
Aliens Coming by Ships: Distribution and Origins of the Ocellated Skink Populations in Peninsular Italy

 ,
, 
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Molecular Analyses
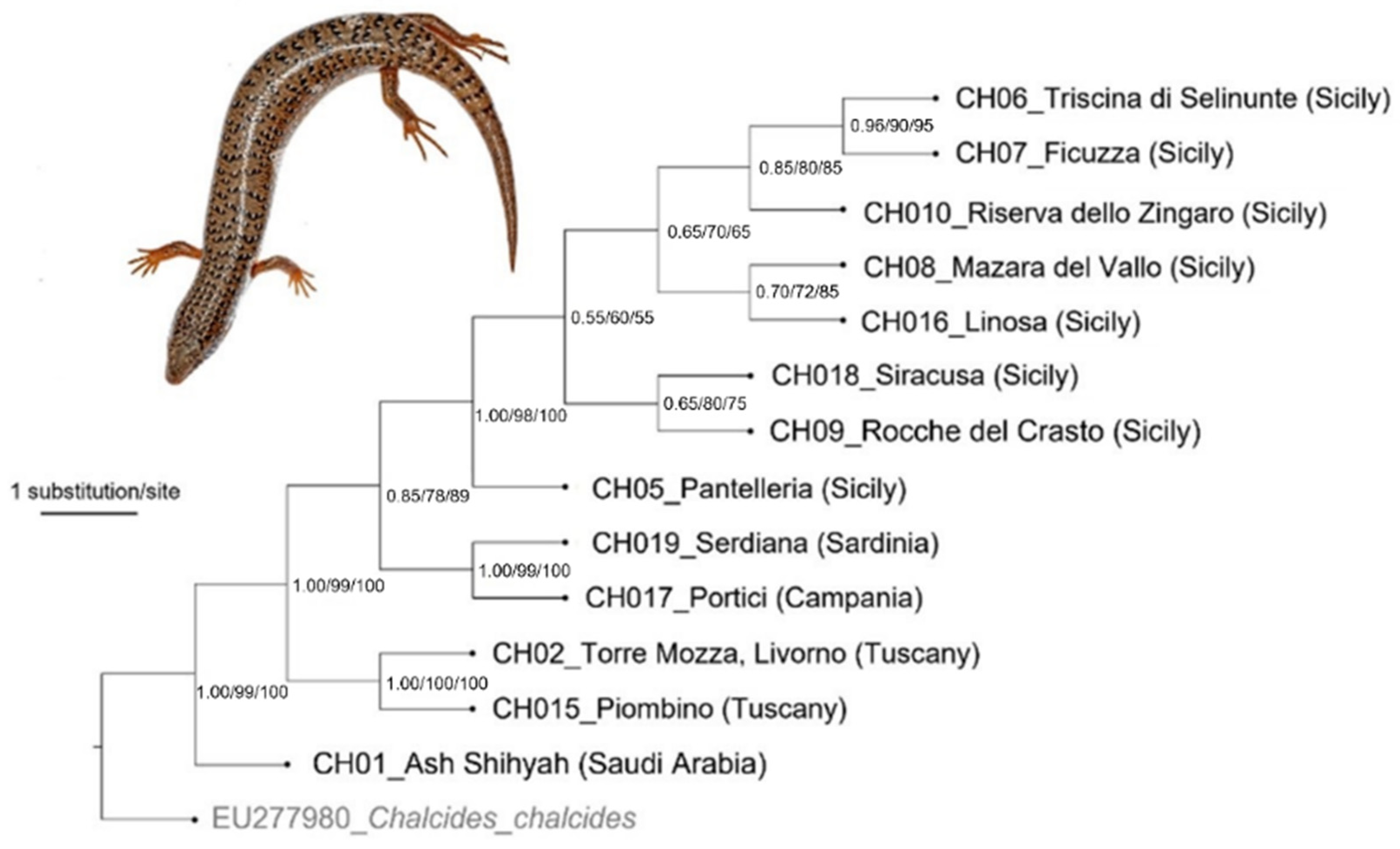
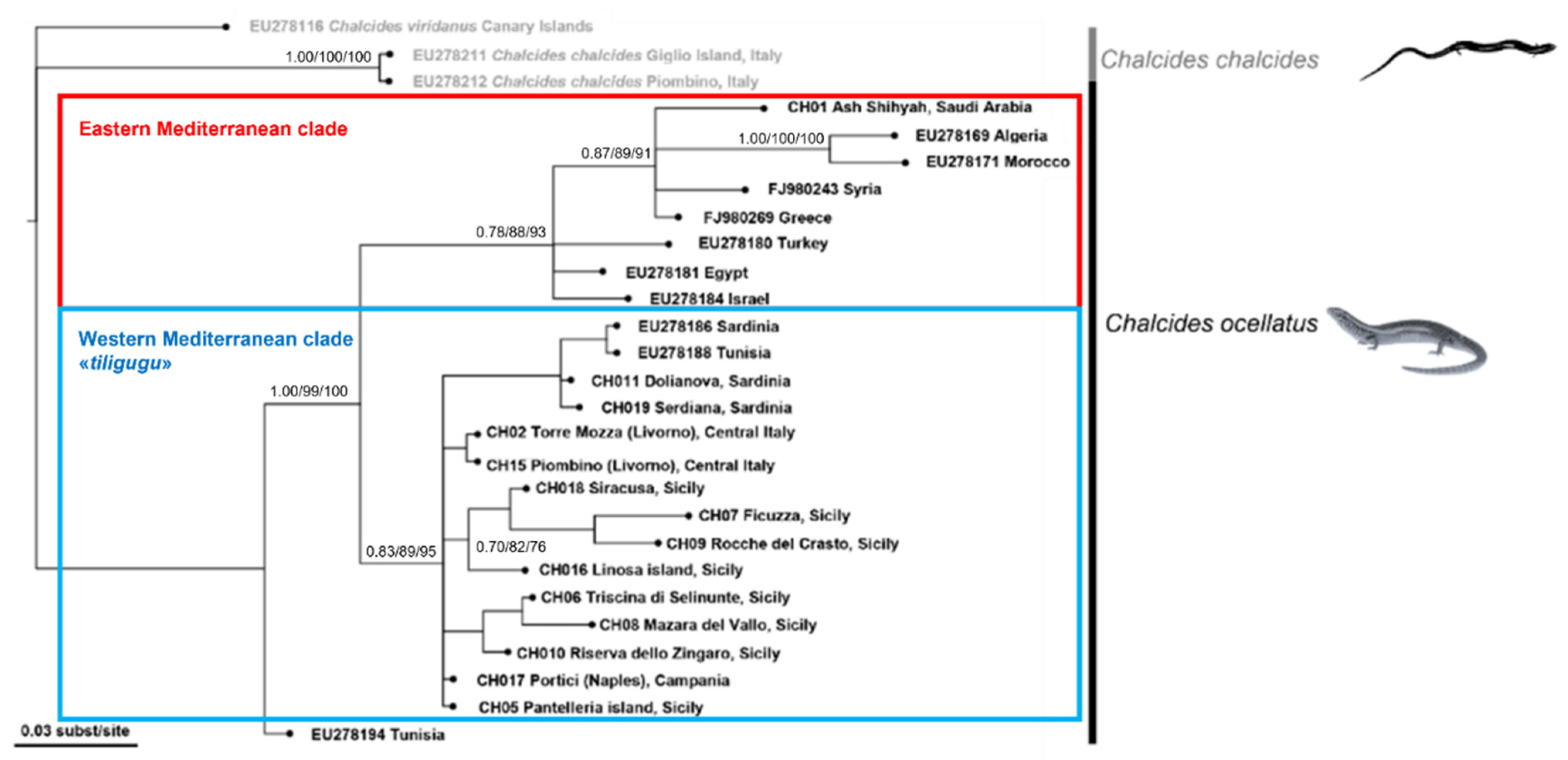
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seebens, H. Invasion ecology: Expanding trade and the dispersal of alien species. Curr. Biol. 2019, 29, 120–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tingley, R.; García-Díaz, P.; Arantes, C.R.R.; Cassey, P. Integrating transport pressure data and species distribution models to estimate invasion risk for alien stowaways. Ecography 2018, 41, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Inghilesi, A.F.; Mazza, G.; De Silva, J.; Francardi, V.; Nuccitelli, L.; Pennacchio, F.; Sinatra, G.; Sposimo, P.; Gherardi, F. Valutazione dell’incidenza degli aeroporti di Fiumicino e Ciampino e del porto di Civitavecchia sull’introduzione di fauna alloctona. In Alieni, la Minaccia Delle Specie Alloctone per la Biodiversità del Lazio; Monaco, A., Ed.; Palombi Editori: Roma, Italia, 2014; pp. 98–115. [Google Scholar]
- Turbelin, A.J.; Malamud, B.D.; Francis, R.A. Mapping the global state of invasive alien species: Patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 2017, 26, 78–92. [Google Scholar] [CrossRef] [Green Version]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species in an era of globalization. J. Anim. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Hulme, P.E. Invasion pathways at a crossroad: Policy and research challenges for managing alien species introductions. J. Anim. Ecol. 2015, 52, 1418–1424. [Google Scholar] [CrossRef]
- van den Burg, M.P.; Brisbane, J.L.; Knapp, C.R. Post-hurricane relief facilitates invasion and establishment of two invasive alien vertebrate species in the Commonwealth of Dominica, West Indies. Biol. Invasions 2020, 22, 195–203. [Google Scholar] [CrossRef]
- Bertolino, S.; Ancillotto, L.; Bartolommei, P.; Benassi, G.; Capizzi, D.; Gasperini, S.; Lucchesi, M.; Mori, E.; Scillitani, L.; Sozio, G.; et al. A framework for prioritising present and potentially invasive mammal species for a national list. NeoBiota 2020, 62, 31–54. [Google Scholar] [CrossRef]
- Karnakowski, W. Quarantine pests intercepted in consignments of ornamental plants imported into Poland in 1993/1998. OEPP/EPPO Bull. 1999, 29, 45–49. [Google Scholar] [CrossRef]
- Di Nicola, M.; Cavigioli, L.; Luiselli, L.; Andreone, F. Anfibi & Rettili d’Italia. Edizione Aggiornata; Historia Naturae; Edizioni Belvedere: Latina, Italy, 2021; p. 576. [Google Scholar]
- Mares, G.; Novarini, N. A likely population of the alien gecko Mediodactylus kotschyi (Steindachner, 1870) in the province of Belluno (Northeastern Italian Alps). Boll. Mus. Stor. Nat. Venezia 2020, 71, 83–88. [Google Scholar]
- Mezzasalma, M.; Odierna, G.; Maio, N.; Petraccioli, A.; Picariello, O.; Guarino, F.M. Habitat features and distribution of Hemidactylus turcicus and Tarentola mauritanica in Campania (southern Italy). In Atti VIII Congresso Nazionale Societas Herpetologica Italica; Di Tizio, L., Di Cerbo, A.R., Di Francesco, N., Cameli, A., Eds.; Ianieri Edizioni: Pescara, Italy, 2010; pp. 129–133. [Google Scholar]
- Mori, E.; Plebanim, M. First records of Moorish gecko Tarentola mauritanica and Turkish gecko Hemidactylus turcicus (Squamata, Gekkonidae) in the Southern Metalliferous Hills, Tuscany, Italy. Atti Soc. Tosc. Sci. Nat. 2012, 119, 51–54. [Google Scholar]
- De Fatis, K.T.; Di Nicola, M.R.; Lebech Nassling Iversen, D. Prima segnalazione di Mediodactlylus kotschyi (Steindachner, 1870) per il Trentino-Alto Adige/Südtirol (Italia) (Squamata: Gekkonidae). Studi Trentini Sci. Nat. 2020, 99, 25–27. [Google Scholar]
- Andreone, F.; Angelici, F.M.; Carlino, P.; Tripepi, S.; Crottini, A. The common chameleon Chamaeleo chamaeleon in southern Italy: Evidence for allochthony of populations in Apulia and Calabria (Reptilia: Squamata: Chamaeleonidae). Ital. J. Zool. 2016, 83, 372–381. [Google Scholar] [CrossRef] [Green Version]
- Basso, R.; Vannuccini, M.L.; Nerva, L.; Mazza, G.; Seno, M.; Mori, E. Multiple origins of the common chameleon in southern Italy. Herpetozoa 2019, 32, 11–19. [Google Scholar] [CrossRef]
- Faraone, F.P.; Barraco, L.; Giacalone, G.; Muscarella, C.; Schifani, E.; Vecchioni, L. First records of the Brahminy blind snake, Indotyphlops braminus (Daudin, 1803) (Squamata: Typhlopidae), in Italy. Herpetol. Notes 2019, 12, 1225–1229. [Google Scholar]
- Paolino, G.; Scotti, R.; Grano, M. First detection of the “flowerpot snake” Indotyphlops braminus (Daudin, 1803) (Serpentes Typhlopidae) in Ischia (Italy): A new possible invasive species. Biodiv. J. 2019, 10, 321–324. [Google Scholar] [CrossRef]
- Kornilios, P.; Kyriazi, P.; Poulakakis, N.; Kumlutaş, Y.; Ilgaz, Ç.; Mylonas, M.; Lymberakis, P. Phylogeography of the ocellated skink Chalcides ocellatus (Squamata, Scincidae), with the use of mtDNA sequences: A hitch-hiker’s guide to the Mediterranean. Mol. Phylogenet. Evol. 2010, 54, 445–456. [Google Scholar] [CrossRef]
- Lavin, B.R.; Papenfuss, T.J. The phylogenetic position of Chalcides ocellatus (Squamata: Scincidae) from Yemen and Somalia. Zootaxa 2012, 3221, 26–36. [Google Scholar] [CrossRef]
- Carranza, S.; Arnold, E.N.; Geniez, P.; Roca, J.; Mateo, J.A. Radiation, multipledispersal and parallelism in the skinks, Chalcides and Sphenops (Squamata: Scincidae), with comments on Scincus and Scincopus the age of the Sahara Desert. Mol. Phylogenet. Evol. 2008, 46, 1071–1094. [Google Scholar] [CrossRef]
- Sindaco, R.; Jeremčenko, V.K. The Reptiles of the Western Palearctic; Edizioni Belvedere: Latina, Italy, 2008; Volume 1. [Google Scholar]
- Stöck, M.; Grifoni, G.; Armor, N.; Scheidt, U.; Sicilia, A.; Novarini, N. On the origin of the recent herpetofauna of Sicily: Comparative phylogeography using homologous mitochondrial and nuclear genes. Zool. Anz. 2016, 261, 70–81. [Google Scholar] [CrossRef]
- Maio, N.; Mancini, D.; Mezzasalma, M.; Odierna, G.; Petraccioli, A.; Nugnes, F.; Picariello, O.L.A.; Guarino, F.M. Extinct or not extinct: The case of the Chalcides ocellatus (Squamata: Scincidae) population from the Park of the ex Bourbonic Royal Palace of Portici (Naples, Italy). Nat. Hist. Sci. 2015, 2, 43–54. [Google Scholar]
- Stoch, F.; Genovesi, P. Manuali per il Monitoraggio di Specie e Habitat di Interesse Comunitario (Direttiva 92/43/CEE) in Italia: Specie Animali; Serie Manuali e Linee Guida, 141/2016; ISPRA Publisher: Roma, Italy, 2016. [Google Scholar]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Nat. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiens, J.J.; Reeder, T.W. Phylogeny of the spiny lizards (Sceloporus) based on molecular and morphological evidence. Herpetol. Monogr. 1997, 11, 1–101. [Google Scholar] [CrossRef]
- Leaché, A.D.; Reeder, T.W. Molecular systematics of the eastern fence lizard (Sceloporus undulatus): A comparison of parsimony, likelihood, and Bayesian approaches. Syst. Biol. 2002, 51, 44–68. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Baratti, M.; Goti, E.; Messana, G. High level of genetic differentiation in the marine isopod Sphaeroma terebrans (Crustacea Isopoda Sphaeromatidae) as inferred by mitochondrial DNA analysis. J. Exp. Mar. Biol. Ecol. 2005, 315, 225–234. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Nat. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [Green Version]
- Buhay, J.E. COI-like sequences are becoming problematic in molecular systematic and DNA barcoding studies. J. Crustacean Biol. 2009, 29, 96–110. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Meth. 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: Approximate methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shackleton, J.C.; Van Andel, T.H.; Runnels, C.N. Coastal paleogeography of the central and western Mediterranean during the last 125,000 years and its archaeological implications. J. Field Archaeol. 1984, 11, 307–314. [Google Scholar]
- Fritz, U.; Harris, D.J.; Fahd, S.; Rouag, R.; Martínez, E.G.; Casalduero, A.G.; Široký, P.; Kalboussi, M.; Jdeidi, T.B.; Hundsdörfer, A.K. Mitochondrial phylogeography of Testudo graeca in the Western Mediterranean: Old complex divergence in North Africa and recent arrival in Europe. Amphibia-Reptilia 2009, 30, 63–80. [Google Scholar] [CrossRef] [Green Version]
- Guicking, D.; Joger, U.; Wink, M. Molecular phylogeography of the viperine snake Natrix maura (Serpentes: Colubridae): Evidence for strong intraspecific differentiation. Org. Divers. Evol. 2008, 8, 130–145. [Google Scholar] [CrossRef] [Green Version]
- Faraone, F.P.; Melfi, R.; Di Nicola, M.R.; Giacalone, G.; Lo Valvo, M. Phylogenetic relationships of the Italian populations of Horseshoe Whip Snake Hemorrhois hippocrepis (Serpentes, Colubridae). Acta Herpetol. 2020, 15, 129–135. [Google Scholar]
- Corona, P.; Quatrini, V.; Schirru, M.; Dettori, S.; Puletti, N. Towards the economic valuation of ecosystem production from cork oak forests in Sardinia (Italy). iForest-Biogeosci. For. 2018, 11, 660. [Google Scholar] [CrossRef] [Green Version]
- Pantaleoni, R.A.; Cocco, A.; Floris, I.; Letardi, A.; Loru, L. Going overseas: From island to continent colonization in the Mediterranean snakefly Fibla maclachlani (Albarda, 1891). BioInvasions Rec. 2019, 8, 442–451. [Google Scholar] [CrossRef]
- Capula, M.; Luiselli, L. Resource partitioning in a Mediterranean lizard community. Ital. J. Zool. 1994, 61, 173–177. [Google Scholar] [CrossRef]
- Casale, A.; Bastianini, M.; Minniti, M. Sulla presenza in Toscana di Carabus Macrothorax morbillosus Fabricius (Coleoptera, Carabidae, Carabini) e sul suo significato zoogeografico. Frustula Entomol. 1987, 10, 67–72. [Google Scholar]
- Trozzi, C.; Vaccaro, R.; Nicolo, L. Air pollutants emissions estimate from maritime traffic in the Italian harbours of Venice and Piombino. Sci. Total Environ. 1995, 169, 257–263. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset 1 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Dataset 2 | |||||||||
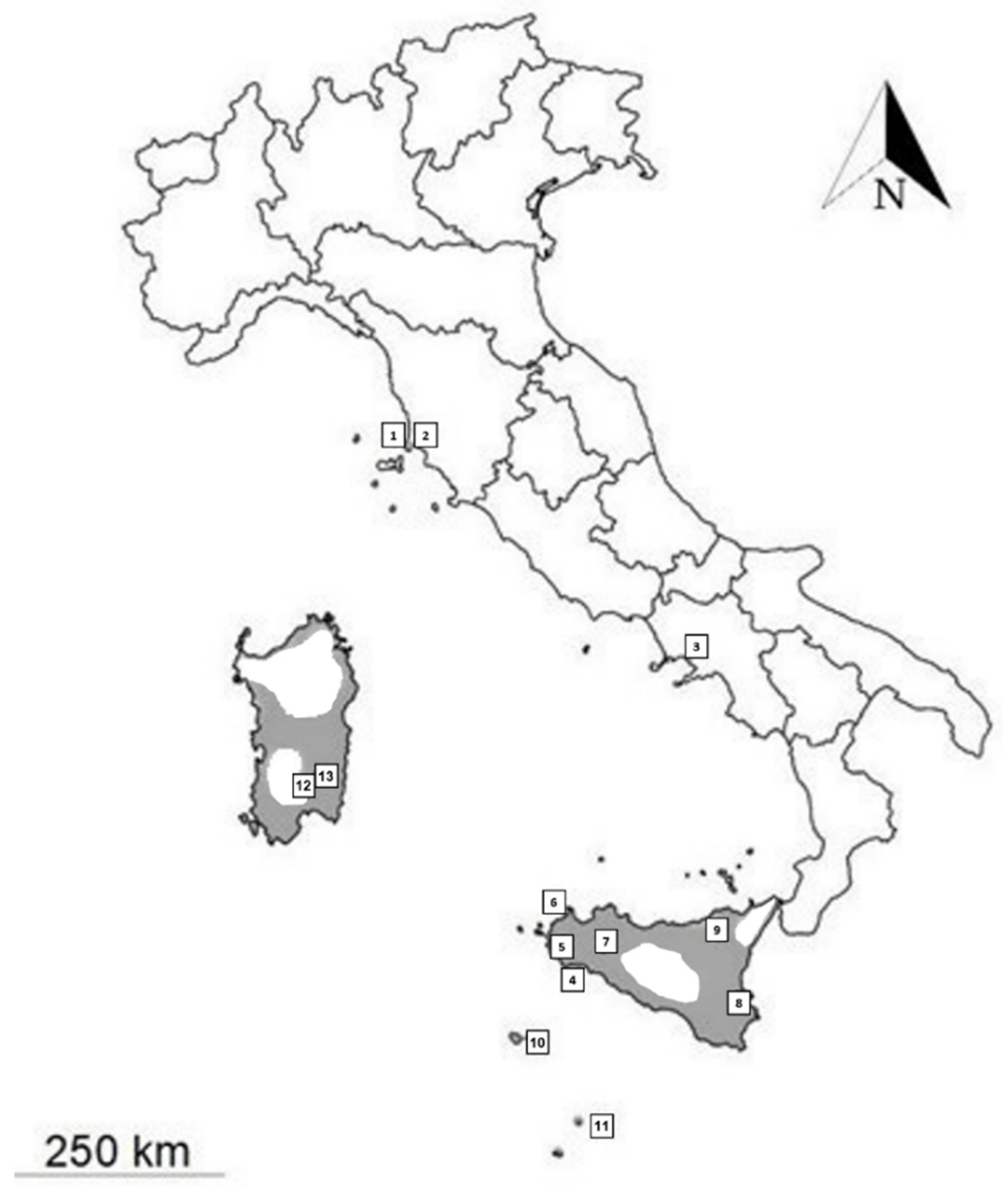
| Species | Region/District | Locality (Point in Figure 2) | Latitude | Longitude | Sample Label | Accession Numbers | COXI | 12S | CYTB |
| C. ocellatus | Saudi Arabia | Ash Shihyah | 26.269 | 43.597 | CH01_ARABIA | ON534012, ON534203, ON551370 | X | X | X |
| C. ocellatus | Tuscany/Livorno | Torre Mozza (2) | 42.946 | 10.693 | CH02_TORRE_MOZZA | ON534009, ON534204, ON551371 | X | X | X |
| C. ocellatus | Sicily/Trapani | Pantelleria island (10) | 36.817 | 12.003 | CH05_PANTELLERIA | ON534007, ON534202, ON551381 | X | X | X |
| C. ocellatus | Sicily/Trapani | Triscina di Selinunte (4) | 37.584 | 12.785 | CH06_SELINUNTE | ON534013, ON534206, ON551377 | X | X | X |
| C. ocellatus | Sicily/Trapani | Ficuzza (7) | 37.883 | 13.373 | CH07_FICUZZA | ON534014,ON534207, ON551375 | X | X | X |
| C. ocellatus | Sicily/Trapani | Mazara del Vallo (5) | 37.650 | 12.599 | CH08_MAZARA | ON534015, ON534208, ON551378 | X | X | X |
| C. ocellatus | Sicily/Messina | Rocche del Crasto (9) | 38.023 | 14.746 | CH09_MESSINA | ON534019, ON53420, ON551376 | X | X | X |
| C. ocellatus | Sicily/Trapani | Riserva dello Zingaro (6) | 38.081 | 12.808 | CH010_ZINGARO | ON534016, ON534209, ON551379 | X | X | X |
| C. ocellatus | Sardegna/Sud Sardegna | Dolianova (13) | 39.374 | 9.180 | CH011_SARDINIA | ON551373 | X | ||
| C. ocellatus | Tuscany/Livorno | Piombino (1) | 42.924 | 10.544 | CH015_PIOMBINO | ON534010, ON534205, ON551372 | X | X | X |
| C. ocellatus | Sicily/Agrigento | Linosa island (11) * | 35.870 | 12.862 | CH016_LINOSA | ON534017, ON534210, ON551382 | X | X | X |
| C. ocellatus | Campania/Naples | Portici (3) | 40.804 | 14.349 | CH017_PORTICI | ON534011, ON534211, ON551380 | X | X | X |
| C. ocellatus | Sicily/Siracusa | Siracusa (8) | 37.080 | 15.286 | CH018_SIRACUSA | ON534018, ON534212, ON551374 | X | X | X |
| C. ocellatus | Sardegna/Sud Sardegna | Serdiana (12) | 39.372 | 9.159 | CH019_SARDINIA | ON534008,ON534213, ON551383 | X | X | X |
| C. ocellatus | Algeria | EU278169_ALGERIA | EU278169 | X | |||||
| C. ocellatus | Morocco | EU278171_MOROCCO | EU278171 | X | |||||
| C. ocellatus | Turkey | EU278180_TURKEY | EU278180 | X | |||||
| C. ocellatus | Syria | FJ980143_SYRIA | FJ980143 | X | |||||
| C. ocellatus | Greece | FJ980268_GREECE | FJ980268 | X | |||||
| C. ocellatus | Egypt | EU278181_EGYPT | EU278181 | X | |||||
| C. ocellatus | Israel | EU278184_ISRAEL | EU278184 | X | |||||
| C. ocellatus | Tunisia | EU278194_TUNISIA | EU278194 | X | |||||
| C. ocellatus | Sardinia | EU278194_SARTIL | EU278186 | X | |||||
| C. ocellatus | Tunisia | EU278188_TUNTIL | EU278188 | X | |||||
| C. chalcides | Italy | Giglio island (Grosseto) | EU278211_CCGIGLIO | EU278211 | X | ||||
| C. chalcides | Italy | Piombino (Livorno) | EU278212_CCPIOMBINO | EU278212 | X | ||||
| C. viridanus | Spain | Canary Islands | EU278116_CVCANARY | EU278211 | X | ||||
| Target Gene | Label | Sequence 5′-3′ | Reference | Fragment Length (bp) |
|---|---|---|---|---|
| cytb | L14841 | AAAAAGCTTCCATCCAACATCTCACATGATGAAA | [26] | 325 |
| H15149 | AAACTGCAGCCCCTCAGAATGATATTTGTCCTCA | |||
| 12S | 12StPhe | AAAGCACRGCACTGAAGATGC | [27] | 350 |
| 12 g | TATCGATTATAGGACAGGCTCCTCTA | [28] | ||
| COXI | LCO1490 | GGTCAACAAATCATAAAGATATTGG | [29] | 670 |
| HCO 2198 | TAAACTTCAGGGTGACCAAAAAATCA |
| Torre Mozza | Piombino | Serdiana | Siracusa | Ficuzza | Rocche Crasto | Triscina Selinunte | Mazara Vallo | Riserva Zingaro | Portici | Pantelleria | Linosa | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Torre Mozza | 0.5% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | |
| Piombino | 1.4% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | |
| Serdiana | 4.3% | 3.6% | 0% | 0% | 0% | 0% | 0% | 0% | 2% | 0% | 0% | |
| Siracusa | 2.5% | 3.5% | 1.4% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | |
| Ficuzza | 4% | 4% | 2.6% | 1.6% | 0% | 0% | 0% | 0% | 0% | 0% | 0% | |
| Rocche Crasto | 2.5% | 3.2% | 1.4% | 0.1% | 1.6% | 0% | 0% | 0% | 0% | 0% | 0% | |
| Triscina Selinunte | 3.2% | 2.5% | 2.1% | 0.1% | 0.5% | 1% | 0% | 0% | 0% | 0% | 0% | |
| Mazara Vallo | 2.3% | 4% | 1.2% | 0.2% | 1.4% | 1.4% | 2% | 0% | 0% | 0% | 0% | |
| Riserva Zingaro | 2% | 2% | 1.7% | 0.7% | 1% | 1% | 1% | 3% | 0% | 0% | 0% | |
| Portici | 4.3% | 3.6% | 0.2% | 1% | 2.6% | 1.8% | 2% | 4% | 1% | 0% | 0% | |
| Pantelleria | 2.5% | 2.5% | 1.7% | 1% | 1.6% | 0.7% | 2% | 0.5% | 1% | 0% | 0% | |
| Linosa | 2.5% | 2.5% | 1.2% | 0.2% | 0.9% | 1.4% | 1.4% | 1.5% | 1.3% | 2% | 2% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mori, E.; Andreone, F.; Viviano, A.; Faraone, F.P.; Di Nicola, M.R.; Borri, B.; Bruni, G.; Mazza, G.; Banchi, R.; Zaccaroni, M.; et al. Aliens Coming by Ships: Distribution and Origins of the Ocellated Skink Populations in Peninsular Italy. Animals 2022, 12, 1709. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131709
Mori E, Andreone F, Viviano A, Faraone FP, Di Nicola MR, Borri B, Bruni G, Mazza G, Banchi R, Zaccaroni M, et al. Aliens Coming by Ships: Distribution and Origins of the Ocellated Skink Populations in Peninsular Italy. Animals. 2022; 12(13):1709. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131709
Chicago/Turabian StyleMori, Emiliano, Franco Andreone, Andrea Viviano, Francesco Paolo Faraone, Matteo Riccardo Di Nicola, Bernardo Borri, Giacomo Bruni, Giuseppe Mazza, Riccardo Banchi, Marco Zaccaroni, and et al. 2022. "Aliens Coming by Ships: Distribution and Origins of the Ocellated Skink Populations in Peninsular Italy" Animals 12, no. 13: 1709. https://0-doi-org.brum.beds.ac.uk/10.3390/ani12131709