Effect of Heat Stress on Bovine Mammary Cellular Metabolites and Gene Transcription Related to Amino Acid Metabolism, Amino Acid Transportation and Mammalian Target of Rapamycin (mTOR) Signaling

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. RNA Extraction and Real-Time PCR Analysis
2.3. LC-MS Analysis
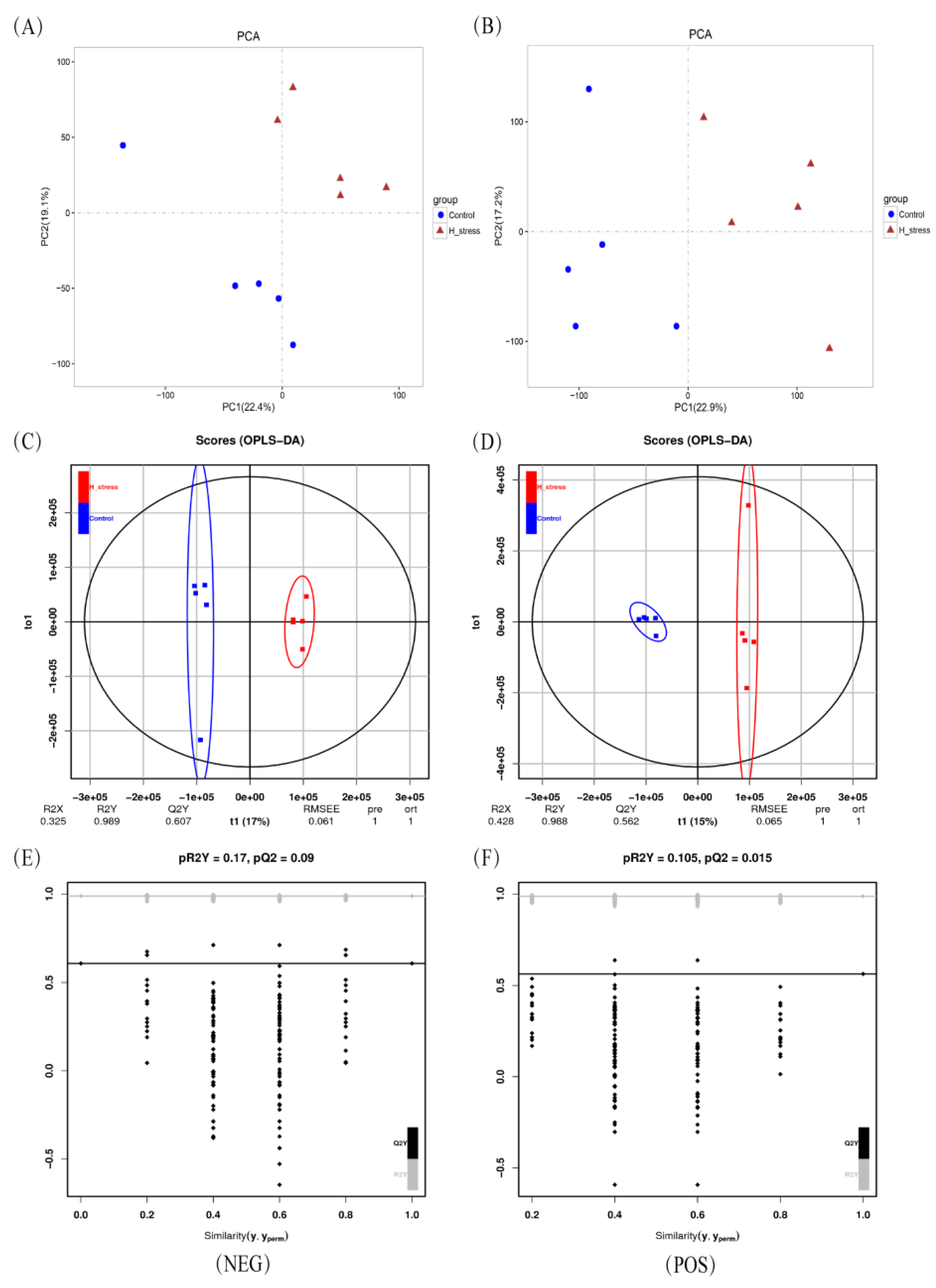
2.4. Data Analysis
3. Results
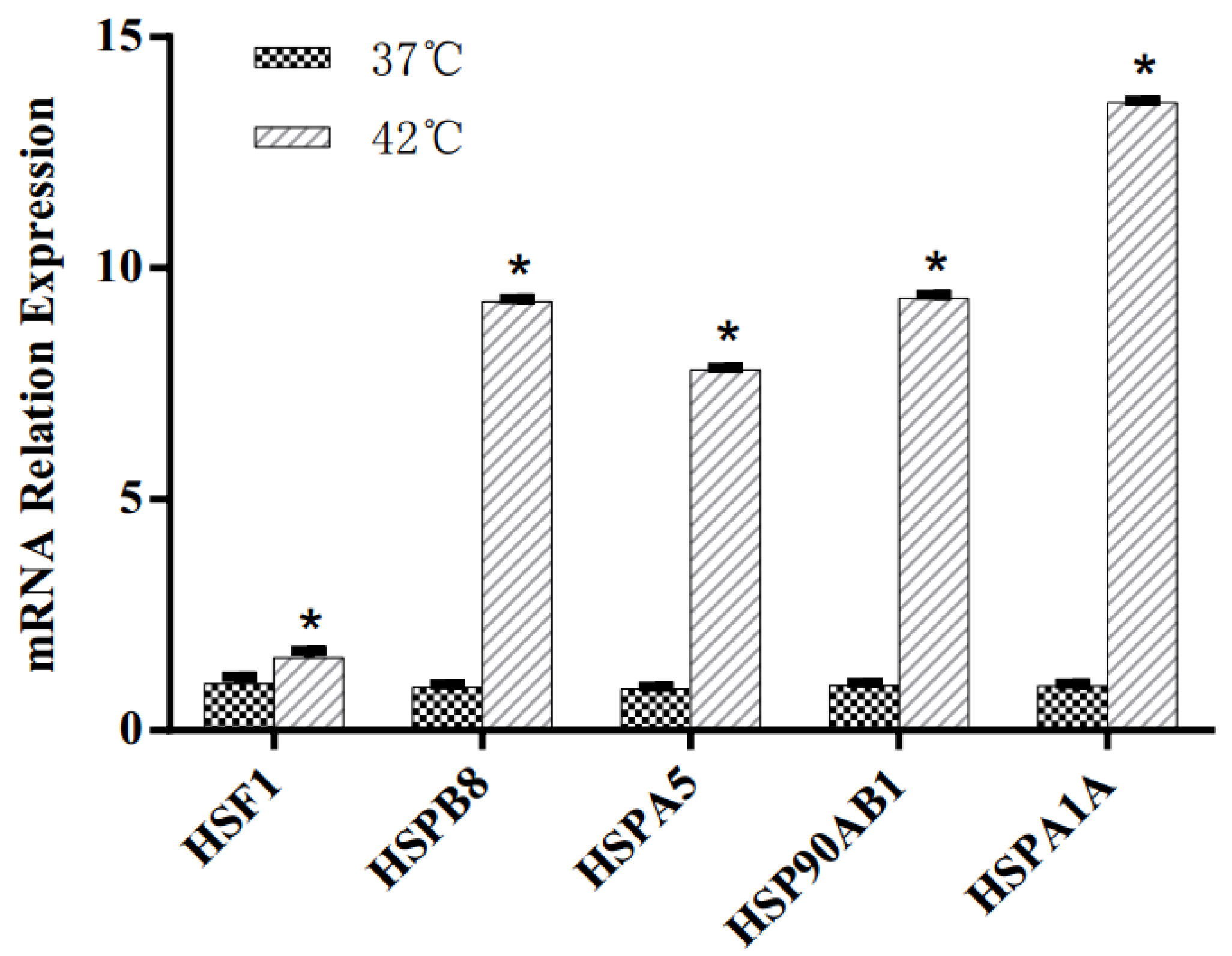
3.1. Heat Shock Response of MAC-T Cells
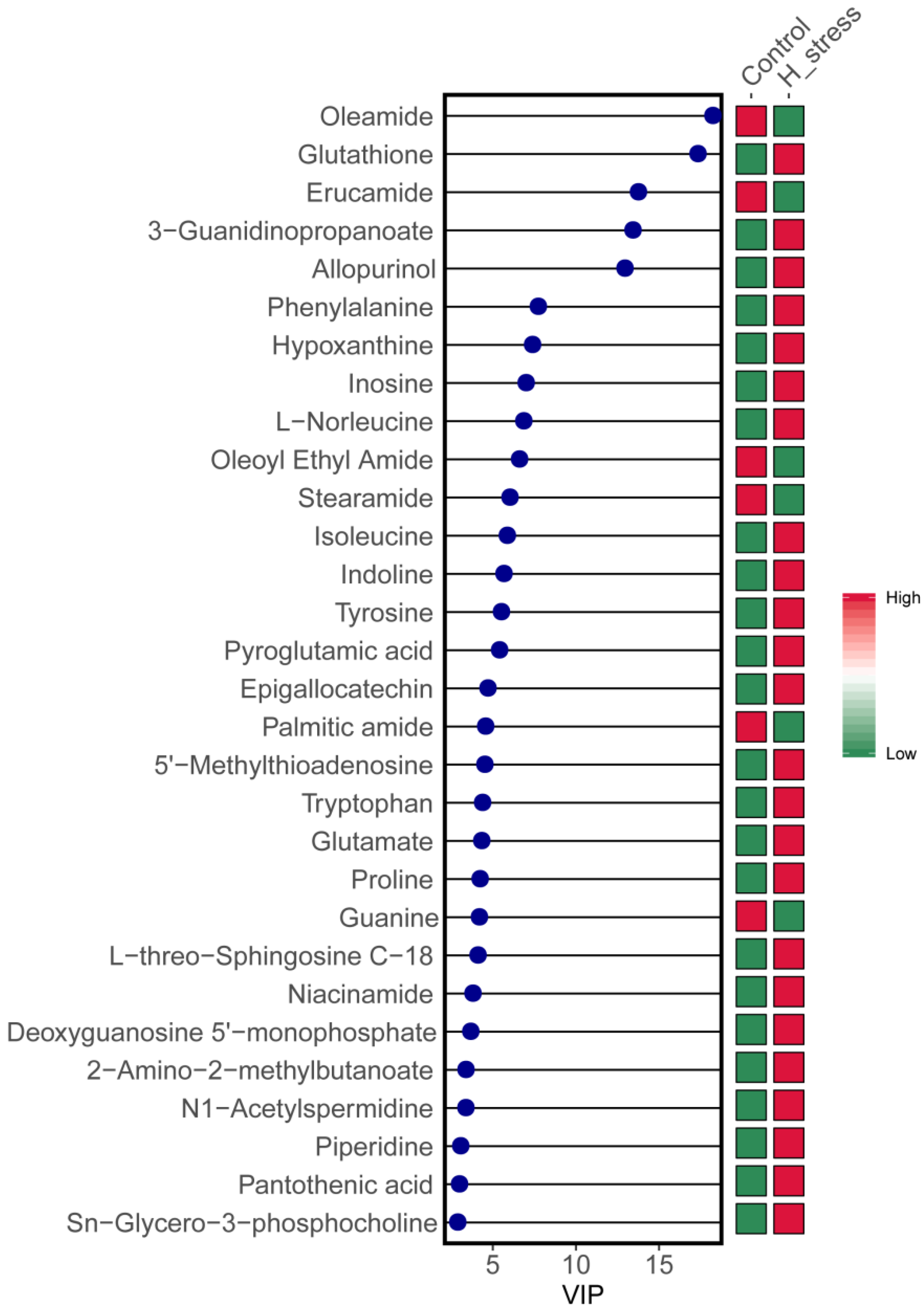
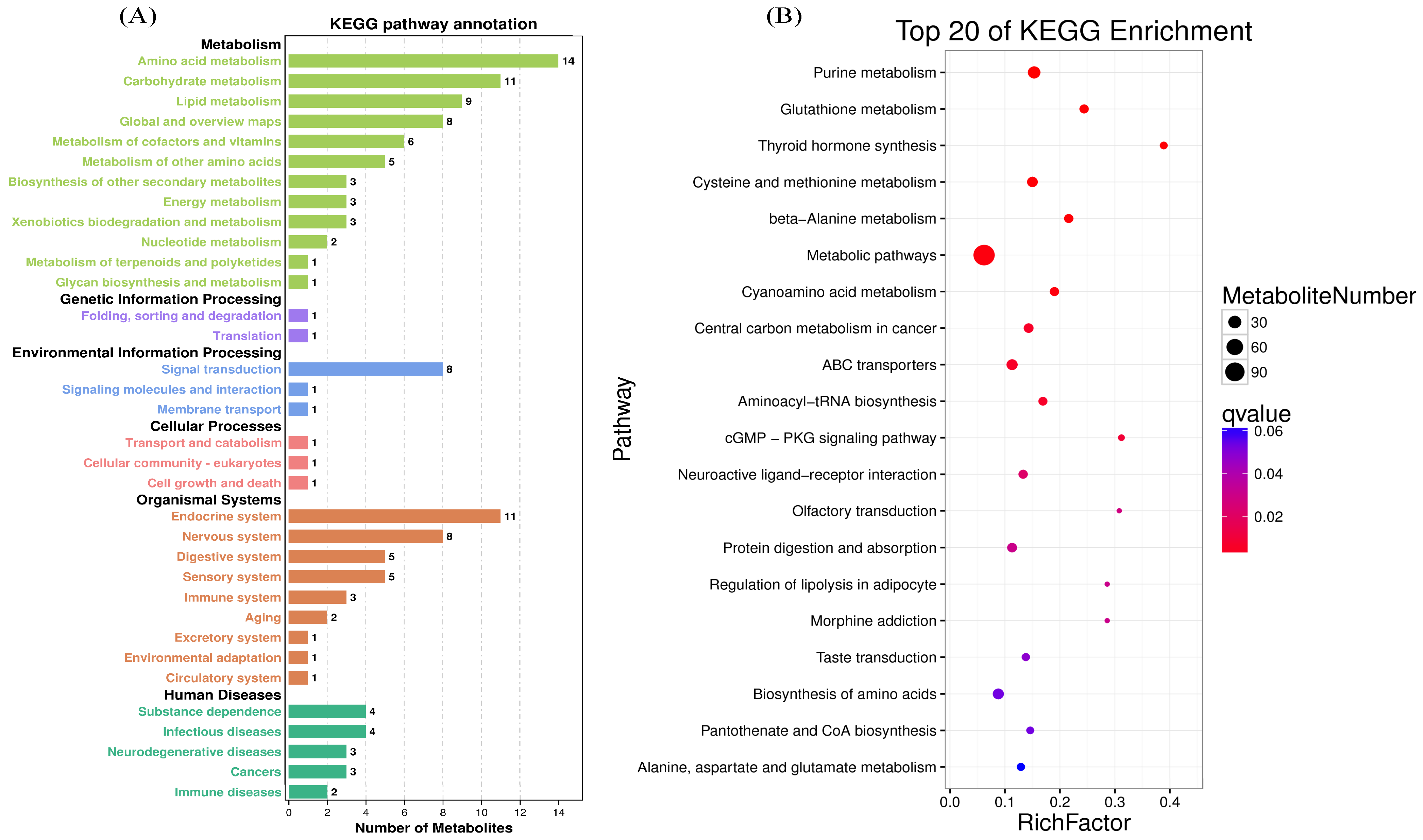
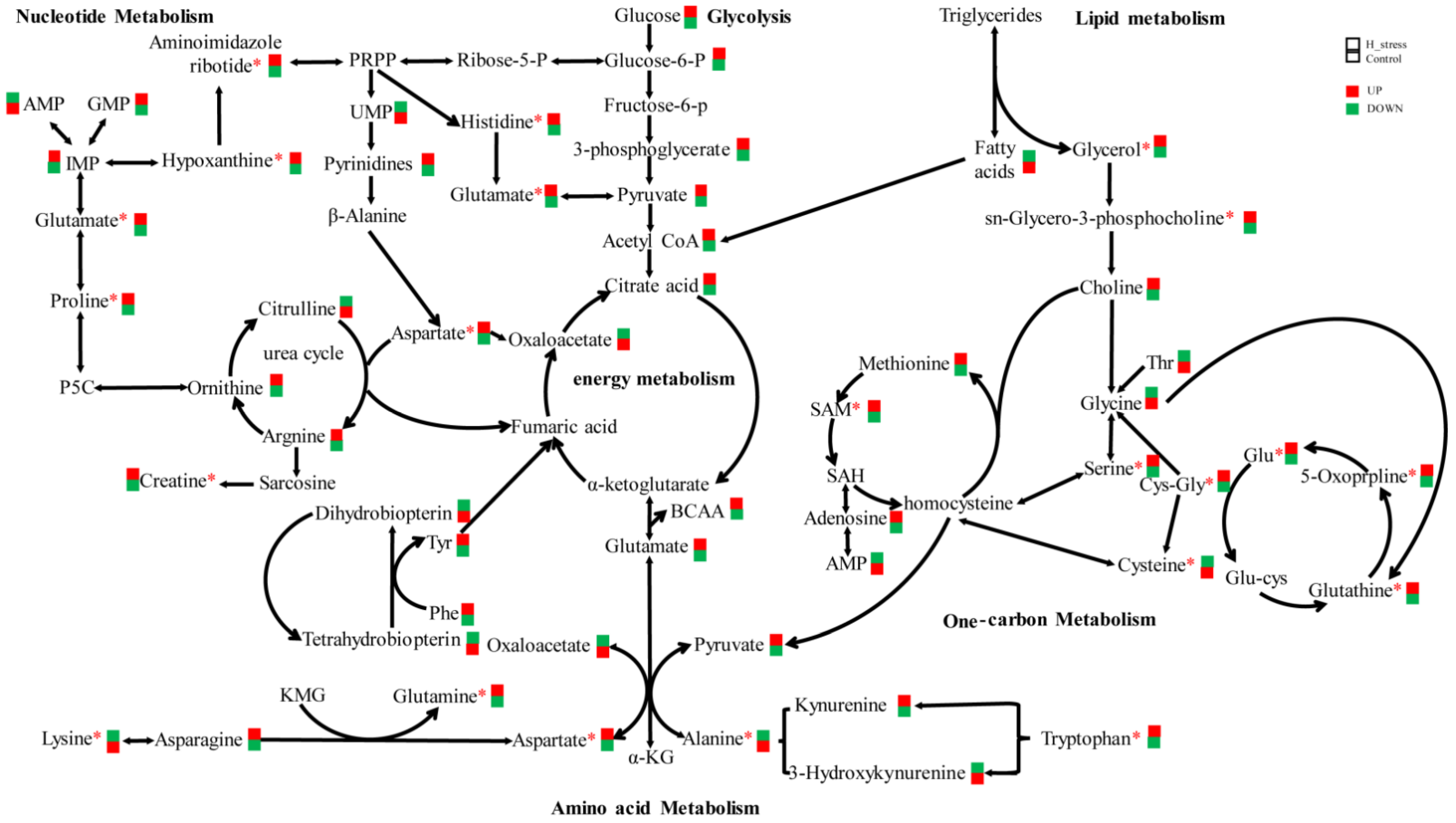
3.2. Intracellular Metabolism of MAC-T Cells
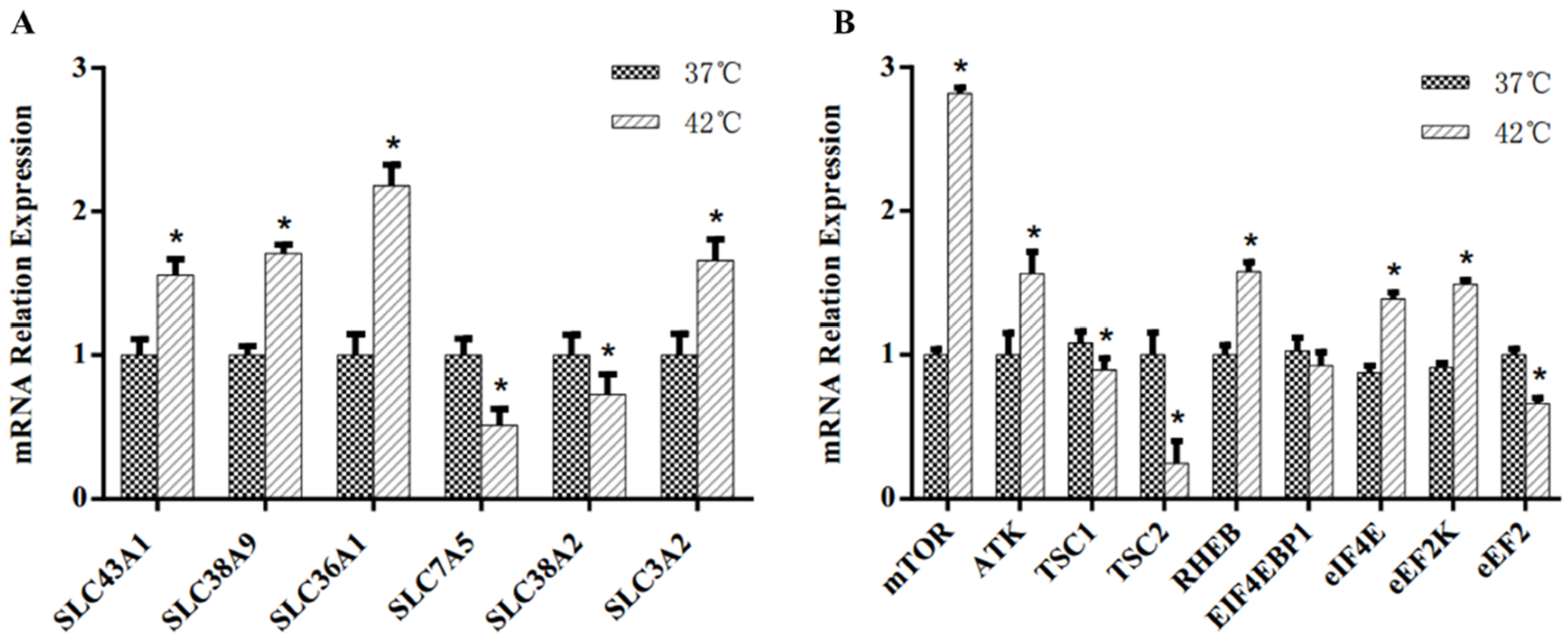
3.3. Milk Protein Synthesis Regulation of MAC-T Cells
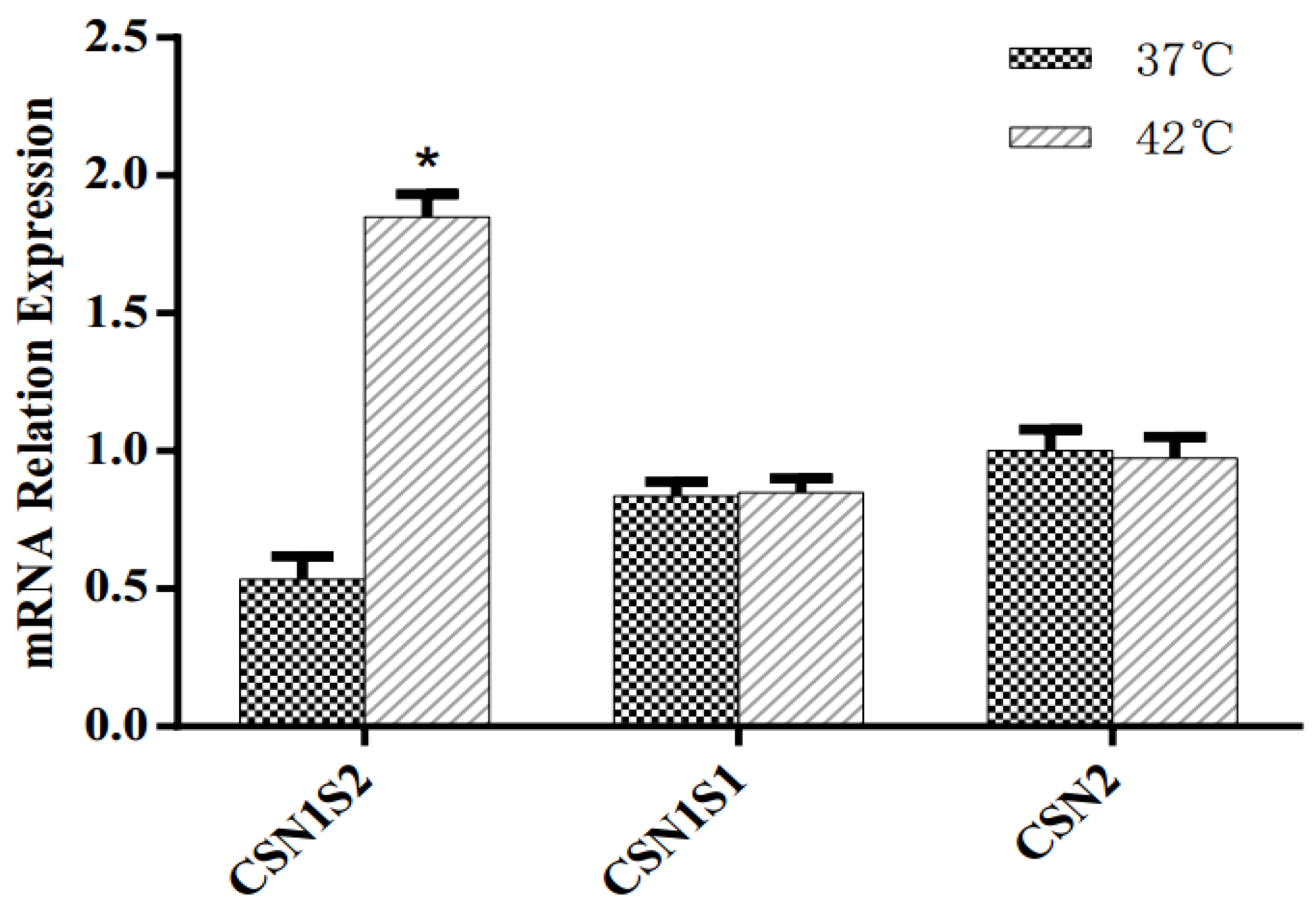
3.4. Casein Synthesis of MAC-T Cells
4. Discussion
4.1. Heat Stress Model Construction
4.2. Metabolomics
4.2.1. Glutathione Metabolism
4.2.2. Phenylalanine, Tyrosine and Tryptophan Metabolism
4.2.3. Valine, Leucine and Isoleucine Metabolism
4.2.4. Arginine and Proline Metabolism
4.2.5. Alanine, Aspartate and Glutamate Metabolism
4.2.6. Cysteine and Methionine Metabolism
4.3. Amino Acid Transporters
4.4. mTOR Signaling Pathway
4.5. Milk Protein Synthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tao, S.; Orellana, R.M.; Weng, X.; Marins, T.N.; Dahl, G.E.; Bernard, J.K. Symposium Review: The Influences of Heat Stress on Bovine Mammary Gland Function. J. Dairy Sci. 2018, 101, 5642–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlgemuth, S.E.; Ramirez-Lee, Y.; Tao, S.; Monteiro, A.P.A.; Ahmed, B.M.; Dahl, G.E. Short Communication: Effect of Heat Stress on Markers of Autophagy in the Mammary Gland during the Dry Period. J. Dairy Sci. 2016, 99, 4875–4880. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Orellana Rivas, R.M.; Marins, T.N.; Chen, Y.C.; Gao, J.; Bernard, J.K. Impact of Heat Stress on Lactational Performance of Dairy Cows. Theriogenology 2020, 150, 437–444. [Google Scholar] [CrossRef]
- Ogunbode, C.A.; Doran, R.; Böhm, G. Exposure to the IPCC Special Report on 1.5 °C Global Warming Is Linked to Perceived Threat and Increased Concern about Climate Change. Clim. Chang. 2020, 158, 361–375. [Google Scholar] [CrossRef]
- Gao, S.T.; Ma, L.; Zhou, Z.; Zhou, Z.K.; Baumgard, L.H.; Jiang, D.; Bionaz, M.; Bu, D.P. Heat Stress Negatively Affects the Transcriptome Related to Overall Metabolism and Milk Protein Synthesis in Mammary Tissue of Midlactating Dairy Cows. Physiol. Genom. 2019, 51, 400–409. [Google Scholar] [CrossRef]
- Yue, S.; Wang, Z.; Wang, L.; Peng, Q.; Xue, B. Transcriptome Functional Analysis of Mammary Gland of Cows in Heat Stress and Thermoneutral Condition. Animals 2020, 10, 1015. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, J.; Gao, H.; Li, S.; Zhang, Y.; Zheng, N. Heat-Induced Apoptosis and Gene Expression in Bovine Mammary Epithelial Cells. Anim. Prod. Sci. 2016, 56, 918. [Google Scholar] [CrossRef]
- Xiao, Y.; Rungruang, S.; Hall, L.W.; Collier, J.L.; Dunshea, F.R.; Collier, R.J. Effects of Niacin and Betaine on Bovine Mammary and Uterine Cells Exposed to Thermal Shock in Vitro. J. Dairy Sci. 2017, 100, 4025–4037. [Google Scholar] [CrossRef]
- Weng, X.; Monteiro, A.P.A.; Guo, J.; Li, C.; Orellana, R.M.; Marins, T.N.; Bernard, J.K.; Tomlinson, D.J.; DeFrain, J.M.; Wohlgemuth, S.E.; et al. Effects of Heat Stress and Dietary Zinc Source on Performance and Mammary Epithelial Integrity of Lactating Dairy Cows. J. Dairy Sci. 2018, 101, 2617–2630. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Sun, Y.; Wu, J.; Li, X.; Luo, M.; Wang, G. The Global Effect of Heat on Gene Expression in Cultured Bovine Mammary Epithelial Cells. Cell Stress Chaperones 2015, 20, 381–389. [Google Scholar] [CrossRef] [Green Version]
- Loor, J.J.; Bionaz, M.; Drackley, J.K. Systems Physiology in Dairy Cattle: Nutritional Genomics and Beyond. Annu. Rev. Anim. Biosci. 2013, 1, 365–392. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, M. Metabolomic Profiles in Yak Mammary Gland Tissue during the Lactation Cycle. PLoS ONE 2019, 14, e0219220. [Google Scholar] [CrossRef]
- Tong, J.; Zhang, H.; Zhang, Y.; Xiong, B.; Jiang, L. Microbiome and Metabolome Analyses of Milk from Dairy Cows with Subclinical Streptococcus Agalactiae Mastitis—Potential Biomarkers. Front. Microbiol. 2019, 10, 2547. [Google Scholar] [CrossRef]
- Yang, Y.; Zheng, N.; Zhao, X.; Zhang, Y.; Han, R.; Yang, J.; Zhao, S.; Li, S.; Guo, T.; Zang, C.; et al. Metabolomic Biomarkers Identify Differences in Milk Produced by Holstein Cows and Other Minor Dairy Animals. J. Proteom. 2016, 136, 174–182. [Google Scholar] [CrossRef]
- Sun, H.Z.; Shi, K.; Wu, X.H.; Xue, M.Y.; Wei, Z.H.; Liu, J.X.; Liu, H.Y. Lactation-Related Metabolic Mechanism Investigated Based on Mammary Gland Metabolomics and 4 Biofluids’ Metabolomics Relationships in Dairy Cows. BMC Genom. 2017, 18, 936. [Google Scholar] [CrossRef] [Green Version]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC–MS Metabolomics Identifies Metabolite Alterations That Precede Subclinical Mastitis in the Blood of Transition Dairy Cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef]
- Dong, X.; Zhou, Z.; Wang, L.; Saremi, B.; Helmbrecht, A.; Wang, Z.; Loor, J.J. Increasing the Availability of Threonine, Isoleucine, Valine, and Leucine Relative to Lysine While Maintaining an Ideal Ratio of Lysine:Methionine Alters Mammary Cellular Metabolites, Mammalian Target of Rapamycin Signaling, and Gene Transcription. J. Dairy Sci. 2018, 101, 5502–5514. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Zhou, Z.; Saremi, B.; Helmbrecht, A.; Wang, Z.; Loor, J.J. Varying the Ratio of Lys:Met While Maintaining the Ratios of Thr:Phe, Lys:Thr, Lys:His, and Lys:Val Alters Mammary Cellular Metabolites, Mammalian Target of Rapamycin Signaling, and Gene Transcription. J. Dairy Sci. 2018, 101, 1708–1718. [Google Scholar] [CrossRef]
- Salama, A.A.K.; Duque, M.; Wang, L.; Shahzad, K.; Olivera, M.; Loor, J.J. Enhanced Supply of Methionine or Arginine Alters Mechanistic Target of Rapamycin Signaling Proteins, Messenger RNA, and MicroRNA Abundance in Heat-Stressed Bovine Mammary Epithelial Cells in Vitro. J. Dairy Sci. 2019, 102, 2469–2480. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Wang, Y.; Li, L.; Han, Z.; Mao, S.; Wang, G. Betaine protects against heat exposure-induced oxidative stress and apoptosis in bovine mammary epithelial cells via regulation of ROS production. Cell Stress Chaperones 2019, 24, 453–460. [Google Scholar] [CrossRef]
- Yang, M.; Kuang, M.; Wang, G.; Ali, I.; Tang, Y.; Yang, C.; Li, Y.; Li, L. Choline attenuates heat stress-induced oxidative injury and apoptosis in bovine mammary epithelial cells by modulating PERK/Nrf-2 signaling pathway. Mol. Immunol. 2021, 135, 388–397. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene Networks Driving Bovine Mammary Protein Synthesis during the Lactation Cycle. Bioinform. Biol. Insights 2011, 5, BBI.S7003. [Google Scholar] [CrossRef]
- Livak, K.; JcSchmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ding, J.; Huang, Y.; Liu, S.; Zhang, S.; Zou, H.; Wang, Z.; Zhu, W.; Geng, J. Toxicological effects of nano- and micro-polystyrene plastics on red tilapia: Are larger plastic particles more harmless? J. Hazard. Mater. 2020, 396, 122693. [Google Scholar] [CrossRef]
- Jin, Q.; Jiao, C.; Sun, S.; Song, C.; Cai, Y.; Lin, Y.; Fan, H.; Zhu, Y. Metabolic Analysis of Medicinal Dendrobium officinale and Dendrobium huoshanense during Different Growth Years. PLoS ONE 2016, 11, e0146607. [Google Scholar] [CrossRef]
- Saccenti, E.; Hoefsloot, H.C.J.; Smilde, A.K. Reflections on univariate and multivariate analysis of metabolomics data. Metabolomics 2014, 10, 361–374. [Google Scholar] [CrossRef]
- Wu, G. Amino Acids: Biochemistry and Nutrition; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Li, A.; Guo, X.; Xie, J.; Liu, X.; Zhang, Z.; Li, Y.; Zhang, Y. Validation of biomarkers in cardiotoxicity induced by Periplocin on neonatal rat cardiomyocytes using UPLC-Q-TOF/MS combined with a support vector machine. J. Pharm Biomed. Anal. 2016, 123, 179–185. [Google Scholar] [CrossRef]
- Avila, M.A.; García-Trevijano, E.R.; Lu, S.C.; Corrales, F.J.; Mato, J.M. Methylthioadenosine. Int. J. Biochem. Cell Biol. 2004, 36, 2125–2130. [Google Scholar] [CrossRef]
- Collier, M.P.; Benesch, J.L.P. Small Heat-Shock Proteins and Their Role in Mechanical Stress. Cell Stress Chaperones 2020, 25, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Nowakowska, M.; Gualtieri, F.; von Rüden, E.-L.; Hansmann, F.; Baumgärtner, W.; Tipold, A.; Potschka, H. Profiling the Expression of Endoplasmic Reticulum Stress Associated Heat Shock Proteins in Animal Epilepsy Models. Neuroscience 2020, 429, 156–172. [Google Scholar] [CrossRef]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated Metabolomics Study of the Milk of Heat-Stressed Lactating Dairy Cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef]
- Min, L.; Zhao, S.; Tian, H.; Zhou, X.; Zhang, Y.; Li, S.; Yang, H.; Zheng, N.; Wang, J. Metabolic Responses and “Omics” Technologies for Elucidating the Effects of Heat Stress in Dairy Cows. Int. J. Biometeorol. 2017, 61, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, B.; Zhang, L.; Li, J.; Xing, T.; Jiang, Y.; Gao, F. Heat Stress Alters Muscle Protein and Amino Acid Metabolism and Accelerates Liver Gluconeogenesis for Energy Supply in Broilers. Poult. Sci. 2021, 100, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Gao, S.; Quan, S.; Zhang, Y.; Bu, D.; Wang, J. Blood Amino Acids Profile Responding to Heat Stress in Dairy Cows. Asian-Australas J. Anim. Sci. 2018, 31, 47–53. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; He, X.; Lu, Z.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Chronic Heat Stress Affects Muscle Hypertrophy, Muscle Protein Synthesis and Uptake of Amino Acid in Broilers via Insulin like Growth Factor-Mammalian Target of Rapamycin Signal Pathway. Poult. Sci. 2018, 97, 4150–4158. [Google Scholar] [CrossRef] [PubMed]
- Anchordoquy, J.P.; Lizarraga, R.M.; Anchordoquy, J.M.; Nikoloff, N.; Rosa, D.E.; Fabra, M.C.; Peral-García, P.; Furnus, C.C. Effect of Cysteine, Glutamate and Glycine Supplementation to in Vitro Fertilization Medium during Bovine Early Embryo Development. Reprod. Biol. 2019, 19, 349–355. [Google Scholar] [CrossRef]
- Nabenishi, H.; Ohta, H.; Nishimoto, T.; Morita, T.; Ashizawa, K.; Tsuzuki, Y. The effects of cysteine addition during in vitro maturation on the developmental competence, ROS, GSH and apoptosis level of bovine oocytes exposed to heat stress. Zygote 2012, 20, 249–259. [Google Scholar] [CrossRef]
- Pocius, P.A.; Clark, J.H.; Baumrucker, C.R. Glutathione in bovine blood: Possible source of amino acids for milk protein synthesis. J. Dairy Sci. 1981, 64, 1551–1554. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.E.; Lee, J.S.; Park, J.S.; Moon, J.O.; Lee, H.G. Phenylalanine and valine differentially stimulate milk protein synthetic and energy-mediated pathway in immortalized bovine mammary epithelial cells. J. Anim. Sci. Technol. 2020, 62, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Doelman, J.; Curtis, R.V.; Carson, M.; Kim, J.J.M.; Metcalf, J.A.; Cant, J.P. Essential Amino Acid Infusions Stimulate Mammary Expression of Eukaryotic Initiation Factor 2Bε but Milk Protein Yield Is Not Increased during an Imbalance. J. Dairy Sci. 2015, 98, 4499–4508. [Google Scholar] [CrossRef]
- Lieberman, H.R.; Georgelis, J.H.; Maher, T.J.; Yeghiayan, S.K. Tyrosine prevents effects of hyperthermia on behavior and increases norepinephrine. Physiol. Behav. 2005, 84, 3338. [Google Scholar] [CrossRef]
- Lee, S.B.; Lee, K.W.; Wang, T.; Lee, J.S.; Jung, U.S.; Nejad, J.G.; Oh, Y.K.; Baek, Y.C.; Kim, K.H.; Lee, H.G. Intravenous administration of L–tryptophan stimulates gastrointestinal hormones and melatonin secretions: Study on beef cattle. J. Anim. Sci. Technol. 2019, 61, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.T.; Nejad, J.G.; Moon, J.O.; Lee, H.G. Dietary supplementation of acetate-conjugated tryptophan alters feed intake, milk yield and composition, blood profile, physiological variables, and heat shock protein gene expression in heat-stressed dairy cows. J. Therm. Biol. 2021, 98, 102949. [Google Scholar] [CrossRef] [PubMed]
- Indikatiya Hewage, I. The Effect of a Limited Supply of Phenylalanine, Threonine, or Tryptophan on Mammary Metabolism in Dairy Cows. Master’s Thesis, University of Calgary, Calgary, AB, Canada, 2015. [Google Scholar]
- Rezaei, R.; Wu, Z.; Hou, Y.; Bazer, F.W.; Wu, G. Amino Acids and Mammary Gland Development: Nutritional Implications for Milk Production and Neonatal Growth. J. Anim. Sci. Biotechnol. 2016, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Zeng, X.; Ren, M.; Mao, X.; Qiao, S. Novel Metabolic and Physiological Functions of Branched Chain Amino Acids: A Review. J. Anim. Sci. Biotechnol. 2017, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.I.; Kim, T.G.; Lim, D.H.; Kim, S.B.; Park, S.M.; Lim, H.J.; Kim, H.J.; Ki, K.S.; Kwon, E.G.; Kim, Y.J.; et al. The Effect of Nanoemulsified Methionine and Cysteine on the in Vitro Expression of Casein in Bovine Mammary Epithelial Cells. Asian-Australas J. Anim. Sci. 2019, 32, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manjarín, R.; Columbus, D.A.; Suryawan, A.; Nguyen, H.V.; Hernandez-García, A.D.; Hoang, N.-M.; Fiorotto, M.L.; Davis, T. Leucine Supplementation of a Chronically Restricted Protein and Energy Diet Enhances MTOR Pathway Activation but Not Muscle Protein Synthesis in Neonatal Pigs. Amino Acids 2016, 48, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Appuhamy, J.A.D.R.N.; Knoebel, N.A.; Nayananjalie, W.A.D.; Escobar, J.; Hanigan, M.D. Isoleucine and Leucine Independently Regulate MTOR Signaling and Protein Synthesis in MAC-T Cells and Bovine Mammary Tissue Slices. J. Nutr. 2012, 142, 484–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, S.M., Jr. Enzymes of Arginine Metabolism. J. Nutr. 2004, 134 (Suppl. 10), 2743S–2747S. [Google Scholar] [CrossRef] [PubMed]
- Manjarin, R.; Bequette, B.J.; Wu, G.; Trottier, N.L. Linking our understanding of mammary gland metabolism to amino acid nutrition. Amino Acids 2014, 46, 2447–2462. [Google Scholar] [CrossRef]
- O’Quinn, P.R.; Knabe, D.A.; Wu, G. Arginine Catabolism in Lactating Porcine Mammary Tissue. J. Anim. Sci. 2002, 80, 467–474. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Mancuso, A.; Daikhin, E.; Nissim, I.; Yudkoff, M.; Wehrli, S.; Thompson, C.B. Beyond Aerobic Glycolysis: Transformed Cells Can Engage in Glutamine Metabolism That Exceeds the Requirement for Protein and Nucleotide Synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 19345–19350. [Google Scholar] [CrossRef] [Green Version]
- Metallo, C.M.; Gameiro, P.A.; Bell, E.L.; Mattaini, K.R.; Yang, J.; Hiller, K.; Jewell, C.M.; Johnson, Z.R.; Irvine, D.J.; Guarente, L.; et al. Reductive Glutamine Metabolism by IDH1 Mediates Lipogenesis under Hypoxia. Nature 2012, 481, 380–384. [Google Scholar] [CrossRef] [Green Version]
- Meijer, A.J. Amino Acids as Regulators and Components of Nonproteinogenic Pathways. J. Nutr. 2003, 133, 2057S–2062S. [Google Scholar] [CrossRef]
- Bott, A.J.; Shen, J.; Tonelli, C.; Zhan, L.; Sivaram, N.; Jiang, Y.P.; Yu, X.; Bhatt, V.; Chiles, E.; Zhong, H.; et al. Glutamine Anabolism Plays a Critical Role in Pancreatic Cancer by Coupling Carbon and Nitrogen Metabolism. Cell Rep. 2019, 29, 1287–1298. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Nagao, M.; Toh, R.; Irino, Y.; Shinohara, M.; Iino, T.; Yoshikawa, S.; Tanaka, H.; Satomi-Kobayashi, S.; Ishida, T.; et al. Critical Role of Glutamine Metabolism in Cardiomyocytes under Oxidative Stress. Biochem. Biophys. Res. Commun. 2021, 534, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Bin, P.; Huang, R.; Zhou, X. Oxidation Resistance of the Sulfur Amino Acids: Methionine and Cysteine. BioMed Res. Int. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kalhan, S.C.; Marczewski, S.E. Methionine, Homocysteine, One Carbon Metabolism and Fetal Growth. Rev. Endocr. Metab. Disord. 2012, 13, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Garrow, T.A.; Dong, X.; Luchini, D.N.; Loor, J.J. Hepatic Activity and Transcription of Betaine-Homocysteine Methyltransferase, Methionine Synthase, and Cystathionine Synthase in Periparturient Dairy Cows Are Altered to Different Extents by Supply of Methionine and Choline. J. Nutr. 2017, 147, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Menchini, R.J.; Chaudhry, F.A. Multifaceted Regulation of the System A Transporter Slc38a2 Suggests Nanoscale Regulation of Amino Acid Metabolism and Cellular Signaling. Neuropharmacology 2019, 161, 107789. [Google Scholar] [CrossRef] [PubMed]
- Morotti, M.; Zois, C.E.; El-Ansari, R.; Craze, M.L.; Rakha, E.A.; Fan, S.J.; Valli, A.; Haider, S.; Goberdhan, D.C.I.; Green, A.R.; et al. Increased Expression of Glutamine Transporter SNAT2/SLC38A2 Promotes Glutamine Dependence and Oxidative Stress Resistance, and Is Associated with Worse Prognosis in Triple-Negative Breast Cancer. Br. J. Cancer 2021, 124, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.R.; Oh, Y.J.; Kang, S.W.; Lee, E.B.; Lee, W.W. Role of SLC7A5 in Metabolic Reprogramming of Human Monocyte/Macrophage Immune Responses. Front. Immunol. 2018, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, L.; Scalise, M.; Galluccio, M.; Pochini, L.; Albanese, L.M.; Indiveri, C. LAT1 Is the Transport Competent Unit of the LAT1/CD98 Heterodimeric Amino Acid Transporter. Int. J. Biochem. Cell Biol. 2015, 67, 25–33. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhang, N.; Zhao, R. Dexamethasone Downregulates SLC7A5 Expression and Promotes Cell Cycle Arrest, Autophagy and Apoptosis in BeWo Cells: DEXAMETHASONE REGULATES SLC7A5 EXPRESSION. J. Cell. Physiol. 2016, 231, 233–242. [Google Scholar] [CrossRef]
- Zhang, L.; Duan, Y.; Guo, Q.; Wang, W.; Li, F. A Selectively Suppressing Amino Acid Transporter: Sodium-Coupled Neutral Amino Acid Transporter 2 Inhibits Cell Growth and Mammalian Target of Rapamycin Complex 1 Pathway in Skeletal Muscle Cells. Anim. Nutr. 2020, 6, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, W.; Zhou, Y.; Li, F.; Wei, H.; Peng, J. Recent Advances in Understanding Amino Acid Sensing Mechanisms That Regulate MTORC1. Int. J. Mol. Sci. 2016, 17, 1636. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Li, S.; Zou, Y.; Zhao, F.Q.; Liu, J.; Liu, H. Lysine Stimulates Protein Synthesis by Promoting the Expression of ATB0,+ and Activating the MTOR Pathway in Bovine Mammary Epithelial Cells. J. Nutr. 2018, 148, 1426–1433. [Google Scholar] [CrossRef]
- Tokunaga, C.; Yoshino, K.; Yonezawa, K. MTOR Integrates Amino Acid- and Energy-Sensing Pathways. Biochem. Biophys. Res. Commun. 2004, 313, 443–446. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, W.; Niu, J.; Yang, G.; Luo, J.; Lan, D.; Wu, J.; Li, M.; Sun, Y.; Wang, K.; et al. Heat-Shock Transcription Factor 2 Promotes Sodium Butyrate-Induced Autophagy by Inhibiting MTOR in Ulcerative Colitis. Exp. Cell Res. 2020, 388, 111820. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Chen, Y.; Cortes, I.M.; Coleman, D.N.; Dai, H.; Liang, Y.; Parys, C.; Fernandez, C.; Wang, M.; Loor, J.J. Supply of Methionine and Arginine Alters Phosphorylation of Mechanistic Target of Rapamycin (MTOR), Circadian Clock Proteins, and α-S1-Casein Abundance in Bovine Mammary Epithelial Cells. Food Funct. 2020, 11, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Conejos, J.R.V.; Ghassemi Nejad, J.; Kim, J.E.; Moon, J.O.; Lee, J.-S.; Lee, H.-G. Supplementing with L-Tryptophan Increases Medium Protein and Alters Expression of Genes and Proteins Involved in Milk Protein Synthesis and Energy Metabolism in Bovine Mammary Cells. Int. J. Mol. Sci. 2021, 22, 2751. [Google Scholar] [CrossRef]
- Wu, C.W.; Storey, K.B. MTOR Signaling in Metabolic Stress Adaptation. Biomolecules 2021, 11, 681. [Google Scholar] [CrossRef] [PubMed]
- Aramburu, J.; Ortells, M.C.; Tejedor, S.; Buxade, M.; Lopez-Rodriguez, C. Transcriptional Regulation of the Stress Response by MTOR. Sci. Signal. 2014, 7, re2. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Proud, C.G. Signaling Crosstalk between the MTOR Complexes. Translation 2014, 2, e28174. [Google Scholar] [CrossRef]
- Lapierre, H.; Lobley, G.E.; Doepel, L.; Raggio, G.; Rulquin, H.; Lemosquet, S. TRIENNIAL LACTATION SYMPOSIUM: Mammary Metabolism of Amino Acids in Dairy Cows. J. Anim. Sci. 2012, 90, 1708–1721. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, L.; Zhang, L.; Liu, L.; Yang, H.; Zhou, P.; Song, F.; Dong, G.; Chen, J.; Wang, G.; Dong, X. Effect of Heat Stress on Bovine Mammary Cellular Metabolites and Gene Transcription Related to Amino Acid Metabolism, Amino Acid Transportation and Mammalian Target of Rapamycin (mTOR) Signaling. Animals 2021, 11, 3153. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113153
Fu L, Zhang L, Liu L, Yang H, Zhou P, Song F, Dong G, Chen J, Wang G, Dong X. Effect of Heat Stress on Bovine Mammary Cellular Metabolites and Gene Transcription Related to Amino Acid Metabolism, Amino Acid Transportation and Mammalian Target of Rapamycin (mTOR) Signaling. Animals. 2021; 11(11):3153. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113153
Chicago/Turabian StyleFu, Lin, Li Zhang, Li Liu, Heng Yang, Peng Zhou, Fan Song, Guozhong Dong, Juncai Chen, Gaofu Wang, and Xianwen Dong. 2021. "Effect of Heat Stress on Bovine Mammary Cellular Metabolites and Gene Transcription Related to Amino Acid Metabolism, Amino Acid Transportation and Mammalian Target of Rapamycin (mTOR) Signaling" Animals 11, no. 11: 3153. https://0-doi-org.brum.beds.ac.uk/10.3390/ani11113153