood, J.D.; Doran, O. Δ9 desaturase protein expression and fatty acid composition of longissimus dorsi muscle in lambs fed green herbage or concentrate with or without added tannins. Meat Sci. 2009, 82, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Tsiplakou, E.; Flemetakis, E.; Kalloniati, C.; Papadomichelakis, G.; Katinakis, P.; Zervas, G. Sheep and goats differences in CLA and fatty acids milk fat content in relation with mRNA stearoyl-CoA desaturase and lipogenic genes expression in their mammary gland. J. Dairy Res. 2009, 76, 392–401. [Google Scholar] [CrossRef]
- O’Connell, J.E.; Fox, P.F. Significance and applications of phenolic compounds in the production and quality of milk and dairy products: A review. Int. Dairy J. 2001, 11, 103–120. [Google Scholar] [CrossRef]
- Di Trana, A.; Bonanno, A.; Cecchini, S.; Giorgio, D.; Di Grigoli, A.; Claps, S. Effects of Sulla forage (Sulla coronarium L.) on the oxidative status and milk polyphenol content in goats. J. Dairy Sci. 2015, 98, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Correddu, F.; Nudda, A.; Manca, M.G.; Pulina, G.; Dalsgaard, T.K. Light-induced lipid oxidation in shee p milk: Effects of dietary grape seed and linseed, alone or in combination, on milk oxidative stability. J. Agric. Food Chem. 2015, 63, 3980–3986. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.W.; Santos, G.T.D.; Silva-Kazama, D.C.; Grande, P.A.; Pintro, P.M.; de Marchi, F.E.; Jobim, C.C.; Petit, H.V. Production, composition, and antioxidants in milk of dairy cows fed diets containing soybean oil and grape residue silage. Livest. Sci. 2014, 159, 37–45. [Google Scholar] [CrossRef]





| Species | Dose | Form 2 | Fat/d | Milk Fatty Acids (%) 1 | References | ||||
|---|---|---|---|---|---|---|---|---|---|
| RA | VA | LA | ALA | n6/n3 | |||||
| Goats | 100 g/d | EL | 17 g/d | 52 | 76 | 3 | 78 | −42 | [35] |
| 200 g/d | EL | 34 g/d | 67 | 99 | 3 | 120 | −53 | [35] | |
| 160 g/d | EL | 56 g/d | 112 | 148 | 13 | 132 | −86 | [56] | |
| 200 g/d | EL + P | 70 g/d | 55 | 100 | 1 | 145 | −59 | [36] | |
| 47 g/d | Oil | 47 g/d | 100 | 26 | 12 | 98 | −44 | [57] | |
| 360 + 40 g/d | EL | 126 g/d | 103 | 187 | 5 | 288 | −73 | [58] | |
| 3.4% DMI 3 | Oil | 102 g/d | 133 | 190 | 0 | 325 | −76 | [30] | |
| 130 g/d 4 | Oil | 130 g/d | 300 | 439 | −38 | 11 | −44 | [59] | |
| 130 g/d 5 | Oil | 130 g/d | 213 | 358 | −21 | 263 | −78 | [59] | |
| 511 g/d | EL | 197 g/d | 690 | 1268 | −9 | 415 | −82 | [60] | |
| 30 g/d 6 | Oil | 30 g/d | 48 | 75 | −2 | 89 | −48 | [61] | |
| 30 g/d 7 | Oil | 30 g/d | 135 | 183 | −13 | 102 | −57 | [61] | |
| Sheep | 210 g/d | EL | 79 g/d | 50 | 67 | 52 | 100 | −31 | [62] |
| 200 g/d | EL | 70 g/d | 213 | 294 | 11 | 153 | −56 | [63] | |
| 100 g/d | EL + P | – | 28 | 42 | 245 | 48 | 134 | [64] | |
| 63−70 g/d | Oil | 63 g/d | 308 | 447 | −12 | 106 | −57 | [65] | |
| 29 g/d | EL | 11 g/d | 9 | 30 | 0 | 14 | −13 | [66] | |
| 58 g/d | EL | 22 g/d | 18 | 65 | 9 | 21 | −10 | [66] | |
| 84 g/d | EL | 32 g/d | 73 | 83 | 9 | 43 | −24 | [66] | |
| 210 g/d | EL | 74 g/d | 247 | 273 | 0 | 168 | −63 | [67] | |
| 55 g/d | Oil | 55 g/d | 239 | 251 | −6 | 103 | −54 | [68] | |
| 128 g/d | Oil | 45 g/d | 141 | 256 | −25 | 202 | −75 | [69] | |
| 200 g/d | EL | 70 g/d | 74.84 | 122 | −3 | 43 | −33 | [70] | |
| Species | Dose | Form 2 | Milk Fatty Acids (%) 1 | References | ||||
|---|---|---|---|---|---|---|---|---|
| RA | VA | LA | ALA | n6/n3 | ||||
| Goats | 60 g/d | SO | 199 | 214 | 14 | −10 | 30 | [72] |
| 55 + 11 g/d | SO + FO | 609 | 466 | 34 | 13 | 15 | [73] | |
| SW | −33 | −10 | 50 | 0 | 24 | [30] | ||
| SUN‒oil | 283 | 290 | 55 | 25 | 9 | [30] | ||
| 130 g/d | SUN‒oil | 351 | 472 | 23 | −15 | 934 | [74] | |
| 130 g/d | SUN‒oil | 384 | 554 | 16 | −41 | 226 | [59] | |
| Sheep | 100 g/d | SO | 316 | 803 | 15 | −38 | [75] | |
| 63‒70 g/d | SO | 562 | 736 | 59 | 2 | 54 | [65] | |
| 60‒165 g/d | SO | 171 | 242 | 13 | −23 | 294 | [51] | |
| 182 g/d | SUN—seeds | 130 | 67 | 78 | 56 | 25 | [62] | |
| 65 g/d | SUN—oil | 138 | 195 | 6 | −12 | 21 | [76] | |
| 28.8 g/d | SUN—oil | 29 | 36 | 12 | 31 | −14 | [77] | |
| Specie | Polyphenol Source | Dietary TP 2 | Milk Fatty Acids (%) 1 | References | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SFA | MUFA | PUFA | SA | VA | RA | LA | ALA | ||||
| Sheep | Flowering sulla | 40.0 | −4 | 3 | 11 | −29 | −29 | 18 | 45 | [118] | |
| Lentil straw | 13.2 | −3 | 11 | 12 | −7 | 25 | 45 | 9 | 79 | [121] | |
| Olive leaves | 22.5 | −1 | −1 | 16 | −35 | 48 | 66 | 6 | 197 | [121] | |
| Quebracho and Chestnut extracts | 10.0 | −1 | −3 | −1 | 4 | 2 | −4 | 4 | 6 | [122] | |
| Quebracho extract | 20.0 | −2 | 7 | 3 | 3 | 12 | 8 | 2 | 2 | [123] | |
| Quebracho extract | 39.6 | −3 | 4 | 15 | −11 | 14 | 24 | 9 | 16 | [120] | |
| Chestnut extract | 24.3 | 0 | 0 | 1 | 9 | 7 | −2 | 3 | 3 | [120] | |
| Quebracho extract | 39.6 | −3 | 4 | 15 | −11 | 14 | 24 | 9 | 16 | [82] | |
| Chestnut extract | 24.3 | 0 | 0 | 1 | 9 | 7 | −2 | 3 | 3 | [82] | |
| Olive crude phenolic concentrate | 0.6 | −6 | 26 | 21 | 7 | −4 | [124] | ||||
| Olive crude phenolic concentrate | 0.8 | 0 | −19 | −17 | 16 | 16 | [124] | ||||
| Olive crude phenolic concentrate | 1.2 | 9 | −22 | −26 | 18 | 24 | [124] | ||||
| Goat | Gallnut | 3.0 | −4 | 5 | 5 | −2 | 24 | 24 | −5 | [125] | |
| Gallnut | 6.0 | −4 | 6 | 15 | 11 | 25 | 20 | 12 | [125] | ||
| Gallnut | 9.0 | −5 | 8 | 19 | 7 | 22 | 18 | 18 | [125] | ||
| Oak acorn | 12.0 | −25 | 48 | 68 | 13 | 22 | [126] | ||||
| Mimosa extract | 24.0 | −1 | 1 | 11 | 11 | 20 | 5 | 13 | [127] | ||
| Mimosa extract | 36.0 | −2 | 3 | 24 | 16 | 20 | −2 | 35 | [127] | ||
| Mimosa extract | 48.0 | −5 | 8 | 38 | 15 | 30 | 7 | 48 | [127] | ||
| Flemingia hay | 0.8 | 4 | 14 | 16 | 9 | 16 | [128] | ||||
| Flemingia hay | 1.7 | 8 | 31 | 38 | 23 | 33 | [128] | ||||
| Flemingia hay | 2.5 | −5 | 23 | 29 | 9 | 14 | [128] | ||||
| Flemingia hay | 3.4 | 14 | 77 | 62 | 20 | 32 | [128] | ||||
| Pomegranate seed oil | 3.2 | −5 | 7 | 39 | 1 | 37 | 131 | 8 | 110 | [57,129] | |
| Pomegranate seed pulp | 4.7 | −1 | 4 | 31 | 13 | 121 | 562 | [130] | |||
| Pistachio hulls | 26.6 | 1 | 10 | −1 | −1 | 104 | 431 | [130] | |||
| Tomato pomace | 3.2 | −5 | 24 | −3 | 14 | 100 | 400 | [130] | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nudda, A.; Cannas, A.; Correddu, F.; Atzori, A.S.; Lunesu, M.F.; Battacone, G.; Pulina, G. Sheep and Goats Respond Differently to Feeding Strategies Directed to Improve the Fatty Acid Profile of Milk Fat. Animals 2020, 10, 1290. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10081290
Nudda A, Cannas A, Correddu F, Atzori AS, Lunesu MF, Battacone G, Pulina G. Sheep and Goats Respond Differently to Feeding Strategies Directed to Improve the Fatty Acid Profile of Milk Fat. Animals. 2020; 10(8):1290. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10081290
Chicago/Turabian StyleNudda, Anna, Antonello Cannas, Fabio Correddu, Alberto Stanislao Atzori, Mondina Francesca Lunesu, Gianni Battacone, and Giuseppe Pulina. 2020. "Sheep and Goats Respond Differently to Feeding Strategies Directed to Improve the Fatty Acid Profile of Milk Fat" Animals 10, no. 8: 1290. https://0-doi-org.brum.beds.ac.uk/10.3390/ani10081290