
Artificial Termite-Fishing Tasks as Enrichment for Sanctuary-Housed Chimpanzees: Behavioral Effects and Impact on Welfare

 ,
,  ,
,  , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Study Site
2.2. Experimental Procedure
2.3. Behavioral Observations
2.3.1. Treatment Phase: Baseline and Enrichment Conditions
2.3.2. Pre- and Post-Treatment Phases
2.3.3. Rank Calculation
2.3.4. Inter-Observer Reliability
2.4. Data Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Committee, E.C.V.S. The Welfare of Intensively Kept Pigs: Report of the Scientific Veterinary Committee Adopted 30 September 1997; European Commission: Brussels, Belgium, 1997. [Google Scholar]
- Harrison, R. Animal Machines; Vincent Stuart Ltd.: London, UK, 1964. [Google Scholar]
- Cole, J.; Fraser, D. Zoo Animal Welfare: The Human Dimension. J. Appl. Anim. Welf. Sci. 2018, 21, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagan, R.; Carter, S.; Allard, S. A Universal Animal Welfare Framework for Zoos. J. Appl. Anim. Welf. Sci. 2015, 18, S1–S10. [Google Scholar] [CrossRef] [Green Version]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S. Assessment of welfare in zoo animals: Towards optimum quality of life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, P.; Riley, L. The use of Qualitative Behavioural Assessment to zoo welfare measurement and animal husbandry change. J. Zoo Aquar. Res. 2019, 74, 150–161. [Google Scholar]
- Learmonth, M.J. Dilemmas for natural living concepts of zoo animal welfare. Animals 2019, 9, 318. [Google Scholar] [CrossRef] [Green Version]
- Maple, T.L.; Bloomsmith, M.A. Introduction: The science and practice of optimal animal welfare. Behav. Process. 2018, 156, 1–2. [Google Scholar] [CrossRef]
- Mellor, D.; Hunt, S.; Gusset, M. Caring for Wildlife: The World Zoo and Aquarium Animal Welfare Strategy; WAZA Executive Office: Gland, Switzerland, 2015. [Google Scholar]
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in applied zoo animal welfare science. J. Appl. Anim. Welf. Sci. 2018, 21, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broom, D.M. Animal welfare: Concepts and measurement. J. Anim. Sci. 1991, 69, 4167–4175. [Google Scholar] [CrossRef] [PubMed]
- Maple, T.L.; Perdue, B.M. Environmental Enrichment. In Zoo Animal Welfare; Springer: Berlin/Heidelberg, Germany, 2013; pp. 95–117. [Google Scholar]
- Mellor, D.J. Updating Animal Welfare Thinking: Moving beyond the “Five Freedoms” towards “A Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef]
- Brambell, F.W.R. Report of the Technical Committee to Enquire into the Welfare of Animals Kept under Intensive Livestock Husbandry Systems; Her Majesty’s Stationery Office: London, UK, 1965. [Google Scholar]
- Kiley-Worthington, M. Ecological, ethological, and ethically sound environments for animals: Toward symbiosis. J. Agric. Environ. Ethics 1989, 2, 323–347. [Google Scholar] [CrossRef]
- Bracke, M.B.M.; Hopster, H. Assessing the Importance of Natural Behavior for Animal Welfare. J. Agric. Environ. Ethics 2006, 19, 77–89. [Google Scholar] [CrossRef]
- Hughes, B.O.; Duncan, I.J.H. The notion of ethological ‘need’, models of motivation and animal welfare. Anim. Behav. 1988, 36, 1696–1707. [Google Scholar] [CrossRef]
- Young, R. Environmental Enrichment for Captive Animals; Blackwell Publishing: Oxford, UK, 2003. [Google Scholar]
- Mallapur, A. Animal Welfare research and its implications to non-human primate breeding programs: A Case Study of the Lion-tailed macaque conservation breeding program from India. REDVET 2008, 9, 1–21. [Google Scholar]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef]
- Bain, M.; Buffington, C.T. The Relationship between Mental and Physical Health; Department of Medicine and Epidemiology, School of Veterinary Medicine: UC Davis, CA, USA, 2019; p. 33. [Google Scholar]
- Hill, S.P.; Broom, D.M. Measuring zoo animal welfare: Theory and practice. Zoo Biol. 2009, 28, 531–544. [Google Scholar] [CrossRef]
- Lutz, C.K.; Novak, M.A. Environmental enrichment for nonhuman primates: Theory and application. ILAR J. 2005, 46, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Dutton, M.B.; Pierre, P.J.; Bailoo, J.; Warkins, E.; Michel, G.F.; Bennett, A.J. A Model Quantitative Assessment Tool for Nonhuman Primate Environmental Enrichment Plans. bioRxiv 2018. [Google Scholar] [CrossRef]
- Bloomsmith, M.; Alford, P.; Maple, T. Successful feeding enrichment for captive chimpanzees. Am. J. Primatol. 1988, 16, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.C. Straw and forage material ameliorate abnormal behaviors in adult chimpanzees. Zoo Biol. 1997, 16, 225–236. [Google Scholar] [CrossRef]
- Maki, S.; Alford, P.L.; Bloomsmith, M.A.; Franklin, J. Food puzzle device simulating termite fishing for captive chimpanzees (Pan troglodytes). Am. J. Primatol. 1989, 1, 71–78. [Google Scholar]
- Celli, M.L.; Tomonaga, M.; Udono, T.; Teramoto, M.; Nagano, K. Tool use task as environmental enrichment for captive chimpanzees. Appl. Anim. Behav. Sci. 2003, 81, 171–182. [Google Scholar] [CrossRef]
- Celli, M.L.; Hirata, S.; Tomonaga, M. Socioecological influences on tool use in captive chimpanzees. Int. J. Primatol. 2004, 25, 1267–1281. [Google Scholar] [CrossRef]
- Nash, V.J. Tool use by captive chimpanzees at an artificial termite mound. Zoo Biol. 1982, 1, 211–221. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Matsunaga, M.; Shimada, K.; Kado, R.; Tanaka, M. Introducing tool-based feeders to zoo-housed chimpanzees as a cognitive challenge: Spontaneous acquisition of new types of tool use and effects on behaviours and use of space. J. Zoo Aquar. Res. 2016, 4, 147–155. [Google Scholar]
- Llorente, M.; Campi, A. Is “tool use” a welfare tool? Using an artificial ant-fishing task as environmental enrichment in naturalistically housed chimpanzees from the Fundació Mona. Folia Primatol. 2014, 85, 68. [Google Scholar]
- Goodall, J. The Chimpanzees of Gombe: Patterns of Behavior; Cambridge University Press: Cambridge, MA, USA, 1986. [Google Scholar]
- Boesch, C.; Boesch, H. Tool use and tool making in wild chimpanzees. Folia Primatol. 1990, 54, 86–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquette, D. Discovering and learning tool-use for fishing honey by captive chimpanzees. Hum. Evol. 1992, 7, 17–30. [Google Scholar] [CrossRef]
- Hirata, S.; Celli, M.L. Role of mothers in the acquisition of tool-use behaviours by captive infant chimpanzees. Anim. Cogn. 2003, 6, 235–244. [Google Scholar] [CrossRef]
- Hirata, S.; Morimura, N. Naive chimpanzees’ (Pan troglodytes) observation of experienced conspecifics in a tool-using task. J. Comp. Psychol. 2000, 114, 291–296. [Google Scholar] [CrossRef]
- Hopper, L.M.; Tennie, C.; Ross, S.R.; Lonsdorf, E.V. Chimpanzees create and modify probe tools functionally: A study with zoo-housed chimpanzees. Am. J. Primatol. 2015, 77, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.A.; Whiten, A. Chimpanzees (Pan troglodytes) display limited behavioural flexibility when faced with a changing foraging task requiring tool use. PeerJ 2018, 6, e4366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrew, W.C.; Marchant, L.F. Chimpanzees, tools, and termites: Hand preference or handedness? Curr. Anthropol. 1992, 33, 114–119. [Google Scholar]
- Brent, L.; Eichberg, J.W. Primate puzzleboard: A simple environmental enrichment device for captive chimpanzees. Zoo Biol. 1991, 10, 353–360. [Google Scholar] [CrossRef]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A modest proposal: Displacement activities as an indicator of emotions in primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Clark, F.E.; Ith, L.J. Effect of a Cognitive Challenge Device Containing Food and Non-Food Rewards on Chimpanzee Well-Being. Am. J. Primatol. 2013, 75, 807–816. [Google Scholar] [CrossRef]
- Elder, C.M.; Menzel, C.R. Dissociation of cortisol and behavioral indicators of stress in an orangutan (Pongo pygmaeus) during a computerized task. Primates 2001, 42, 345–357. [Google Scholar] [CrossRef]
- Itakura, S. Emotional Behavior during the Learning of a Contingency Task in a Chimpanzee. Percept. Mot. Skills 1993, 76, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Leavens, D.A.; Aureli, F.; Hopkins, W.D.; Hyatt, C.W. Effects of cognitive challenge on self-directed behaviors by chimpanzees (Pan troglodytes). Am. J. Primatol. 2001, 55, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yamanashi, Y.; Matsuzawa, T. Emotional consequences when chimpanzees (Pan troglodytes) face challenges: Individual differences in self-directed behaviours during cognitive tasks. Anim. Welf. 2010, 19, 25–30. [Google Scholar]
- Krause, J.; Croft, D.P.; James, R. Social network theory in the behavioural sciences: Potential applications. Behav. Ecol. Sociobiol. 2007, 62, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.; Petit, O. Social Network Modeling: A Powerful Tool for the Study of Group Scale Phenomena in Primates. Am. J. Primatol. 2011, 73, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Kasper, C.; Voelkl, B. A social network analysis of primate groups. Primates 2009, 50, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, J.; Micheletta, J.; Powell, L.E.; Bordier, C.; Waller, B.M. The impact of cognitive testing on the welfare of group housed primates. PLoS ONE 2013, 8, e78308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, S.L.; Kwiatt, A.C.; Ross, S.R.; Cronin, K.A. The effects of cognitive testing on the welfare of zoo-housed Japanese macaques (Macaca fuscata). Appl. Anim. Behav. Sci. 2019, 212, 90–97. [Google Scholar] [CrossRef]
- Schapiro, S.J.; Lambeth, S.P. Control, Choice, and Assessments of the Value of Behavioral Management to Nonhuman Primates in Captivity. J. Appl. Anim. Welf. Sci. 2007, 10, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Hopper, L.M.; Price, S.A.; Freeman, H.D.; Lambeth, S.P.; Schapiro, S.J.; Kendal, R.L. Influence of personality, age, sex, and estrous state on chimpanzee problem-solving success. Anim. Cogn. 2014, 17, 835–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruetz, J.D.; Bertolani, P.; Ontl, K.B.; Lindshield, S.; Shelley, M.; Wessling, E.G. New evidence on the tool-assisted hunting exhibited by chimpanzees (Pan troglodytes verus) in a savannah habitat at Fongoli, Sénégal. R. Soc. Open. Sci. 2015, 2, 140507. [Google Scholar] [CrossRef] [Green Version]
- Boesch, C.; Boesch, H. Sex differences in the use of natural hammers by wild chimpanzees: A preliminary report. J. Hum. Evol. 1981, 10, 585–593. [Google Scholar] [CrossRef]
- McGrew, W. Evolutionary implications of sex differences in chimpanzee predation and tool use. In The Great Apes; Benjamin Staples: Menlo Park, CA, USA, 1979; pp. 441–463. [Google Scholar]
- Boose, K.J.; White, F.J.; Meinelt, A. Sex differences in tool use acquisition in bonobos (Pan paniscus). Am. J. Primatol. 2013, 75, 917–926. [Google Scholar] [CrossRef]
- Gruber, T.; Clay, Z.; Zuberbühler, K. A comparison of bonobo and chimpanzee tool use: Evidence for a female bias in the Pan lineage. Anim. Behav. 2010, 80, 1023–1033. [Google Scholar] [CrossRef] [Green Version]
- Bakeman, R.; Quera, V. Sequential Analysis and Observational Methods for the Behavioral Sciences; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Meyer, J.S.; Hamel, A.F. Models of stress in nonhuman primates and their relevance for human psychopathology and endocrine dysfunction. ILAR J. 2014, 55, 347–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schino, G.; Perretta, G.; Taglioni, A.M.; Monaco, V.; Troisi, A. Primate displacement activities as an ethopharmacological model of anxiety. Anxiety 1996, 2, 186–191. [Google Scholar] [CrossRef]
- Wagner, K.E.; Hopper, L.M.; Ross, S.R. Asymmetries in the production of self-directed behavior by chimpanzees and gorillas during a computerized cognitive test. Anim. Cogn. 2016, 19, 343–350. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Russell, J.L.; Freeman, H.; Reynolds, E.A.; Griffis, C.; Leavens, D.A. Lateralized scratching in chimpanzees (Pan troglodytes): Evidence of a functional asymmetry during arousal. Emotion 2006, 6, 553–559. [Google Scholar] [CrossRef] [Green Version]
- Crailsheim, D.; Stüger, H.P.; Kalcher-Sommersguter, E.; Llorente, M. Early life experience and alterations of group composition shape the social grooming networks of former pet and entertainment chimpanzees (Pan troglodytes). PLoS ONE 2020, 15, e0226947. [Google Scholar] [CrossRef] [PubMed]
- Llorente, M.; Riba, D.; Ballesta, S.; Feliu, O.; Rostán, C. Rehabilitation and socialization of chimpanzees (Pan troglodytes) used for entertainment and as pets: An 8-year study at Fundació Mona. Int. J. Primatol. 2015, 36, 605–624. [Google Scholar] [CrossRef]
- Ross, M.; Niemann, T.; Wark, J.; Heintz, M.; Horrigan, A.; Cronin, K.; Shender, M.; Gillespie, K. Zoomonitor (Version 1) [Mobile Application Software]. 2016. Available online: https://zoomonitor.org (accessed on 10 October 2021).
- Wark, J.; Cronin, K.; Niemann, T.; Shender, M.; Horrigan, A.; Kao, A.; Ross, M. Monitoring the behavior and habitat use of animals to enhance welfare using the ZooMonitor app. Anim. Behav. Cogn 2019, 6, 158–167. [Google Scholar] [CrossRef]
- Neumann, C.; Duboscq, J.; Dubuc, C.; Ginting, A.; Irwan, A.M.; Agil, M.; Widdig, A.; Engelhardt, A. Assessing dominance hierarchies: Validation and advantages of progressive evaluation with Elo-rating. Anim. Behav. 2011, 82, 911–921. [Google Scholar] [CrossRef]
- Sánchez-Tójar, A.; Schroeder, J.; Farine, D.R. A practical guide for inferring reliable dominance hierarchies and estimating their uncertainty. J. Anim. Ecol. 2018, 87, 594–608. [Google Scholar] [CrossRef] [PubMed]
- Baayen, R.H. Analyzing Linguistic Data: A Practical Introduction to Statistics Using R; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Dobson, A.J. An introduction to Generalized Linear Models; Chapman & Hall/CRC: Boca Raton, FL, USA, 2002. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means, R package version 1.4.7. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 10 October 2021).
- Field, A. Discovering Statistics Using SPSS; SAGE Publications: London, UK, 2009. [Google Scholar]
- Izzo, G.N.; Bashaw, M.J.; Campbell, J.B. Enrichment and individual differences affect welfare indicators in squirrel monkeys (Saimiri sciureus). J. Comp. Psychol. 2011, 125, 347. [Google Scholar] [CrossRef]
- Coleman, K.; Novak, M.A. Environmental enrichment in the 21st century. ILAR J. 2017, 58, 295–307. [Google Scholar] [CrossRef]
- Coleman, K. Individual differences in temperament and behavioral management practices for nonhuman primates. Appl. Anim. Behav. Sci. 2012, 137, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Altschul, D.; Wallace, E.; Sonnweber, R.; Tomonaga, M.; Weiss, A. Chimpanzee intellect: Personality, performance and motivation with touchscreen tasks. R. Soc. Open. Sci. 2017, 4, 170169. [Google Scholar] [CrossRef] [Green Version]
- Herrelko, E.S.; Vick, S.J.; Buchanan-Smith, H.M. Cognitive Research in Zoo-Housed Chimpanzees: Influence of Personality and Impact on Welfare. Am. J. Phys. Anthropol. 2012, 74, 828–840. [Google Scholar] [CrossRef] [PubMed]
- Reamer, L.A.; Haller, R.L.; Thiele, E.J.; Freeman, H.D.; Lambeth, S.P.; Schapiro, S.J. Factors affecting initial training success of blood glucose testing in captive chimpanzees (Pan troglodytes). Zoo Biol. 2014, 33, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Morton, B.F.; Lee, P.C.; Buchanan-Smith, H.M. Taking personality selection bias seriously in animal cognition research: A case study in capuchin monkeys (Sapajus apella). Anim. Cogn. 2013, 16, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Bard, K.A.; Bakeman, R.; Boysen, S.T.; Leavens, D.A. Emotional engagements predict and enhance social cognition in young chimpanzees. Dev. Sci. 2014, 17, 682–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, E.A.; Sclafani, V.; Paukner, A.; Kaburu, S.S.; Suomi, S.J.; Ferrari, P.F. Handling newborn monkeys alters later exploratory, cognitive, and social behaviors. Dev. Cogn. Neurosci. 2019, 35, 12–19. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Keebaugh, A.C.; Reamer, L.A.; Schaeffer, J.; Schapiro, S.J.; Young, L.J. Genetic influences on receptive joint attention in chimpanzees (Pan troglodytes). Sci. Rep. 2014, 4, 3774. [Google Scholar] [CrossRef]
- Bloomstrand, M.; Riddle, K.; Alford, P.; Maple, T.L. Objective evaluation of a behavioral enrichment device for captive chimpanzees (Pan troglodytes). Zoo Biol. 1986, 5, 293–300. [Google Scholar] [CrossRef]
- Csatádi, K.; Leus, K.; Pereboom, J.J. A brief note on the effects of novel enrichment on an unwanted behaviour of captive bonobos. Appl. Anim. Behav. Sci. 2008, 112, 201–204. [Google Scholar] [CrossRef]
- Gilloux, I.; Gurnell, J.; Shepherdson, D. An Enrichment Device for Great Apes. Anim. Welf 1992, 1, 279–289. [Google Scholar]
- Nakamichi, M. Tool-use and tool-making by captive, group-living orangutans (Pongo pygmaeus abelii) at an artificial termite mound. Behav. Processes 2004, 65, 87–93. [Google Scholar] [CrossRef]
- Birkett, L.P.; Newton-Fisher, N.E. How abnormal is the behaviour of captive, zoo-living chimpanzees? PLoS ONE 2011, 6, e20101. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, D.H.; Ghirardo, S.; Minier, D.E.; Sharpe, N.; Tatum, L.; McCowan, B. Efficacy of 3 types of foraging enrichment for rhesus macaques (Macaca mulatta). J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 888–894. [Google Scholar]
- Rooney, M.B.; Sleeman, J. Effects of selected behavioral enrichment devices on behavior of Western Lowland Gorillas (Gorilla gorilla gorilla). J. Appl. Anim. Welf. Sci. 1998, 1, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.R.; Mendes, F.D.C. Induced Tool Use as Environmental Enrichment for Captive Capuchin Monkeys (Sapajus libidinosus)/Inducao do Uso de Ferramentas como Enriquecimento Ambiental para Macacos-prego (Sapajus libidinosus) Cativos. Psicologia Teoria Pesquisa 2016, 32, 1–8. [Google Scholar]
- Pfaller-Sadovsky, N.; Arnott, G.; Hurtado-Parrado, C. Using Principles from Applied Behaviour Analysis to Address an Undesired Behaviour: Functional Analysis and Treatment of Jumping Up in Companion Dogs. Animals 2019, 9, 1091. [Google Scholar] [CrossRef] [Green Version]
- Hanley, G.P.; Iwata, B.A.; McCord, B.E. Functional analysis of problem behavior: A review. J. Appl. Behav. Anal. 2003, 36, 147–185. [Google Scholar] [CrossRef] [Green Version]
- Hanley, G.P. Functional assessment of problem behavior: Dispelling myths, overcoming implementation obstacles, and developing new lore. Behav. Anal. Pract. 2012, 5, 54–72. [Google Scholar] [CrossRef] [Green Version]
- Lopresti-Goodman, S.M.; Kameka, M.; Dube, A. Stereotypical behaviors in chimpanzees rescued from the african bushmeat and pet trade. Behav. Sci. 2013, 3, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Bloomsmith, M.A.; Marr, M.J.; Maple, T.L. Addressing nonhuman primate behavioral problems through the application of operant conditioning: Is the human treatment approach a useful model? Appl. Anim. Behav. Sci. 2007, 102, 205–222. [Google Scholar] [CrossRef]
- Coleman, K.; Maier, A. The use of positive reinforcement training to reduce stereotypic behavior in rhesus macaques. Appl. Anim. Behav. Sci. 2010, 124, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, G.A.; Capaldo, T.; Lindner, L.; Grow, G. Building an inner sanctuary: Complex PTSD in chimpanzees. J. Trauma Dissociation 2008, 9, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Kummrow, M. Diagnostic and Therapeutic Guidelines to Abnormal Behavior in Captive Nonhuman Primates. Vet. Clin. N. Am. Exot. Anim. Pract. 2021, 24, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, S.R.; Vazquez, M.; Brasky, K. Combination therapy reduces self-injurious behavior in a chimpanzee (Pan Troglodytes Troglodytes): A case report. J. Appl. Anim. Welf. Sci. 2007, 10, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Robbertsen, F. Effect of Intervention on Abnormal Behaviour of Retired Laboratory Chimpanzees; Utrecht University: Utrectht, The Netherlands, 2014. [Google Scholar]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavioral Category | Definition |
|---|---|
| 1. Participation * | The Chimpanzee is Actively Interacting or in Contact with The Artificial Termite Mound. |
| 2. Tool use * | To use a Mobile Element, External to the Body, to Perform a Directed Action. Includes Tool Modification and Transportation. |
| 3. Feeding * | Searching, Locating, Handling, and Ingesting or Transporting Food. Includes Fluid Intake. |
| 4. Inactivity | No Action or Activity, Sitting or Lying Down. Includes Self-Observation, Yawning, and Sleeping. |
| 5. Abnormal Behaviors | Maladjusted Stereotypical Behaviors such as Rocking, Pacing, Self–Harm, Coprophagy (Eating Feces), Regurgitation, and Reingestion, Trichotillomania (Hair-Pulling), Trichotillophagia (Hair-Pulling Eating Hair), Ear-Poking, Eye-Poking. |
| 6. Self-Directed Behaviors | Instantaneous Scan Sampling (All Phases): Behaviors Directed towards the Subject, such as Body Inspection, Self-Grooming, Masturbation and Scratching or Rubbing One’s Body with the Hands or Fingers. Untimed-Event Focal Sampling (Treatment Phase): Includes the Following Behaviors: (1) Scratches: Continuous Movement of the Hand over the Skin Involving the Ends of the Digits or Nails; (2) Rubs: Continuous Movement of the Hand over The Skin not Involving the Ends of the Digits Performed Either with the Palmar, Dorsal or Lateral Side of the Hand. This Category also Includes Self-Touches such as Nose-Wiping [48] or Face Stroking [46]. |
| 7. Social Proximity * | The Chimpanzee is at Less Than One-Arm Length from One or More Subjects while Performing any Solitary Activity, but with no Social Interaction between Subjects. |
| 8. Affiliation-related Behaviors | Includes the Following Behaviors: (1) Grooming: Body-Cleansing Behavior (Grooming) from one Individual to Another (includes Mutual Grooming), Performed with the Upper Extremities or with the Mouth; (2) Social Play: Playful Behavior between Two or More Individuals Associated with Behavioral Indicators of Play (e.g., Play-Face, Laugh, Friendly Head Bobbing, Soft Knocking on the Ground, and Playful Chasing); (3) Sexual Behavior: Sexual Interaction, or search for Sexual Interaction, between Two Individuals Including Behaviors such as Copulation, Attempted Copulation, Genital Presentation, and Other Behaviors Directed Towards the Genitals of Another Individual; (4) Other Behaviors Identified as Affiliative, but do not fit the Criteria of Grooming, Social Play or Sexual Activity (Embrace, Feed Together and Follow). |
| 9. Aggression-related Behaviors | Includes the Following Behaviors: (1) Agonistic Dominance: Threat-Related Behaviors such as Direct Aggression, Charging Display, Displacement and Resource Appropriation (e.g., Steal Food or Objects). Can be Accompanied by Vocalizations; (2) Agonistic Submission: Avoiding, Bared-Teeth, Displays, Food Submission (e.g., Leave/Drop Food and Move Away when Others try to Steal It), Hand-To-Mouth, Finger-To-Mouth. Can be Accompanied by Vocalizations such as Pant-Grunts. Includes Running Away from Others in Conflict Situations; (3) Other Behaviors Identified as Agonistic, but do not Fit the Criteria of Agonistic Dominance or Agonistic Submission. (Appeasing, Consolation, Reconciliation and Requesting Support). |
| Not Visible/Not Present | The Chimpanzee or the Behavior cannot be Identified, or the Chimpanzee is not in the Outdoor Enclosure (e.g., he is in the Sleeping Areas or in the Outdoor Cages). |
| Models & Predictions | Supported? | ||||
|---|---|---|---|---|---|
| 1. Participation in E… | Decreases Across Sessions is Predicted by: Sex Age Time (am/pm) | No * No No Yes | |||
| E Presence/Participation Predicts a/an… | E Presence (Models 2–9) | Participation (Models 2b–9b) | |||
| Short–Term | Long–Term | ||||
| Baseline ≠ E | Pre ≠ E | Pre ≠ Post | |||
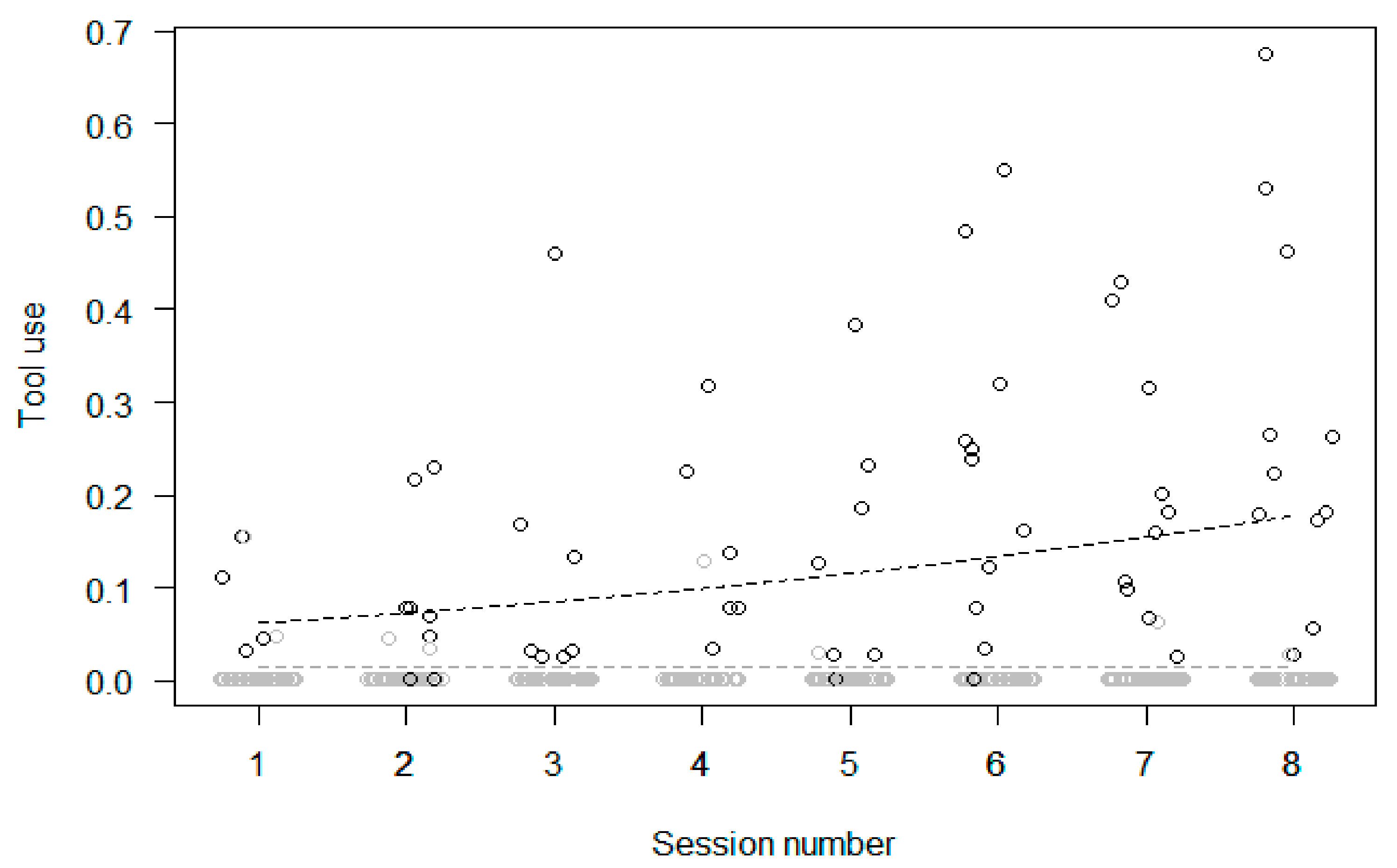
| 2/2b Increase in Tool Use | Yes | Yes | Yes | Yes | |
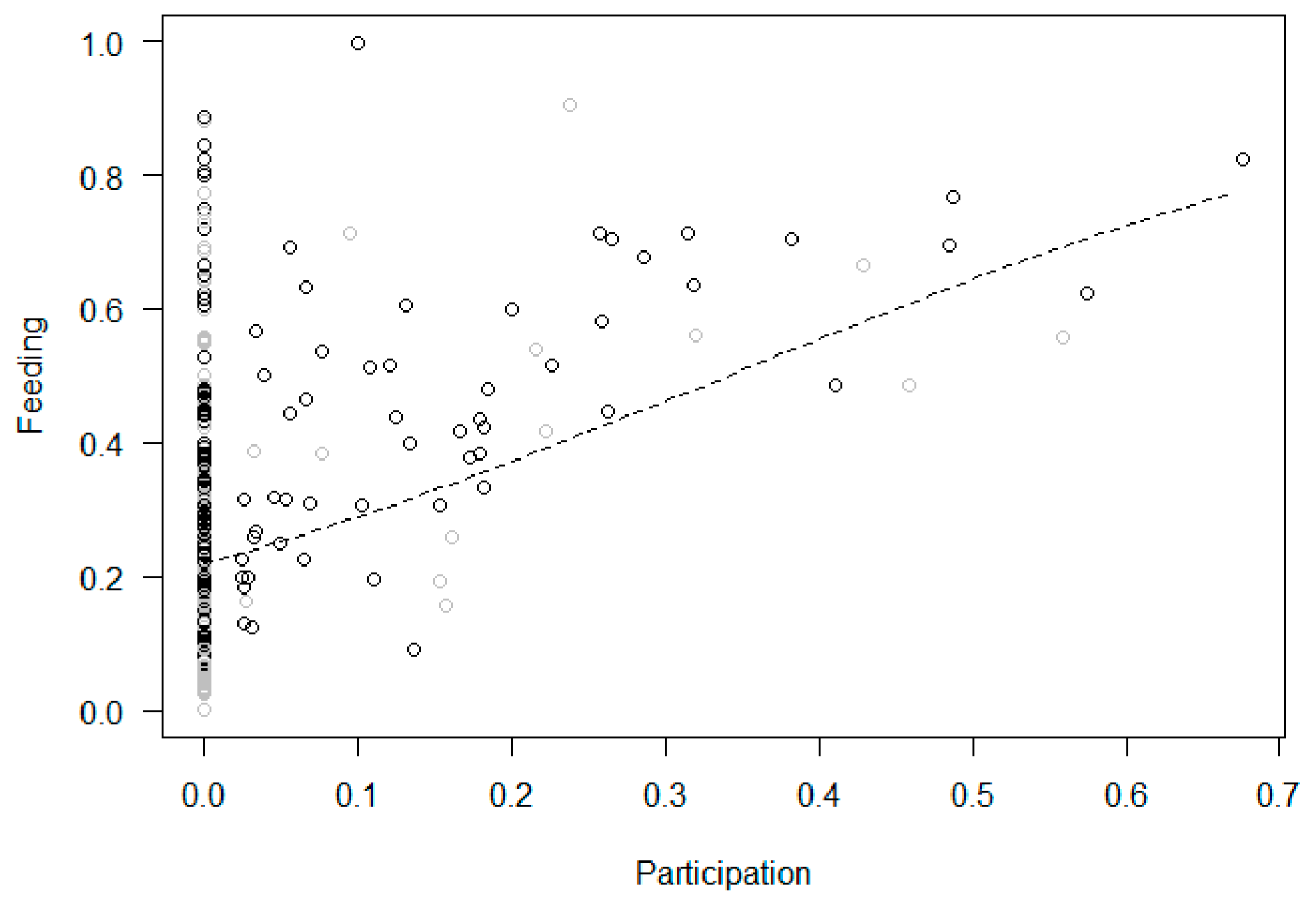
| 3/3b Increase in Feeding | Yes | Yes | Yes | Yes | |
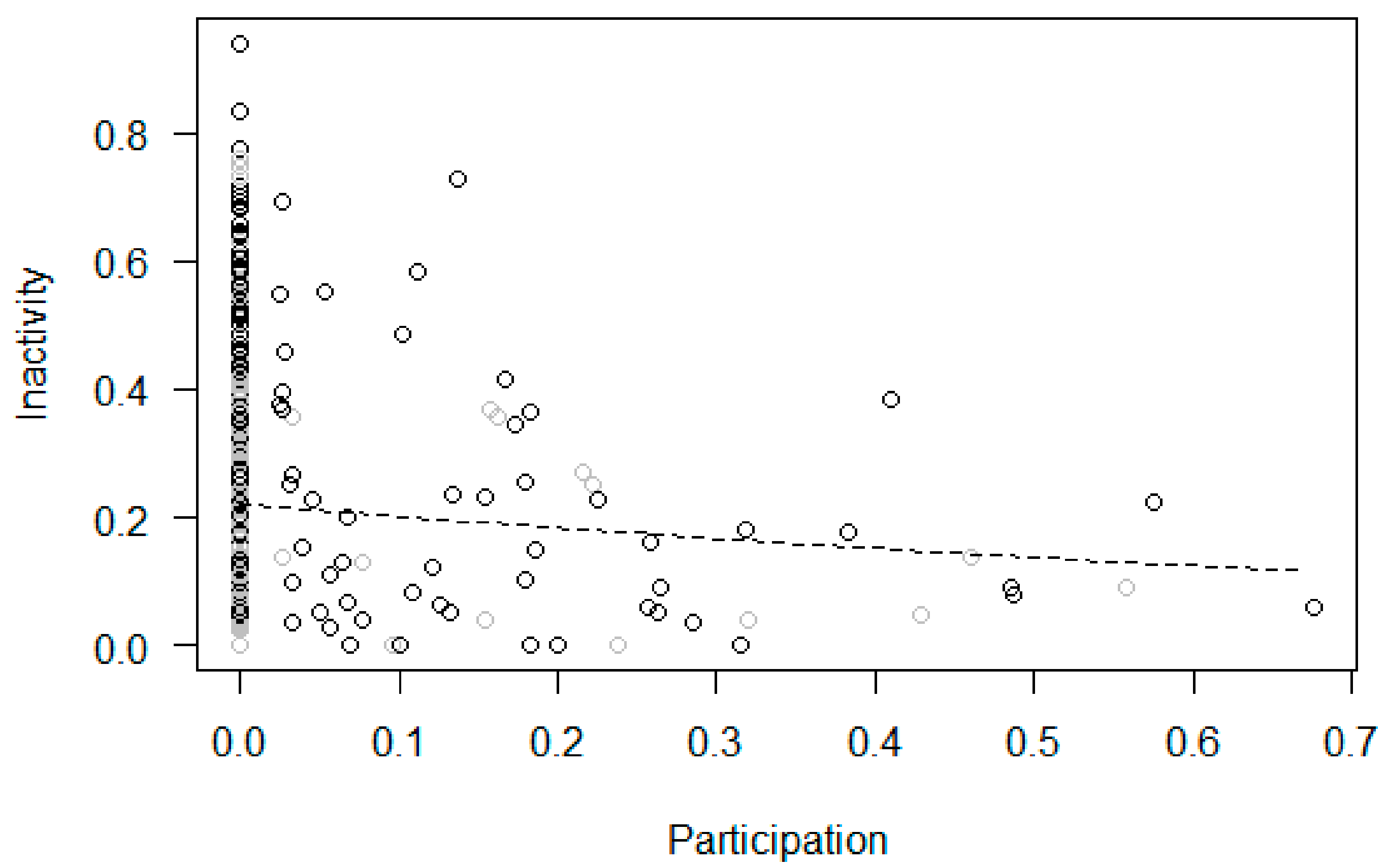
| 4/4b Decrease in Inactivity | Yes | Yes | Yes | Yes | |
| 5/5b Decrease in Abnormal Behaviors | Yes | No | No ** | No | |
| 6 Decrease in Self-Directed Behaviors | Yes | No ** | No ** | – | |
| 6b Decrease in Rubs and Scratches | – | – | – | No | |
| 7/7b Increase in Social Proximity | No | Yes | Yes | Yes | |
| 8/8b Decrease in Affiliation-related Behaviors | No | No ** | No ** | No | |
| 9/9b Increase in Aggression-related Behaviors | No | Yes | No | No | |
| 10. E Presence Predicts Changes in Proximity Social Networks | No | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padrell, M.; Amici, F.; Córdoba, M.P.; Giberga, A.; Broekman, A.; Almagro, S.; Llorente, M. Artificial Termite-Fishing Tasks as Enrichment for Sanctuary-Housed Chimpanzees: Behavioral Effects and Impact on Welfare. Animals 2021, 11, 2941. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11102941
Padrell M, Amici F, Córdoba MP, Giberga A, Broekman A, Almagro S, Llorente M. Artificial Termite-Fishing Tasks as Enrichment for Sanctuary-Housed Chimpanzees: Behavioral Effects and Impact on Welfare. Animals. 2021; 11(10):2941. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11102941
Chicago/Turabian StylePadrell, Maria, Federica Amici, Maria Pau Córdoba, Albert Giberga, Antonio Broekman, Susana Almagro, and Miquel Llorente. 2021. "Artificial Termite-Fishing Tasks as Enrichment for Sanctuary-Housed Chimpanzees: Behavioral Effects and Impact on Welfare" Animals 11, no. 10: 2941. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11102941