
Adverse Effects of Single Neutrophil Extracellular Trap-Derived Components on Bovine Sperm Function


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Acquisition and Ethical Declarations
2.2. Study Site
2.3. Reagents
2.4. Sperm Selection
2.5. Isolation of Bovine PMN
2.6. PMN/Sperm Co-Cultures and NET Visualisation via Scanning Electronic Microscopy (SEM) Analysis
2.7. Immunofluorescence for Neutrophil Elastase
2.8. Individual NET-Derived Component Exposure to Bovine Sperm
2.8.1. Sperm Membrane Integrity
2.8.2. Sperm Acrosome Integrity
2.8.3. Lipid Peroxidation
2.8.4. Membrane Phospholipid Disorder
2.9. Flow Cytometry
2.10. Statistical Analyses
3. Results
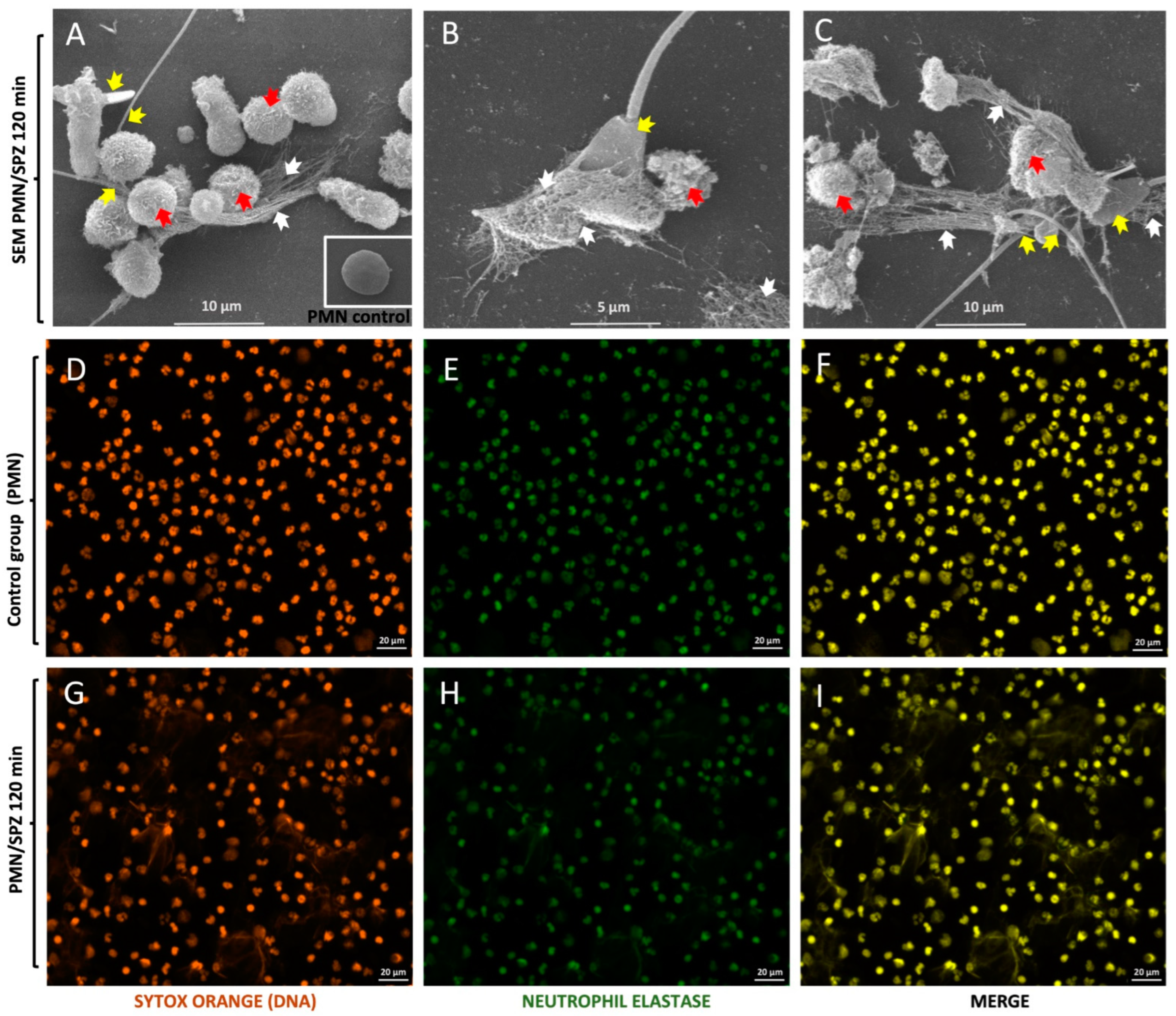
3.1. Visualisation of Bovine Sperm-Induced NETs
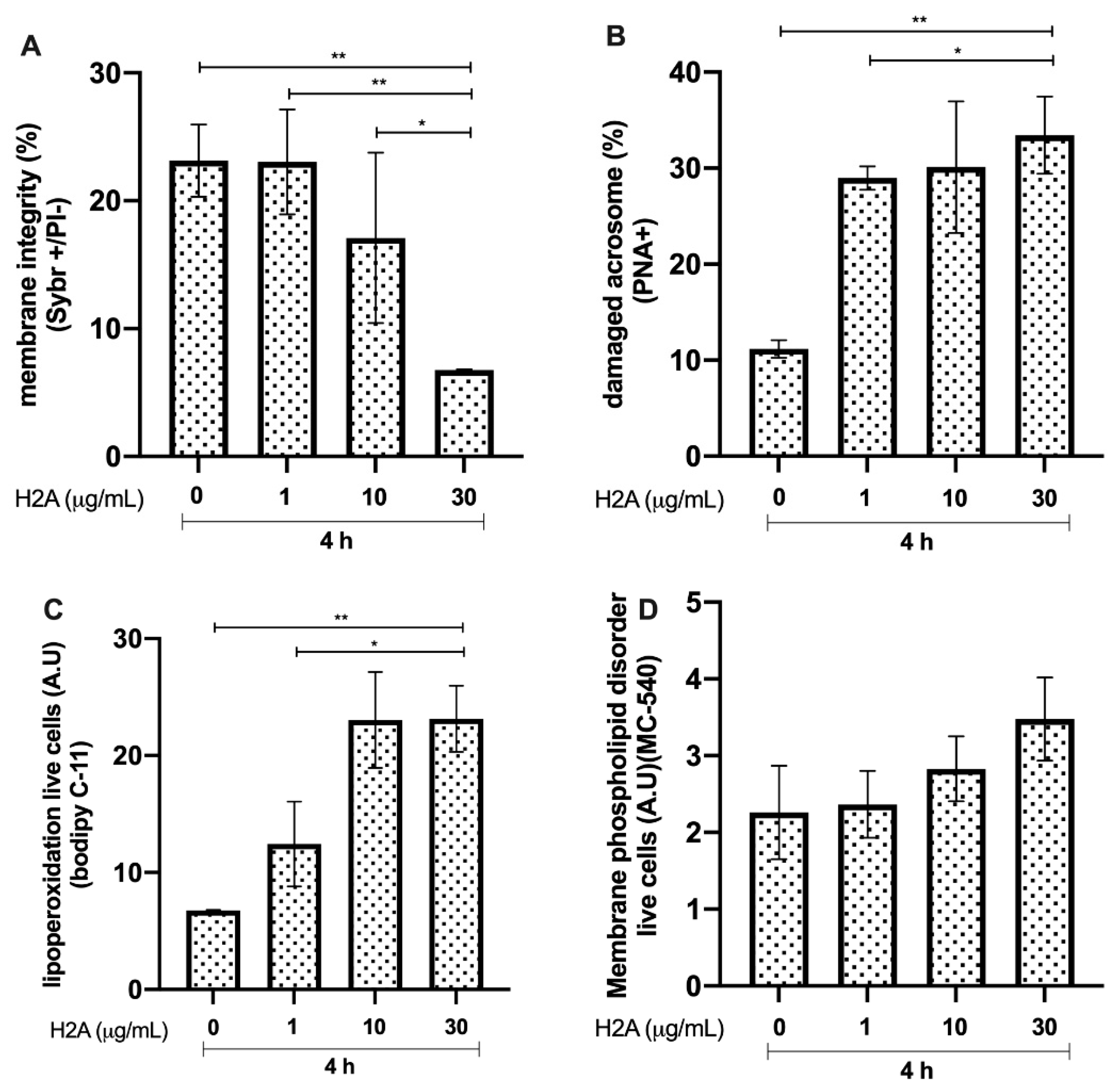
3.2. Effects of H2A on Bovine Sperm Viability
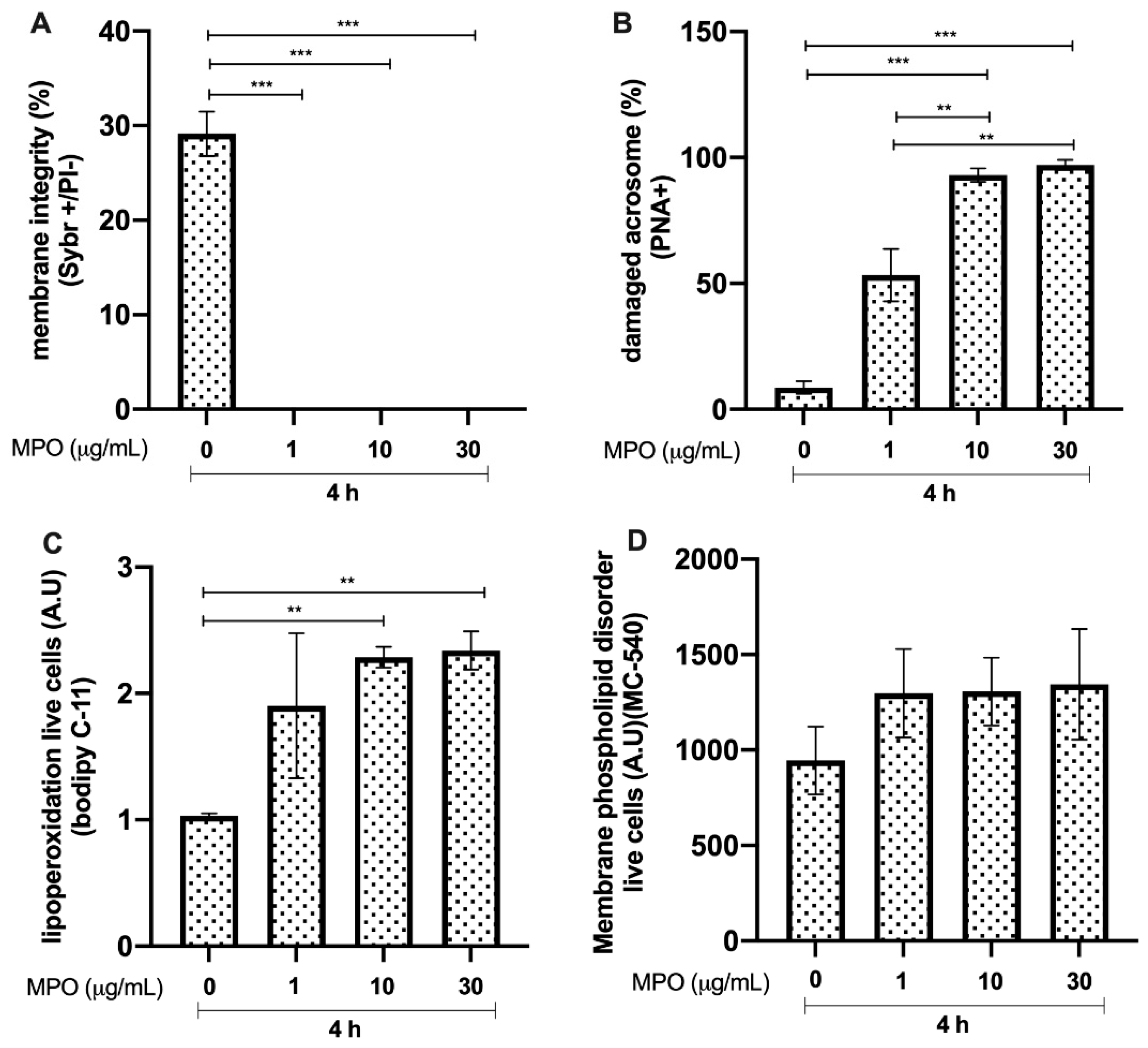
3.3. Effects of MPO on Bull Sperm
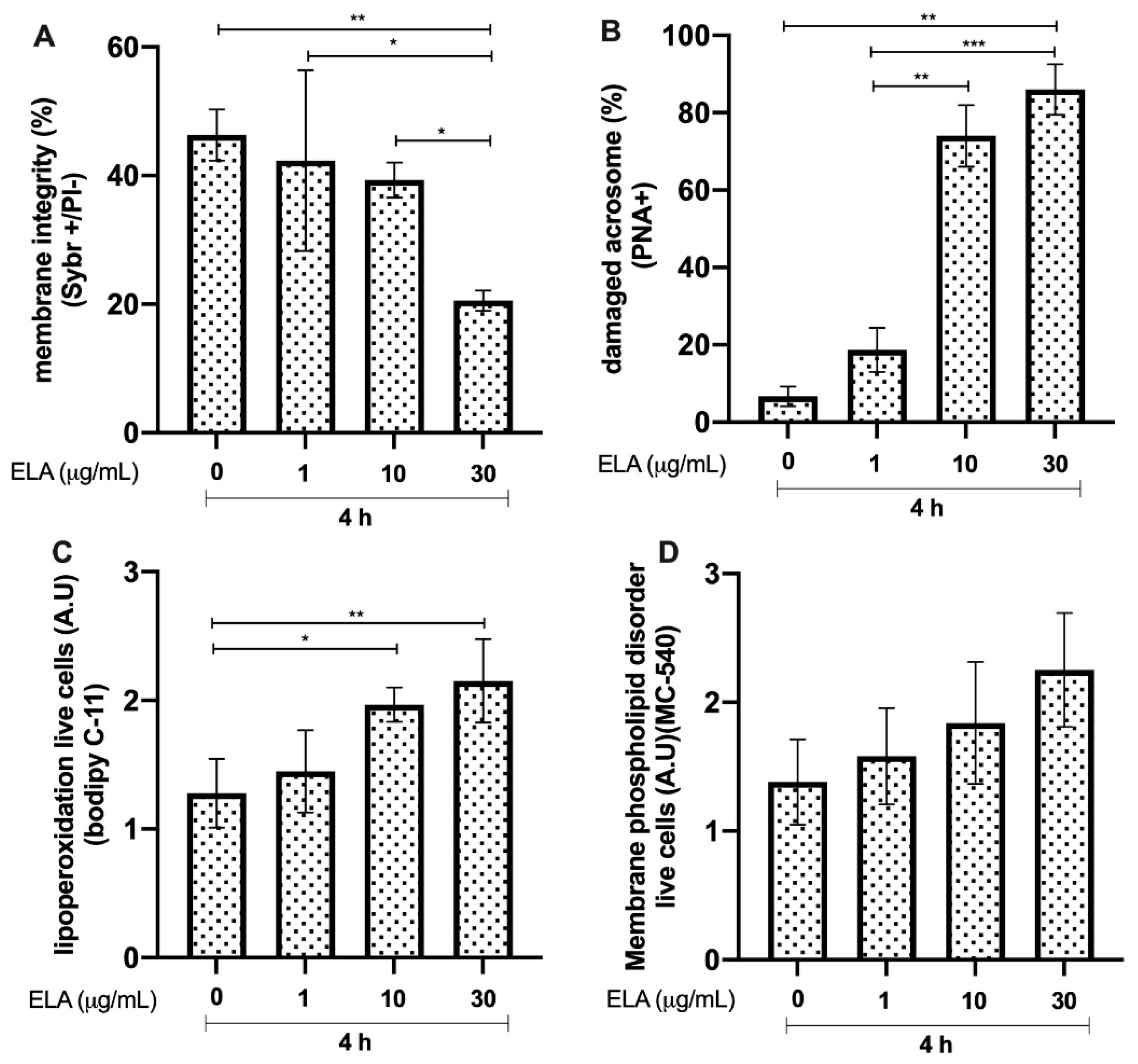
3.4. Effects of ELA on Bull Sperm
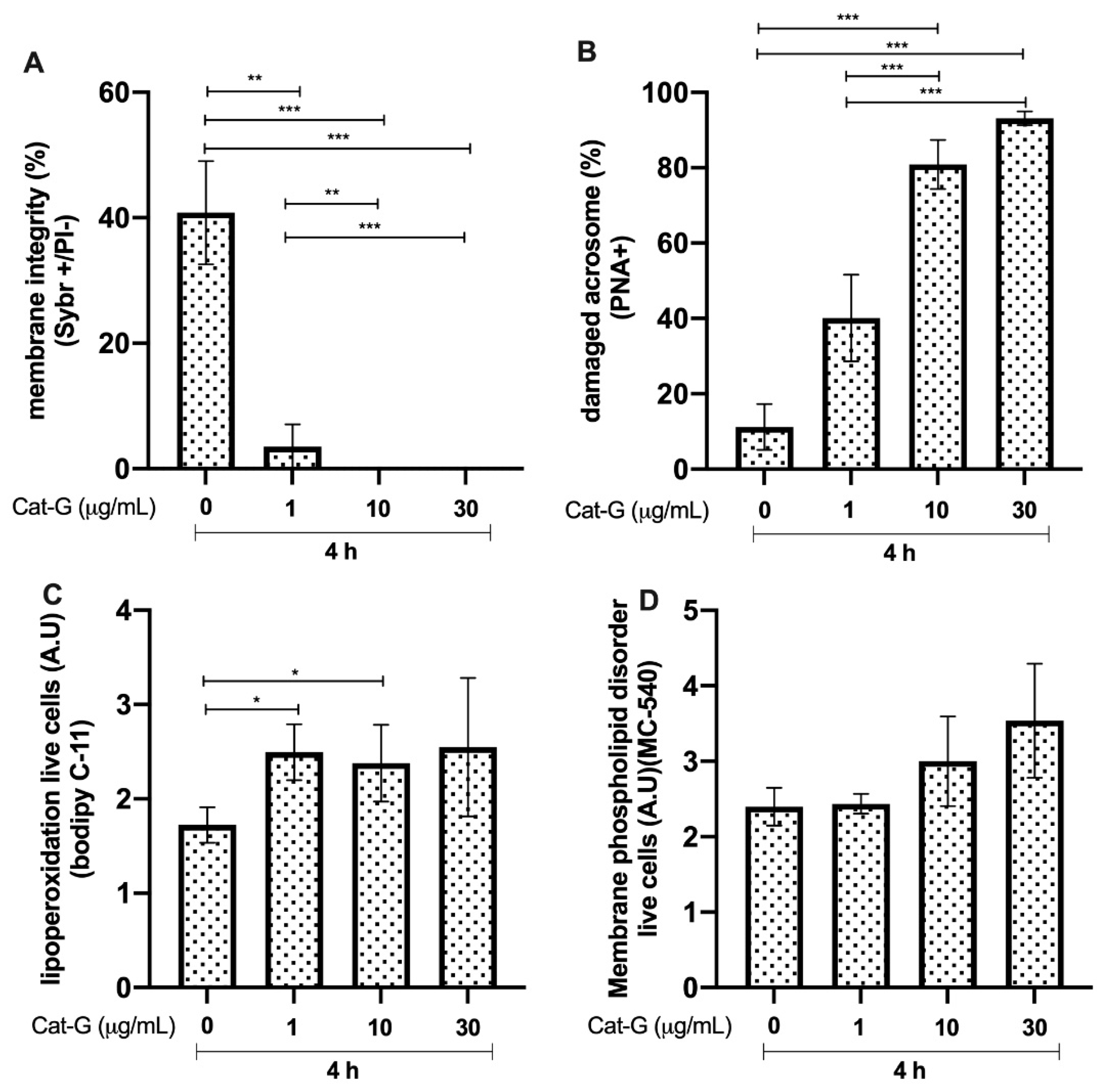
3.5. Effects of Cat-G on Bull Sperm
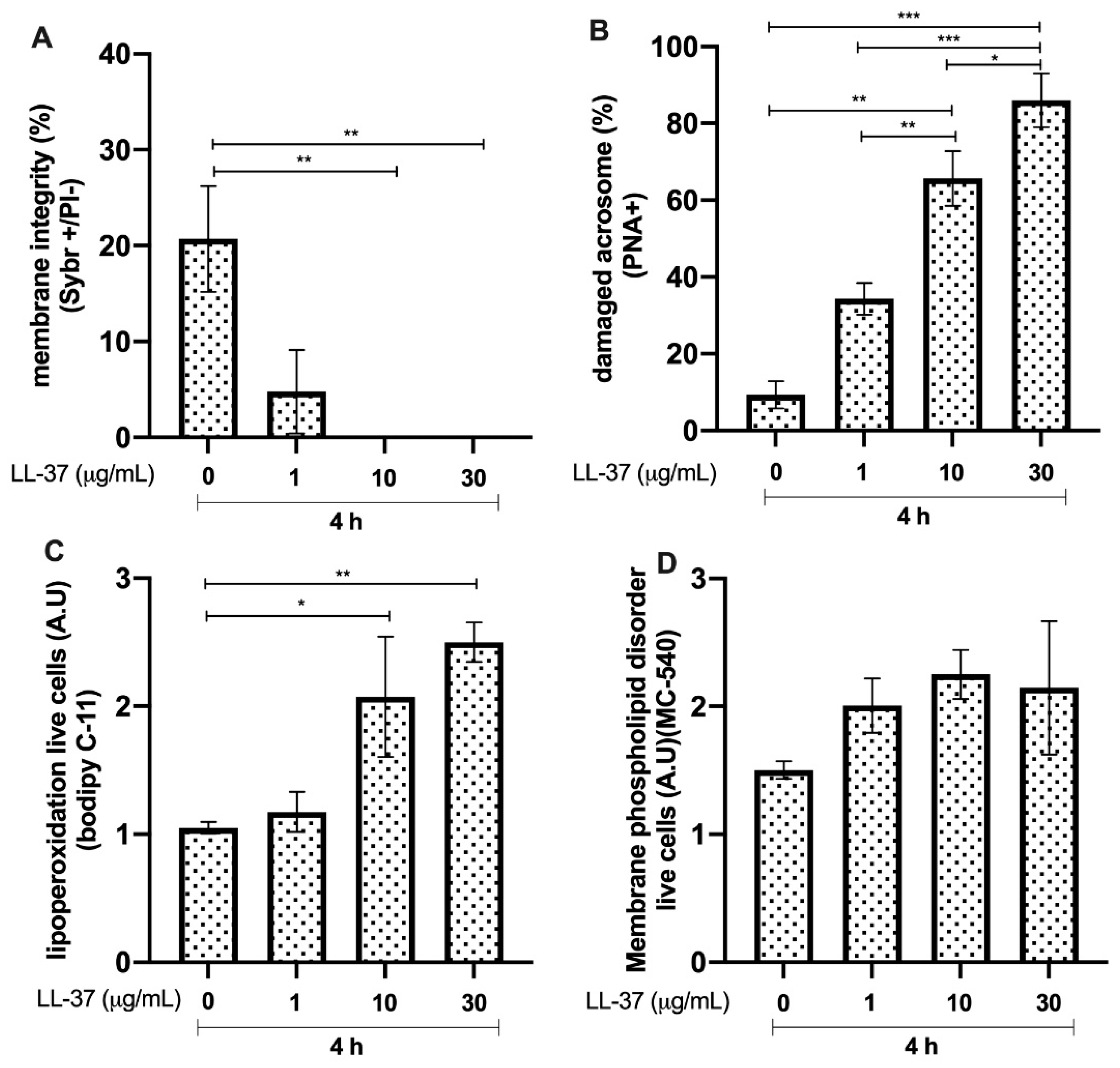
3.6. Effects of LL-37 on Bull Sperm
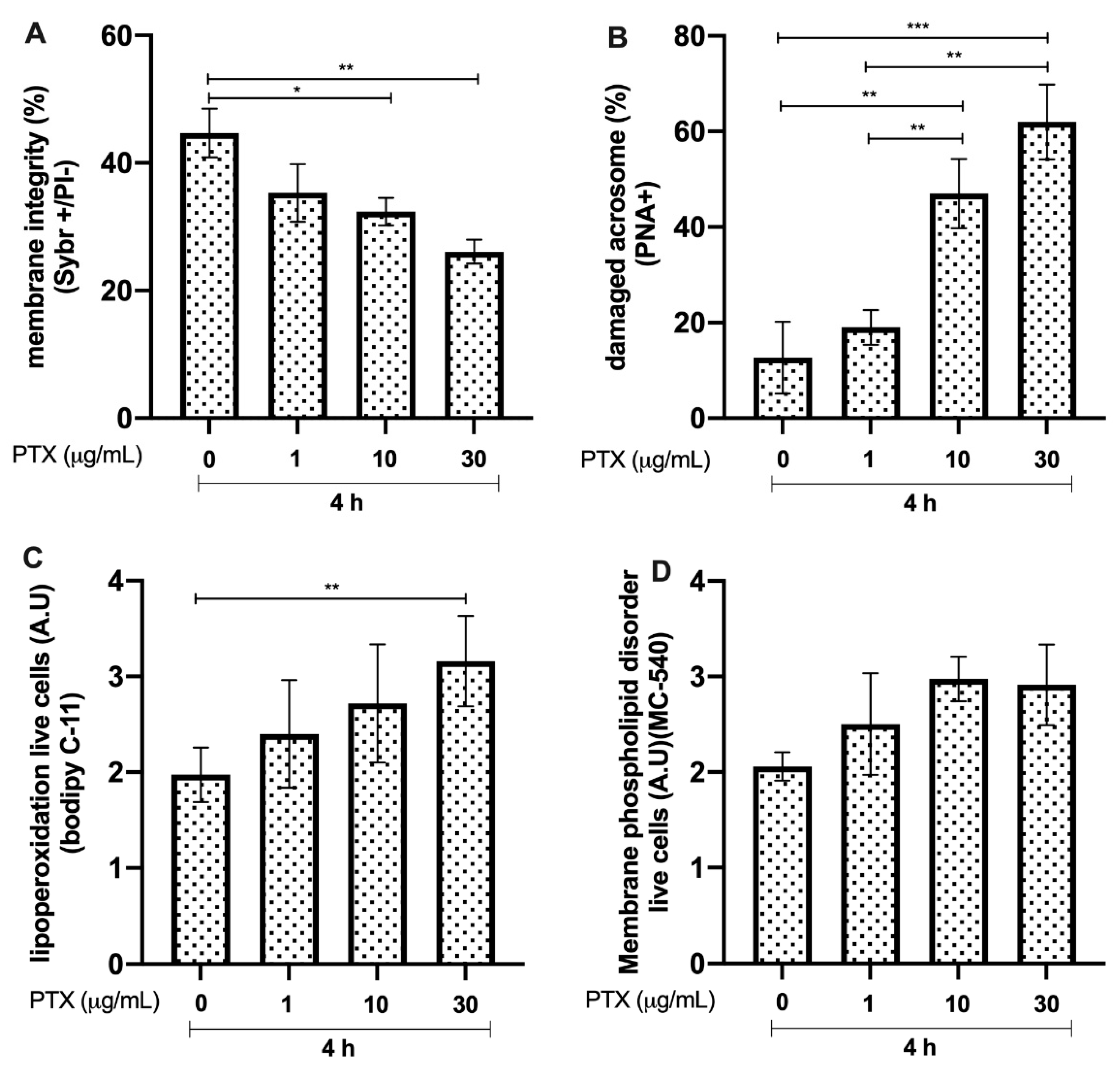
3.7. Effects of PTX on Bull Sperm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fichtner, T.; Kotarski, F.; Gärtner, U.; Conejeros, I.; Hermosilla, C.; Wrenzycki, C.; Taubert, A. Bovine sperm samples induce different NET phenotypes in a NADPH Oxidase-, PAD4-, and Ca++-dependent process. Biol. Reprod. 2020, 102, 902–914. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martinez, H.; Saravia, F.; Wallgren, M.; Martinez, E.A.; Sanz, L.; Roca, J.; Vazquez, J.M.; Calvete, J.J. Spermadhesin PSP-I/PSP-II Heterodimer Induces Migration of Polymorphonuclear Neutrophils into the Uterine Cavity of the Sow. J. Reprod. Immunol. 2010, 84, 57–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, D.; Schuberth, H.J.; Coy, P.; Taylor, U. Sperm Interactions from Insemination to Fertilization: Sperm Interactions. Reprod. Domest. Anim. 2008, 43 (Suppl. 5), 2–11. [Google Scholar] [CrossRef] [PubMed]
- Bader, H. An Investigation of Sperm Migration into the Oviducts of the Mare. J. Reprod. Fertil. Suppl. 1982, 32, 59–64. [Google Scholar] [PubMed]
- Katila, T. Post-Mating Inflammatory Responses of the Uterus: Post-Mating Inflammation. Reprod. Domest. Anim. 2012, 47 (Suppl. 5), 31–41. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Dąbrowska, D.; Jabłońska, E.; Garley, M.; Ratajczak-Wrona, W.; Iwaniuk, A. New aspects of the biology of neutrophil extracellular traps. Scand. J. Immunol. 2016, 84, 317–322. [Google Scholar] [CrossRef]
- Delgado-Rizo, V.; Martínez-Guzmán, M.A.; Iñiguez-Gutierrez, L.; García-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil extracellular traps and its implications in inflammation: An overview. Front. Immunol. 2017, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Zambrano, F.; Silva, L.; Uribe, P.; Gärtner, U.; Taubert, A.; Schulz, M.; Sánchez, R.; Hermosilla, C. SOCE-inhibitor reduced human sperm-induced formation of neutrophil extracellular traps. Reproduction. 2021, 161, 21–29. [Google Scholar] [CrossRef]
- Zambrano, F.; Namuncura, C.; Uribe, P.; Schulz, M.; Pezo, F.; Burgos, R.A.; Taubert, A.; Hermosilla, C.; Sanchez, R. Swine spermatozoa trigger aggregated neutrophil extracellular traps leading to adverse effects on sperm function. J. Reprod. Immunol. 2021, 146, 103339. [Google Scholar] [CrossRef]
- Daigo, K.; Takamatsu, Y.; Hamakubo, T. The protective effect against extracellular histones afforded by Long-Pentraxin PTX3 as a regulator of NETs. Front. Immunol. 2016, 7, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conejeros, I.; Velásquez, Z.D.; Grob, D.; Zhou, E.; Salecker, H.; Hermosilla, C.; Taubert, A. Histone H2A and Bovine Neutrophil Extracellular Traps Induce Damage of Besnoitia Besnoiti-Infected Host Endothelial Cells but Fail to Affect Total Parasite Proliferation. Biology 2019, 8, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, F.; Aguila, L.; Arias, M.E.; Sánchez, R.; Felmer, R. Improved preimplantation development of bovine ICSI embryos generated with spermatozoa pretreated with membrane-destabilizing agents Lysolecithin and Triton X-100. Theriogenology 2016, 86, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- Koppers, A.J.; De Iuliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of Mitochondrial Reactive Oxygen Species in the Generation of Oxidative Stress in Spermatozoa. J. Clin. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef] [Green Version]
- Steckler, A.; Stout, T.A.E.; Durandt, C.; Nöthling, J.O. Validation of merocyanine 540 staining as a technique for assessing capacitation-related membrane destabilization of fresh dog sperm. Theriogenology 2015, 83, 1451–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alghamdi, A.S.; Foster, D.N. Seminal DNase Frees Spermatozoa Entangled in Neutrophil Extracellular Traps. Biol. Reprod. 2005, 73, 1174–1181. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Yu, T.; Wang, J.; Wang, C.; Liu, X.; Han, Z.; Zhang, X.; Zhang, Y.; Ouyang, H.; Yang, Z. Swine Sperm Induces Neutrophil Extracellular Traps That Entangle Sperm and Embryos. Reproduction. 2020, 160, 217–225. [Google Scholar] [CrossRef]
- Mateo-Otero, Y.; Zambrano, F.; Catalán, J.; Sánchez, R.; Yeste, M.; Miro, J.; Fernandez-Fuertes, B. Seminal Plasma, and Not Sperm, Induces Time and Concentration-Dependent Neutrophil Extracellular Trap Release in Donkeys. Equine Vet. J. 2022, 54, 415–426. [Google Scholar] [CrossRef]
- Fichtner, T.; Kotarski, F.; Hermosilla, C.; Taubert, A.; Wrenzycki, C. Semen Extender and Seminal Plasma Alter the Extent of Neutrophil Extracellular Traps (NET) Formation in Cattle. Theriogenology 2021, 160, 72–80. [Google Scholar] [CrossRef]
- Thomas, G.M.; Carbo, C.; Curtis, B.R.; Martinod, K.; Mazo, I.B.; Schatzberg, D.; Cifuni, S.M.; Fuchs, T.A.; von Andrian, U.H.; Hartwig, J.H.; et al. Extracellular DNA traps are associated with the pathogenesis of TRALI in humans and mice. Blood 2012, 119, 6335–6343. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.C.; Chuang, Y.H.; Tsai, Y.F.; Yu, H.P. Role of neutrophil extracellular traps following injury. Shock 2014, 41, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, F.; Carrau, T.; Gärtner, U.; Seipp, A.; Taubert, A.; Felmer, R.; Sanchez, R.; Hermosilla, C. Leukocytes coincubated with human sperm trigger classic neutrophil extracellular traps formation, reducing sperm motility. Fertil. Steril. 2016, 106, 1053–1060.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, E.; Silva, L.M.R.; Conejeros, I.; Velásquez, Z.D.; Hirz, M.; Gärtner, U.; Jacquiet, P.; Taubert, A.; Hermosilla, C. Besnoitia Besnoiti Bradyzoite Stages Induce Suicidal- and Rapid Vital-NETosis. Parasitology 2020, 147, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Schulz, M.; Zambrano, F.; Schuppe, H.C.; Wagenlehner, F.; Taubert, A.; Ulrich, G.; Sánchez, R.; Hermosilla, C. Determination of Leucocyte Extracellular Traps (ETs) in Seminal Fluid (Ex Vivo) in Infertile Patients-A Pilot Study. Andrologia 2019, 51, e13356. [Google Scholar] [CrossRef]
- Zambrano, F.; Schulz, M.; Pilatz, A.; Wagenlehner, F.; Schuppe, H.C.; Conejeros, I.; Uribe, P.; Taubert, A.; Sánchez, R.; Hermosilla, C. Increase of Leucocyte-Derived Extracellular Traps (ETs) in Semen Samples from Human Acute Epididymitis Patients-a Pilot Study. J. Assist. Reprod. Genet. 2020, 37, 2223–2231. [Google Scholar] [CrossRef]
- Piasecka, M.; Fraczek, M.; Gaczarzewicz, D.; Gill, K.; Szumala-Kakol, A.; Kazienko, A.; Laszczynska, M.; Lenart, S.; Beutin, L.; Kurpisz, M. Novel morphological findings of human sperm removal by leukocytes in in vivo and in vitro conditions: Preliminary study. Am. J. Reprod. Immunol. 2014, 72, 348–358. [Google Scholar] [CrossRef]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil Extracellular Traps Contain Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida Albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [Green Version]
- Saffarzadeh, M.; Juenemann, C.; Queisser, M.A.; Lochnit, G.; Barreto, G.; Galuska, S.P.; Lohmeyer, J.; Preissner, K.T. Neutrophil Extracellular Traps Directly Induce Epithelial and Endothelial Cell Death: A Predominant Role of Histones. PLoS ONE. 2012, 7, e32366. [Google Scholar] [CrossRef]
- Donis-Maturano, L.; Sánchez-Torres, L.E.; Cerbulo-Vázquez, A.; Chacón-Salinas, R.; García-Romo, G.S.; Orozco-Uribe, M.C.; Yam-Puc, J.C.; González-Jiménez, M.A.; Paredes-Vivas, Y.L.; Calderón-Amador, J.; et al. Prolonged exposure to neutrophil extracellular traps can induce mitochondrial damage in macrophages and dendritic cells. Springerplus 2015, 4, 161. [Google Scholar] [CrossRef] [Green Version]
- Bradley, P.P.; Christensen, R.D.; Rothstein, G. Cellular and Extracellular Myeloperoxidase in Pyogenic Inflammation. Blood 1982, 60, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Kudo, T.; Kigoshi, H.; Hagiwara, T.; Takino, T.; Yamazaki, M.; Yui, S. Cathepsin G, a Neutrophil Protease, Induces Compact Cell-Cell Adhesion in MCF-7 Human Breast Cancer Cells. Mediat. Inflamm. 2009, 2009, 850940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchner, T.; Möller, S.; Klinger, M.; Solbach, W.; Laskay, T.; Behnen, M. The impact of various reactive oxygen species on the formation of neutrophil extracellular traps. Mediat. Inflamm. 2012, 2012, 849136. [Google Scholar] [CrossRef] [PubMed]
- Odeberg, H.; Olsson, I. Antibacterial Activity of Cationic Proteins from Human Granulocytes. J. Clin. Investig. 1975, 56, 1118–1124. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.E.; Bowdish, D.M.E.; Cosseau, C.; Hancock, R.E.W.; Davidson, D.J. Apoptosis of Airway Epithelial Cells: Human Serum Sensitive Induction by the Cathelicidin LL-37: Human Serum Sensitive Induction by the Cathelicidin LL-37. Am. J. Respir. Cell Mol. Biol. 2006, 34, 399–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshari, R.S.; Jyoti, A.; Kumar, S.; Dubey, M.; Verma, A.; Srinag, B.S.; Krishnamurthy, H.; Barthwal, M.K.; Dikshit, M. Neutrophil extracellular traps contain mitochondrial as well as nuclear DNA and exhibit inflammatory potential. Cytometry A 2012, 81, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Uribe, P.; Meriño, J.; Matus, C.E.; Schulz, M.; Zambrano, F.; Villegas, J.V.; Conejeros, I.; Taubert, A.; Hermosilla, C.; Sánchez, R. Autophagy is activated in human spermatozoa subjected to oxidative stress and its inhibition impairs sperm quality and promotes cell death. Hum. Reprod. 2022, 37, 680–695. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef]
- Yánez-Ortiz, I.; Catalán, J.; Mateo-Otero, Y.; Dordas-Perpinyà, M.; Gacem, S.; Yeste, N.; Bassols, A.; Yeste, M.; Miró, J. Extracellular Reactive Oxygen Species (ROS) Production in Fresh Donkey Sperm Exposed to Reductive Stress, Oxidative Stress and NETosis. Antioxidants 2021, 10, 1367. [Google Scholar] [CrossRef]
- Greene, C.M.; McElvaney, N.G. Proteases and antiproteases in chronic neutrophilic lung disease—Relevance to drug discovery: Proteases and antiproteases in lung disease. Br. J. Pharmacol. 2009, 158, 1048–1058. [Google Scholar] [CrossRef] [Green Version]
- Kumaresan, A.; Kadirvel, G.; Bujarbaruah, K.M.; Bardoloi, R.K.; Das, A.; Kumar, S.; Naskar, S. Preservation of Boar Semen at 18 Degrees C induces lipid peroxidation and apoptosis like changes in spermatozoa. Anim. Reprod. Sci. 2009, 110, 162–171. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, C.; Rivera-Concha, R.; Pezo, F.; Uribe, P.; Schulz, M.; Sánchez, R.; Hermosilla, C.; Taubert, A.; Gärtner, U.; Zambrano, F. Adverse Effects of Single Neutrophil Extracellular Trap-Derived Components on Bovine Sperm Function. Animals 2022, 12, 1308. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101308
Moya C, Rivera-Concha R, Pezo F, Uribe P, Schulz M, Sánchez R, Hermosilla C, Taubert A, Gärtner U, Zambrano F. Adverse Effects of Single Neutrophil Extracellular Trap-Derived Components on Bovine Sperm Function. Animals. 2022; 12(10):1308. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101308
Chicago/Turabian StyleMoya, Claudia, Rodrigo Rivera-Concha, Felipe Pezo, Pamela Uribe, Mabel Schulz, Raúl Sánchez, Carlos Hermosilla, Anja Taubert, Ulrich Gärtner, and Fabiola Zambrano. 2022. "Adverse Effects of Single Neutrophil Extracellular Trap-Derived Components on Bovine Sperm Function" Animals 12, no. 10: 1308. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12101308