
Uncoupling Protein-1 (UCP1) in the Adult Horse: Correlations with Body Weight, Rectal Temperature and Lipid Profile

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design.
2.2. Sampling Procedures and Laboratory Analysis
2.3. Statistical analysis.
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azzu, V.; Brand, M. The on-off switches of the mitochondrial uncoupling proteins. Trends Biochem. Sci. 2010, 35, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Dalgaard, L.T.; Pedersen, O. Uncoupling proteins: Functional characteristics and role in the pathogenesis of obesity and Type II diabetes. Diabetologia 2001, 44, 946–965. [Google Scholar] [CrossRef] [Green Version]
- Lowell, B.B.; Spiegelman, B.M. Towards a molecular understanding of adaptive thermogenesis. Nature 2000, 404, 652–660. [Google Scholar] [CrossRef]
- Virtanen, K.; Nuutila, P. Brown adipose tissue in humans. Curr. Opin. Lipidol. 2011, 22, 49–54. [Google Scholar] [CrossRef]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.-J.; Enerbäck, S.; et al. Functional Brown Adipose Tissue in Healthy Adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef]
- Bouillaud, F.; Ricquier, D.; Tiraby, C.; Tavernier, G.; Lefort, C.; Larrouy, D.; Langin, D. Acquirement of brown fat cell features by hu-man white adipocytes. J. Biol. Chem. 2003, 278, 33370–33376. [Google Scholar]
- Coskun, T.; Bina, H.A.; Schneider, M.A.; Dunbar, J.D.; Hu, C.C.; Chen, Y.; Moller, D.E.; Kharitonenkov, A. Fibroblast growth factor 21 cor-rects obesity in mice. Endocrinology 2008, 149, 6018–6027. [Google Scholar] [CrossRef]
- Keipert, S.; Ost, M.; Johann, K.; Imber, F.; Jastroch, M.; Van Schothorst, E.M.; Keijer, J.; Klaus, S. Skeletal muscle mitochondrial uncoupling drives endocrine cross-talk through the induction of FGF21 as a myokine. Am. J. Physiol. Metab. 2014, 306, E469–E482. [Google Scholar] [CrossRef] [PubMed]
- Fisher, F.M.; Kleiner, S.; Douris, N.; Fox, E.C.; Mepani, R.J.; Verdeguer, F.; Wu, J.; Kharitonenkov, A.; Flier, J.S.; Maratos-Flier, E.; et al. FGF21 regulates PGC-1 and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012, 26, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Kharitonenkov, A.; Wroblewski, V.J.; Koester, A.; Chen, Y.-F.; Clutinger, C.K.; Tigno, X.T.; Hansen, B.C.; Shanafelt, A.B.; Etgen, G.J. The Metabolic State of Diabetic Monkeys Is Regulated by Fibroblast Growth Factor-21. Endocrinology 2007, 148, 774–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabalina, I.G.; Petrovic, N.; de Jong, J.M.; Kalinovich, A.V.; Cannon, B.; Nedergaard, J. UCP1 in brite/beige adi-pose tissue mitochondria is functionally thermogenic. Cell Rep. 2013, 5, e1196–e1203. [Google Scholar] [CrossRef] [Green Version]
- Keipert, S.; Jastroch, M. Brite/beige fat and UCP1-is it thermogenesis? Biochim. Biophys. Acta 2014, 1837, e1075–e1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arfuso, F.; Rizzo, M.; Giannetto, C.; Giudice, E.; Fazio, F.; Piccione, G. Age-related changes of serum mitochondrial uncoupling 1, rumen and rectal temperature in goats. J. Therm. Biol. 2016, 59, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Giannetto, C.; Rizzo, M.; Fazio, F.; Giudice, E.; Piccione, G. Serum levels of mitochondrial uncoupling protein 1, leptin, and lipids during late pregnancy and the early postpartum period in mares. Theriogenology 2016, 86, 1156–1164. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Woods, S.C.; Porte, D., Jr.; Seeley, R.J.; Baskin, D.G. Central nervous system control of food intake. Nature 2000, 404, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Giacobino, J.P. Uncoupling proteins, leptin, and obesity: An updated review. Ann. N. Y. Acad. Sci. 2002, 967, 398–402. [Google Scholar] [CrossRef]
- Reidy, S.P.; Weber, J.-M. Accelerated substrate cycling: A new energy-wasting role for leptin in vivo. Am. J. Physiol. Metab. 2002, 282, E312–E317. [Google Scholar] [CrossRef] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredricson, D.S. Estimation of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Brondani, L.D.A.; Assmann, T.S.; Duarte, G.C.K.; Gross, J.L.; Canani, L.H.; Crispim, D. The role of the uncoupling protein 1 (UCP1) on the development of obesity and type 2 diabetes mellitus. Arq. Bras. Endocrinol. Metabol. 2012, 56, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [PubMed] [Green Version]
- Si, Y.; Palani, S.; Jayaraman, A.; Lee, K. Effects of forced uncoupling protein 1 expression in 3T3-L1 cells on mitochondrial function and lipid metabolism. J. Lipid Res. 2007, 48, 826–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozak, L.P.; Anunciado-Koza, R. UCP1: Its involvement and utility in obesity. Int. J. Obes. 2008, 32 (Suppl. 7), S32–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannetto, C.; Arfuso, F.; Giudice, E.; Di Pietro, S.; Piccione, G. Clock gene per 2 daily rhythm: Correlation with the serum level of uncoupling protein 1 (UCP1) in goat and horse. J. Therm. Biol. 2021, 97, 102891. [Google Scholar] [CrossRef] [PubMed]
- Arechaga, I.; Ledesma, A.; Rial, E. The Mitochondrial Uncoupling Protein UCP1: A Gated Pore. IUBMB Life 2001, 52, 165–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.J. Recent advances in regulating energy homeostasis and obesity. Korean J. Pediatr. 2005, 48, 126–137. [Google Scholar]
- Nagai, N.; Sakane, N.; Kotani, K.; Hamada, T.; Tsuzaki, K.; Moritani, T. Uncoupling protein 1 gene −3826 A/G polymorphism is associated with weight loss on a short-term, controlled-energy diet in young women. Nutr. Res. 2011, 31, 255–261. [Google Scholar] [CrossRef]
- Kim-Motoyama, H.; Yasuda, K.; Yamaguchi, T.; Yamada, N.; Katakura, T.; Shuldiner, A.R.; Akanuma, Y.; Ohashi, Y.; Yazaki, Y.; Kadowaki, T. A mutation of the beta 3-adrenergic receptor is associated with visceral obesity but decreased serum triglyceride. Diabetologia 1997, 40, 469–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esterbauer, H.; Oberkofler, H.; Liu, Y.M.; Breban, D.; Hell, E.; Krempler, F.; Patsch, W. Uncoupling protein-1 mRNA expression in obese human subjects: The role of sequence variations at the uncoupling protein-1 gene locus. J. Lipid Res. 1998, 39, 834–844. [Google Scholar] [CrossRef]
- Oh, H.H.; Kim, K.S.; Choi, S.M.; Yang, H.S.; Yoon, Y. The effects of uncoupling protein-1 genotype on lipoprotein cholesterol lev-el in Korean obese subjects. Metabolism 2004, 53, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Kotani, K.; Sakane, N.; Kurozawa, Y.; Kaetsu, A.; Okamoto, M.; Osaki, Y.; Kishimoto, T. Polymorphism of Trp64Arg in beta3-adrenergic receptor gene and serum LDL-cholesterol concentrations in healthy Japanese. Ann. Clin. Biochem. 2008, 45, 313–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushita, H.; Kurabayashi, T.; Tomita, M.; Kato, N.; Tanaka, K. Effects of uncoupling protein 1 and β3-adrenergic receptor gene polymorphism on body size and serum lipid concentrations in Japanese women. Maturitas 2003, 45, 39–45. [Google Scholar] [CrossRef]
- Schäffler, A.; Palitzsch, K.D.; Watzlawek, E.; Drobnik, W.; Schwer, H.; Schölmerich, J.; Schmitz, G. Frequency and significance of the A→G (−3826) polymorphism in the promoter of the gene for uncoupling protein-1 with regard to metabolic parameters and adipocyte transcription factor binding in a large population-based Caucasian cohort. Eur. J. Clin. Investig. 1999, 29, 770–779. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
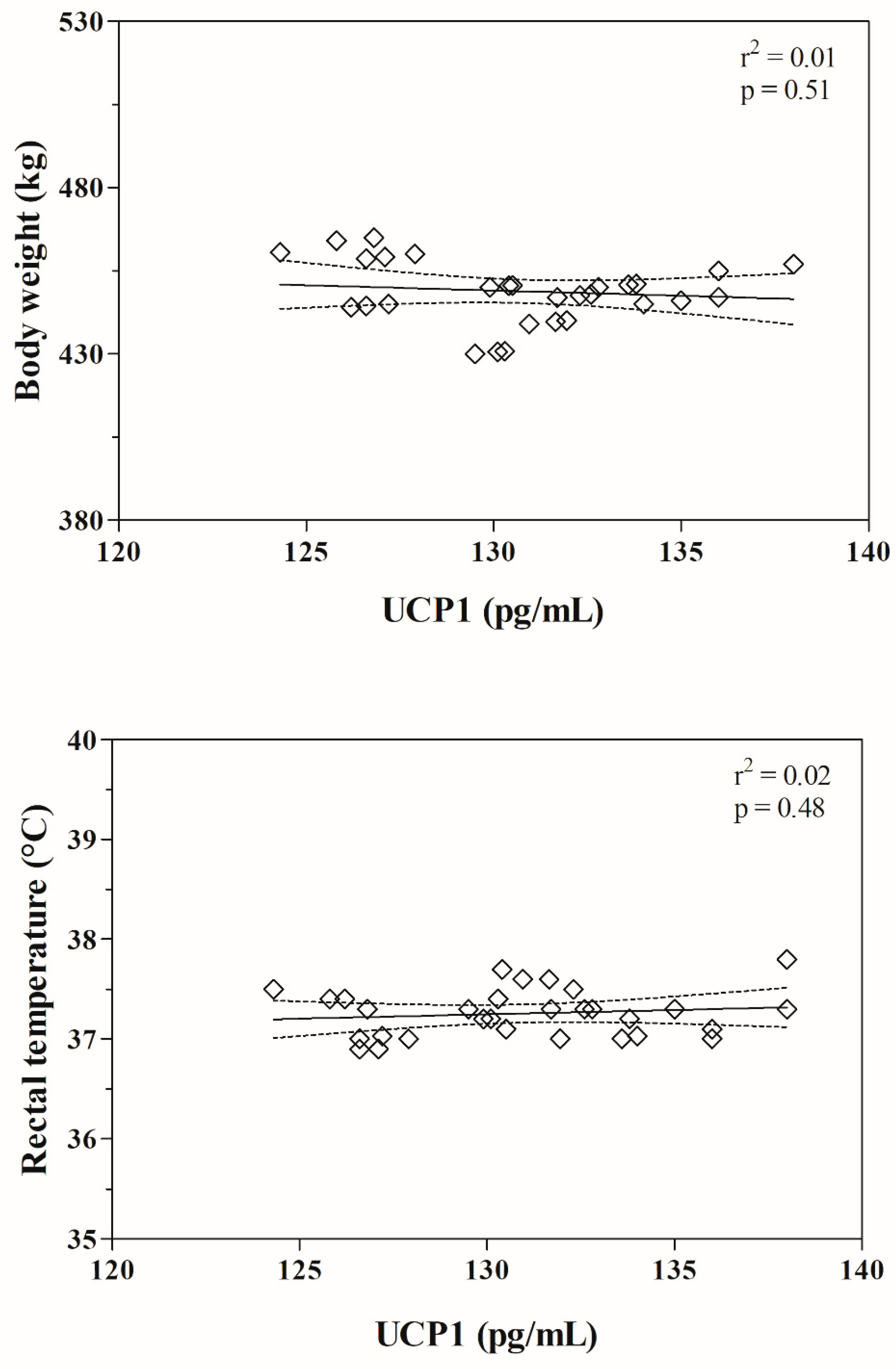
| Parameters | Body Weight (kg) | Rectal Temperature (°C) | ||||||
| UCP1 (pg/mL) | r = −0.12 p = 0.51 | r = 0.13 p = 0.48 | ||||||
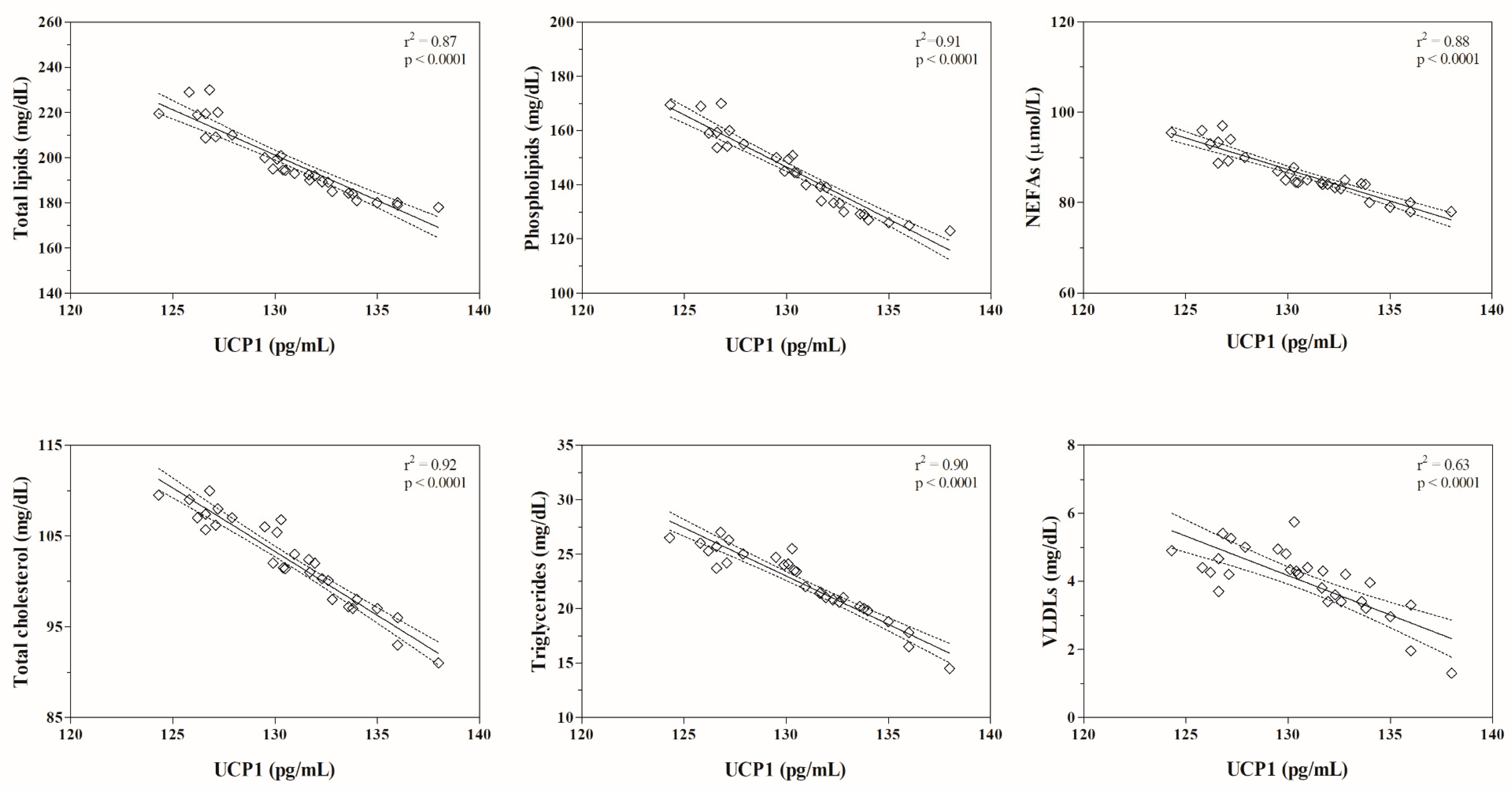
| Total Lipids (mg/dL) | Phospholipids (mg/dL) | NEFAs (μmol/L) | Total Cholesterol (mg/dL) | HDLs (mg/dL) | LDLs (mg/dL) | Triglycerides (mg/dL) | VLDLs (mg/dL) | |
| UCP1 (pg/mL) | r = −0.93 p < 0.0001 | r = −0.96 p < 0.0001 | r = −0.94 p < 0.0001 | r = −0.96 p < 0.0001 | r = −0.11 p = 0.57 | r = −0.47 p = 0.47 | r = −0.95 p < 0.0001 | r = −0.79 p < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arfuso, F.; Giannetto, C.; Panzera, M.F.; Fazio, F.; Piccione, G. Uncoupling Protein-1 (UCP1) in the Adult Horse: Correlations with Body Weight, Rectal Temperature and Lipid Profile. Animals 2021, 11, 1836. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11061836
Arfuso F, Giannetto C, Panzera MF, Fazio F, Piccione G. Uncoupling Protein-1 (UCP1) in the Adult Horse: Correlations with Body Weight, Rectal Temperature and Lipid Profile. Animals. 2021; 11(6):1836. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11061836
Chicago/Turabian StyleArfuso, Francesca, Claudia Giannetto, Maria Francesca Panzera, Francesco Fazio, and Giuseppe Piccione. 2021. "Uncoupling Protein-1 (UCP1) in the Adult Horse: Correlations with Body Weight, Rectal Temperature and Lipid Profile" Animals 11, no. 6: 1836. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11061836