1. Introduction
Seaweeds have been exploited for several purposes, including feed, food and biofuel applications, due to their nutritive and bioactive compounds [
1]. In particular, green macroalgae, mainly the genus
Ulva, have high growth rates leading to rapid biomass accumulation [
2], which caused them to be increasingly produced worldwide [
3]. Although there are environmental and economic impacts of seaweed production, the use of integrated multi-trophic aquaculture (IMTA) offers a more sustainable production system for
Ulva sp. [
4]. The nutritional composition of
Ulva sp. is variable but, in general, these algae are good sources of chlorophylls
a and
b, carotenoids and minerals, such as iodine [
1]. Although the protein content is, on average, 15.8% dry matter (DM), for some species, such as
U. lactuca and
U. prolifera, crude protein can reach values of 31.6 to 41.8% DM [
5]. Total lipids are found in low amounts (up to 6.6% DM) in green seaweeds but with a healthy profile of beneficial polyunsaturated fatty acids (PUFA) [
6]. The bioactive constituents of
Ulva sp. are mainly antioxidant pigments [
7] and the sulphated polysaccharide ulvan. This polymer has many human health-promoting functions, including immunomodulation and antioxidant activity [
8].
Considering the benefits of using
Ulva sp. as a feed ingredient, several studies reported their impact on poultry growth or meat quality [
9,
10,
11,
12,
13,
14,
15]. However, only a few assessed the effect of incorporating over 10% of
Ulva sp. in feeds [
9,
10]. Ventura et al. [
10] observed no negative effect on broilers’ growth of feeding 10% of
Ulva rigida, but higher algae levels compromised animal growth. In fact, high inclusion levels of green seaweeds may impair nutrient digestibility as a result of indigestible algae cell wall polysaccharides [
16], which mainly comprise gel-forming ulvan and insoluble cellulose, together with low amounts of xyloglucan and glucuronans [
17]. Therefore, the use of Carbohydrate-Active enZymes (CAZymes) emerges as a possible solution to degrade the
Ulva sp. cell wall, due to their efficiency in hydrolysing
Ulva sp. biomass for protein and carbohydrate extractions [
18,
19,
20]. In addition, the in vitro ability of a single ulvan lyase from the family of 25 polysaccharide lyases (PL25) to partially disrupt the
U. lactuca cell wall and release mono- and oligosaccharides, and monounsaturated fatty acids (e.g., C18:1
c9), was recently reported [
21]. Furthermore, CAZymes have shown activity towards carbohydrate present in microalgae cell walls [
22,
23] and in grains [
24,
25]. Thus, the degradation of seaweed biomass with feed enzymes would optimize their utilization as feedstuffs to partially replace unsustainable and conventional sources such as maize and soybean meal [
1]. However, to the best of our knowledge, the effect of feeding macroalgae supplemented with CAZymes on broiler performance and meat quality was only evaluated in a few studies using
Ulva sp. [
26,
27] or a brown seaweed,
Laminaria digitata [
28].
In the present study, we hypothesized that the combination of a previously in vitro tested ulvan lyase (PL25) [
21], or a commercial carbohydrase mixture with dietary
U. lactuca replacing 15% of maize and soybean meal, increases the nutritive value of seaweed for broilers with a consequent improvement of meat quality through the deposition of health promoting bioactive compounds, without compromising animal growth performance.
4. Discussion
The incorporation of 15%
U. lactuca in broilers´ diet to partially replace maize and soybean meal did not significantly affect chicken growth performance, although a reduction in final body weight and ADG was found with the dietary supplementation of ulvan lyase, as well as a tendency for that with the other macroalgae treatments, in comparison with broilers fed the control diet. The negative effect of the recombinant CAZyme on body weight had no repercussion on FCR, and, therefore, feed enzymes had only a residual influence on animal performance. However, these results must be interpreted with caution, since an increased sample size or the measurement of performance parameters during a higher experimental period could give different outcomes with a possible decrease in broilers´ growth with
U. lactuca. Controversial results about the influence of feeding broilers with
Ulva sp. on animal growth are described in the literature. Indeed, Abudabos et al. [
11], Cañedo-Castro et al. [
13], and Ventura et al. [
10] reported no effect on ADG and FCR when feeding 3%, 6% or 10% of
Ulva sp. to broiler chicks, but Matshogo et al. [
14], Alagan et al. [
12], and Nhlane et al. [
15] reported an increase in FCR in broilers or hens fed up to 3–3.5% of
Ulva sp. These differences between studies are mostly due to different alga species and growing stage of birds [
1], which can modify the way animals cope with the presence of indigestible polysaccharides in algal biomass and their influence on nutrient digestibility [
16]. However, few studies evaluated the effect of dietary
Ulva sp. on chicken growth when incorporated at levels higher than 10% feed [
9], which is due to the fact that 20 and 30% of seaweed were reported to reduce ADG and increase FCR in broilers [
10]. In the present study, the ulvan lyase supplementation was expected to increase the bioaccessibility of nutrients, which indeed occurred for certain compounds, as, for instance, PUFA. However, this phenomenon was not enough to overcome the slight reduction in ADG in broilers fed 15% of
U. lactuca, which might have been exacerbated by a decrease of 0.14% (as fed) of the estimated available proportion of the essential amino acid lysine in macroalga-containing diets relative to the control diet (
Table 1).
Moreover, dietary
U. lactuca had no effect on the weight and length of most of the gastrointestinal organs analysed, except for a reduction in liver weight and, when combined with ulvan lyase, an increase in ileum length. The impact of
U. lactuca on liver was unexpected, since previous studies on broilers [
14] or hens [
9,
15] fed either 2–3.5% [
14,
15] or 5–25% [
9] of
Ulva sp. reported no effects of seaweed on this organ. However, the present results are possibly caused by an increased content of algal indigestible polysaccharides, such as cellulose, in the broiler´s gut. Indeed, these compounds would decrease nutrient availability and, thus, liver metabolism with a consequent reduction on the organ size [
38]. This effect of
U. lactuca can further explain the decrease in total lipids found in the muscle of broilers fed macroalgae diets, since the liver plays a main role in lipogenesis [
38]. The increase in ileum length with
U. lactuca combined with the recombinant CAZyme might have been due to the bioactivity of algal polysaccharides, namely, ulvan, towards enterocytes proliferation and intestinal microbiota. This functional activity of polysaccharides was suggested by Cañedo-Castro et al. [
13] and Guo et al. [
39] after finding an increase in intestinal villus height in broilers and laying hens caused by 2 to 6% of
Ulva rigida- and 0.5% of
Ulva sp.-extracted polysaccharides in the diet, respectively.
The combination of
U. lactuca and ulvan lyase reduced the ileal viscosity found with
U. lactuca alone or in combination with the commercial CAZyme mixture, even though ulvan lyase did not completely reverse the viscosity level to that observed with the control diet. A similar effect was recently reported when broilers were fed 15% of
L. digitata combined with a recombinant alginate lyase [
28]. The present results may indicate that the ulvan lyase partially disrupted the ulvan gel-forming structure composing the
U. lactuca cell wall, as previously observed in vitro [
21], with a consequent reduction in digesta viscosity caused by the presence of seaweed. However, this activity of ulvan lyase was not reflected in an improvement of broilers´ growth performance, which suggests a minor effect of the recombinant CAZyme on feed passage and nutrient digestibility [
40].
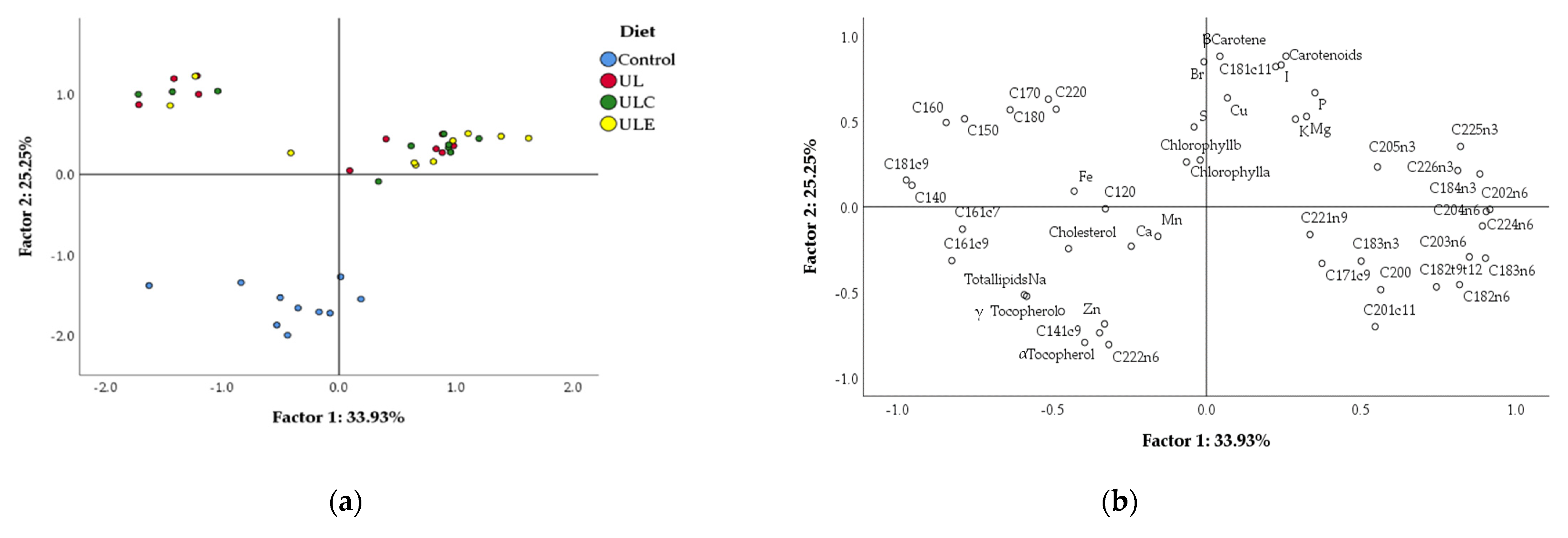
Considering meat quality traits, the breast colour was strongly influenced by dietary
U. lactuca, with a 4-fold increase in b* value and decreases of L* and a* values. These results led to a visible yellowish meat colour for all macroalga-containing treatments. Conversely, the few studies reporting the influence of
Ulva sp. On breast meat colour of broilers showed either no effect of 1–3% of
U. lactuca [
11] or only a time × algae dose (2–3.5%
Ulva sp.) interaction leading to an increase in a* and L* values without any effect on b* values [
14]. These differences are probably due to distinct seaweed levels in the diet, as previously reported with the microalga
Arthrospira platensis. For instance, 15% of
A. platensis fed to broilers increased the b* value of meat [
29], but no effect was observed when feeding a lower alga dose (0.25 to 1%) [
41]. To the best of our knowledge, the present study is the first to evaluate the impact on meat colour of feeding up to 15% of
Ulva to broilers. Such high dose of macroalga promoted a significant increase on the accumulation of carotenoids in meat, which is associated with the development of yellow colour, as previously demonstrated in broilers fed up to 8% [
42] and 15% [
29] of
Arthrospira sp., or in fish fed 10 or 15% of
U. rigida [
43,
44]. Indeed, the zeaxanthin pigment was positively correlated with the presence of yellow colour in the breast muscle of broilers [
42]. A recent study in broilers fed 15% of
L. digitata also demonstrated an increase in carotenoids in meat caused by the presence of algae, but with a milder effect on breast colour [
28].
Ulva sp. are generally rich in several carotenoids, including β- and α-carotenes and xanthophylls, such as astaxanthin, zeaxanthin and lutein [
7,
45]. These compounds have numerous health benefits acting as precursors of vitamin A and retina protectors through its accumulation in eye macula and also as antioxidant and anti-inflammatory agents [
7,
46]. Therefore, in the present study, the meat nutritional value of broilers was enhanced by the 8.9-fold increase in carotenoids in the breast.
Despite that, the yellow–orange colour of meat conferred by the pigments can lead to a negative perception of meat by consumers, particularly in Europe [
47,
48], although in some countries, such as Mexico, and, more variably and less markedly, in the United States, consumers tend to prefer a more pigmented broiler skin [
49,
50]. Moreover, it is worth mentioning that, at the end of the 20th century, the yellow colour of meat was generally desirable, since it was an indicator of healthier meat [
47]. In current times, the search for healthy food might again stimulate the preference for a yellowish meat colour, even though no studies can confirm this aspect. In the present study, it seems that the reduction in meat acceptability with macroalga treatments was caused by a decrease in juiciness and tenderness and the presence of off-flavours (metallic, herb-like or fishy flavours) rather than by the yellowish colour of meat, since the sensory panel did not indicate the colour as a factor that compromised meat overall acceptability. Additionally, the meat was still positively scored (>4.0) for all treatments. However, a previous study [
51] showed that the meat colour significantly influences the perception of consumers towards its tenderness and flavour, and thus, this aspect should not be easily disregarded. Recently, a decrease in juiciness in broiler meat was reported with 15% of dietary
A. platensis [
29] or
L. digitata combined with a recombinant CAZyme [
28]. A negative effect on overall acceptability of fillet was also observed with 10 and 15% of
U. rigida [
44], which was attributed to the change in meat colour, since there was no impact of seaweed on other sensory attributes. However, the effect of
Ulva sp. on the sensory traits of poultry meat needs to be further explored.
The total lipids and fatty acid profile in the meat were influenced by dietary
U. lactuca, since there was a decrease in total lipid content with the sole use of macroalga and an increase in n-3 PUFA, mostly C20:5n-3, C22:5n-3 and C22:6n-3, with
U. lactuca combined with ulvan lyase. A similar impact of macroalga on total
n-3 PUFA in meat was reported when broilers [
52] or piglets [
52] were fed 15% or 10% of
L. digitata, respectively, although without an enzymatic effect. In spite of the present results, lean meats, with an average of 1.89% total lipids (<5%) [
53], were obtained for all treatments. The increase in n-3 PUFA caused by combining
U. lactuca with ulvan lyase indicates a degradation of alga with consequent release of nutritional and bioactive compounds. An effect of the recombinant CAZyme on releasing fatty acids from
U. lactuca biomass was recently demonstrated in vitro, but only some MUFA, particularly C18:1
c9, were significantly released [
21]. The enrichment of poultry meat with n-3 long-chain PUFA can have various human health benefits, such as the diminishment of atherosclerosis and cardiovascular diseases [
54,
55]. In the present study, the 2-fold decrease in n-6/n-3 PUFA ratio in the meat from macroalga treatments relative to control could indicate a healthier meat, according to Wood
, et al. [
56], although the ratio was much higher than the recommended value (<4.0).
Furthermore, the mineral profile of meat was markedly affected by the incorporation of U. lactuca in broiler diet, with an increase in total macrominerals, including magnesium, phosphorus and potassium, accompanied by a decrease in sodium. Most of the microminerals were also influenced by dietary seaweed, since bromine, copper and iodine increased, and zinc decreased, with macroalga treatments.
The increased deposition of macrominerals in meat with
U. lactuca enhances its nutritional value, considering the health benefits of magnesium, phosphorus and potassium. Indeed, magnesium is an important component of bones and is involved in oxygen uptake, energy production and electrolyte balance [
57]; phosphorus participates in numerous biological processes, including ATP synthesis and bone mineralization [
58]; and potassium reduces blood pressure and the risk of cardiovascular diseases [
55,
59]. Although caution must be taken with the high phosphorus content normally present in Western diets, the concentration of phosphorus in the meat from macroalga treatments (average of 254.7 mg/100 g) was about half of that recommended by EFSA [
60] (550 mg/day) for an adult ingesting 100 g of meat per day. Moreover, the increase in potassium in meat with dietary
U. lactuca was due to a high amount of this mineral in the alga biomass (38.822 g/kg dry matter), which is within the range described for
Ulva sp. [
1]. This occurrence explains the 1.24-fold decrease in sodium/potassium ratio in meat with the macroalga treatments. A similar result was recently observed when feeding finishing pigs with 5% of the microalga
Chlorella vulgaris combined with a four-CAZyme mixture [
61]. The reduction in sodium/potassium ratio is desirable due to the imbalance of these minerals in Western diets. Additionally, a high ratio was previously described as a good predictor of high hypertension prevalence and cardiovascular diseases [
62].
The accumulation of copper and iodine in meat with the seaweed can either have positive effects on human health or cause toxicity for consumers [
63,
64], whereas bromine does not have recognized benefits and is considered a toxic element [
65]. Although copper is a cofactor of many oxido-reductive enzymatic processes [
64] and iodine plays a crucial role on the synthesis of thyroid hormone [
63], pathological conditions may occur if allowances for these minerals in the diet are exceeded. Nevertheless, in the present study, feeding broilers with 15% of
U. lactuca is not expected to have a major impact on meat nutritional quality in terms of micromineral content. Indeed, the dietary macroalga was responsible for a small increase in copper (0.068 to 0.085 mg/100 g), iodine (0.001 to 0.003 mg/100 g) and bromine (1.71 to 4.29 µg/kg body weight/day) in broiler meat leading to values much lower than the recommended daily intake for copper [
64] and iodine [
63] (0.90 mg and 0.15 mg /day, respectively) or the maximum allowance for bromine (1000 µg bromine/kg body weight/day) [
65], for an adult person (70 kg) consuming 100 g of meat per day.


 ,
, 



{kind=link}