
Combination of Transcriptomics and Proteomics Reveals Differentially Expressed Genes and Proteins in the Skin of EDAR Gene-Targeted and Wildtype Cashmere Goats
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Collection of Skin Samples from Cashmere Goats
2.3. RNA Sequencing (RNA-seq) Analysis
2.4. Proteomic Analysis
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Haematoxylin–Eosin (H&E) Staining
2.7. Statistical Analyses
3. Results
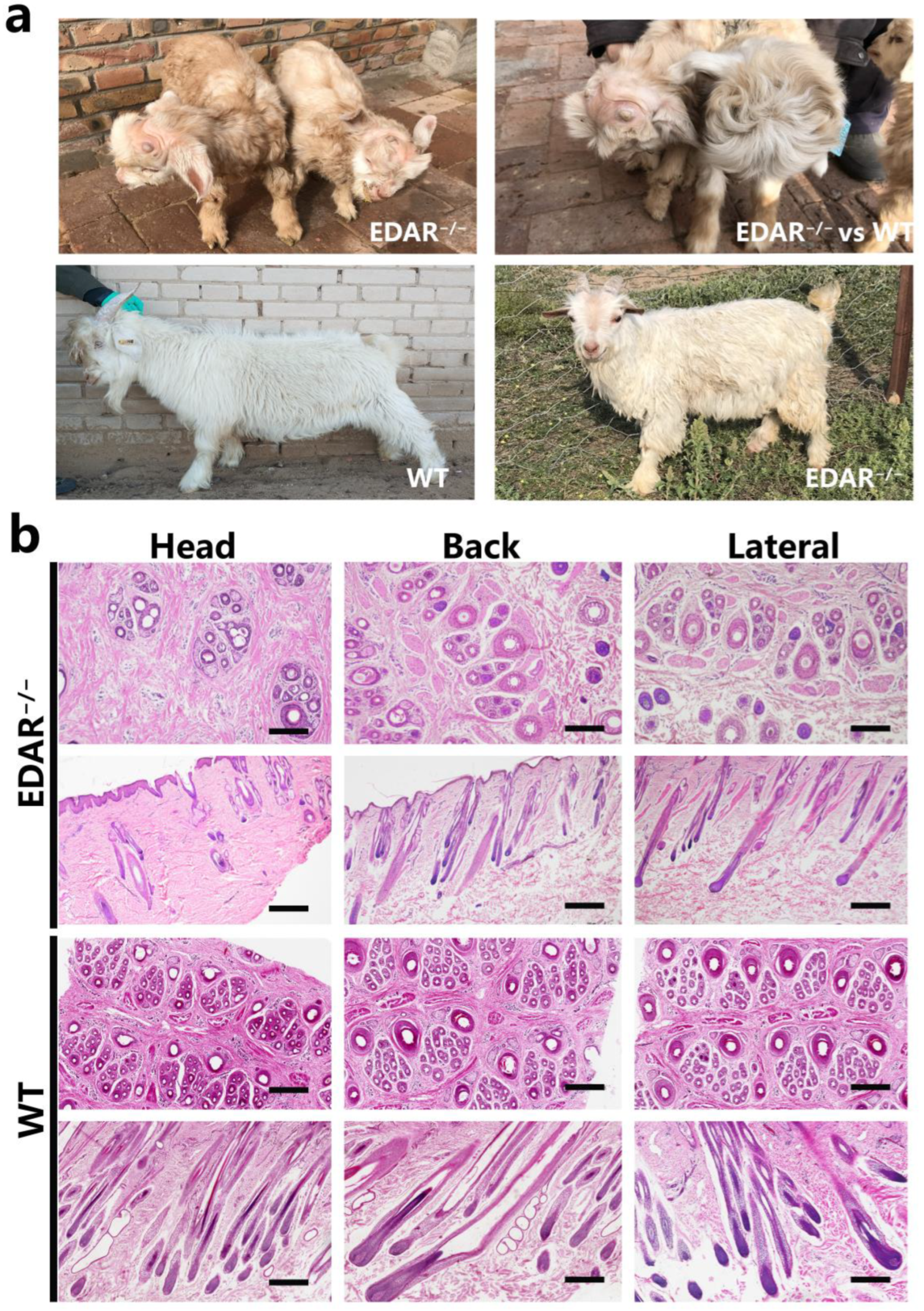
3.1. Phenotypes of EDAR Gene-Targeted Cashmere Goats
3.2. Analysis of Differences in Gene Expression
3.3. Analysis of Differences in Protein Expression
3.4. Enrichment Analysis of DEGs and DEPs in KEGG
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ge, W.; Zhang, W.; Zhang, Y.; Zheng, Y.; Li, F.; Wang, S.; Liu, J.; Tan, S.; Yan, Z.; Wang, L.; et al. A Single-cell Transcriptome Atlas of Cashmere Goat Hair Follicle Morphogenesis. Genom. Proteom. Bioinform. 2021, 19, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, K.; Wang, L.; Wang, Z.; Han, W.; Chen, D.; Wei, Y.; Su, R.; Wang, R.; Liu, Z.; et al. Comparative study on seasonal hair follicle cycling by analysis of the transcriptomes from cashmere and milk goats. Genomics 2020, 112, 332–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fang, X.; Zhao, Y. The occupied proportion of hair follicle group in Inner Mongolia cashmere goat (Capra hircus). Small Rumin. Res. 2022, 210, 106667. [Google Scholar] [CrossRef]
- Tseveenjav, B.; Garrick, D.J.; Batjargal, E.; Yondon, Z. Economic selection index to improve fiber quality in Mongolian Cashmere goats. Livest. Sci. 2020, 232, 103898. [Google Scholar] [CrossRef]
- Wang, H.-D.; Yang, L.; Yu, X.-J.; He, J.-P.; Fan, L.-H.; Dong, Y.-J.; Dong, C.-S.; Liu, T.-F. Immunolocalization of β-catenin and Lef-1 during postnatal hair follicle development in mice. Acta Histochem. 2012, 114, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Mady, L.J.; Ajibade, D.V.; Hsaio, C.; Teichert, A.; Fong, C.; Wang, Y.; Christakos, S.; Bikle, D.D. The Transient Role for Calcium and Vitamin D during the Developmental Hair Follicle Cycle. J. Investig. Dermatol. 2016, 136, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Benavides, F.; Starost, M.F.; Flores, M.; Gimenez-Conti, I.B.; Guénet, J.-L.; Conti, C.J. Impaired Hair Follicle Morphogenesis and Cycling with Abnormal Epidermal Differentiation in nackt Mice, a Cathepsin L-Deficient Mutation. Am. J. Pathol. 2002, 161, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhu, L.; He, J. Morphogenesis, Growth Cycle and Molecular Regulation of Hair Follicles. Front. Cell Dev. Biol. 2022, 10, 899095. [Google Scholar] [CrossRef]
- Yang, F.; Li, R.; Zhao, C.; Che, T.; Guo, J.; Xie, Y.; Wang, Z.; Li, J.; Liu, Z. Single-cell sequencing reveals the new existence form of dermal papilla cells in the hair follicle regeneration of cashmere goats. Genomics 2022, 114, 110316. [Google Scholar] [CrossRef]
- Yin, R.; Wang, Y.; Wang, Z.; Zhu, Y.; Cong, Y.; Wang, W.; Deng, L.; Liu, H.; Guo, D.; Bai, W. Discovery and molecular analysis of conserved circRNAs from cashmere goat reveal their integrated regulatory network and potential roles in secondary hair follicle. Electron. J. Biotechnol. 2019, 41, 37–47. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Li, X.; Han, W.; Yang, K.; Wang, H.; Zhang, Y.; Su, R.; Liu, Z.; Wang, R.; et al. High-throughput sequencing of hair follicle development-related micrornas in cashmere goat at various fetal periods. Saudi J. Biol. Sci. 2018, 25, 1494–1508. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Zhang, L.; Gao, K.; Guo, Y.; Liu, Y.; Zhang, Y. Cashmere production, skin characteristics, and mutated genes in crimped cashmere fibre goats. Animal 2022, 16, 100565. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yang, Z.; Andl, T.; Cui, C.; Kim, N.; Millar, S.E.; Cotsarelis, G. Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding. Nature 2007, 447, 316–320. [Google Scholar] [CrossRef]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Qureshi, W.M.S.; Filip, S.; Mokry, J. Signaling Involved in Hair Follicle Morphogenesis and Development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.B.; Cohen, I.; Kumar, V.; Xu, Z.; Bar, C.; Dauber-Decker, K.L.; Tsai, P.-C.; Marangoni, P.; Klein, O.D.; Hsu, Y.-C.; et al. FGF signalling controls the specification of hair placode-derived SOX9 positive progenitors to Merkel cells. Nat. Commun. 2018, 9, 2333. [Google Scholar] [CrossRef] [PubMed]
- van der Hout, A.H.; Oudesluijs, G.G.; Venema, A.; Verheij, J.B.G.M.; Mol, B.G.J.; Rump, P.; Brunner, H.G.; Vos, Y.J.; van Essen, A.J. Mutation screening of the Ectodysplasin-A receptor gene EDAR in hypohidrotic ectodermal dysplasia. Eur. J. Hum. Genet. 2008, 16, 673–679. [Google Scholar] [CrossRef]
- Fessing, M.Y.; Sharova, T.Y.; Sharov, A.A.; Atoyan, R.; Botchkarev, V.A. Involvement of the Edar signaling in the control of hair follicle involution (catagen). Am. J. Pathol. 2006, 169, 2075–2084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tomann, P.; Andl, T.; Gallant, N.M.; Huelsken, J.; Jerchow, B.; Birchmeier, W.; Paus, R.; Piccolo, S.; Mikkola, M.L.; et al. Reciprocal Requirements for EDA/EDAR/NF-κB and Wnt/β-Catenin Signaling Pathways in Hair Follicle Induction. Dev. Cell 2009, 17, 49–61. [Google Scholar] [CrossRef]
- Pispa, J.; Pummila, M.; Barker, P.A.; Thesleff, I.; Mikkola, M.L. Edar and Troy signalling pathways act redundantly to regulate initiation of hair follicle development. Hum. Mol. Genet. 2008, 17, 3380–3391. [Google Scholar] [CrossRef]
- Schmidt-Ullrich, R.; Tobin, D.J.; Lenhard, D.; Schneider, P.; Paus, R.; Scheidereit, C. NF-κB transmits Eda A1/EdaR signalling to activate Shh and cyclin D1 expression, and controls post-initiation hair placode down growth. Development 2006, 133, 1045–1057. [Google Scholar] [CrossRef]
- Fliniaux, I.; Mikkola, M.L.; Lefebvre, S.; Thesleff, I. Identification of dkk4 as a target of Eda-A1/Edar pathway reveals an unexpected role of ectodysplasin as inhibitor of Wnt signalling in ectodermal placodes. Dev. Biol. 2008, 320, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Yan, W.; Li, X.; Wang, H.; Wang, Y.; Hu, X.; Liu, X.; Liang, H.; Liu, D. Generation of Cashmere Goats Carrying an EDAR Gene Mutant Using CRISPR-Cas9-Mediated Genome Editing. Int. J. Biol. Sci. 2018, 14, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, T.; Zhang, Z.; Payne, S.H.; Zhang, B.; McDermott, J.E.; Zhou, J.-Y.; Petyuk, V.A.; Chen, L.; Ray, D.; et al. Integrated Proteogenomic Characterization of Human High-Grade Serous Ovarian Cancer. Cell 2016, 166, 755–765. [Google Scholar] [CrossRef]
- Luber, C.A.; Cox, J.; Lauterbach, H.; Fancke, B.; Selbach, M.; Tschopp, J.; Akira, S.; Wiegand, M.; Hochrein, H.; O’Keeffe, M.; et al. Quantitative Proteomics Reveals Subset-Specific Viral Recognition in Dendritic Cells. Immunity 2010, 32, 279–289. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chan, J.K.C. The Wonderful Colors of the Hematoxylin–Eosin Stain in Diagnostic Surgical Pathology. Int. J. Surg. Pathol. 2014, 22, 12–32. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, Y.; Han, W.; Yang, K.; Hai, E.; Ma, R.; Di, Z.; Shang, F.; Su, R.; Wang, R.; et al. EDA and EDAR expression at different stages of hair follicle development in cashmere goats and effects on expression of related genes. Arch. Anim. Breed. 2020, 63, 461–470. [Google Scholar] [CrossRef]
- Adkins, D.E.; Khachane, A.N.; McClay, J.L.; Åberg, K.; Bukszár, J.; Sullivan, P.F.; van den Oord, E.J.C.G. SNP-based analysis of neuroactive ligand–receptor interaction pathways implicates PGE2 as a novel mediator of antipsychotic treatment response: Data from the CATIE study. Schizophr. Res. 2012, 135, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cheng, J.; Zhang, C.; Li, X. Cardioprotection Effects of Sevoflurane by Regulating the Pathway of Neuroactive Ligand-Receptor Interaction in Patients Undergoing Coronary Artery Bypass Graft Surgery. Comput. Math. Methods Med. 2017, 2017, 3618213. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.-Y.; Pi, L.-Q.; Hwang, S.T.; Lee, W.-S. Effect of IGF-I on Hair Growth Is Related to the Anti-Apoptotic Effect of IGF-I and Up-Regulation of PDGF-A and PDGF-B. Ann. Derm. 2012, 24, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Rudman, S.M.; Philpott, M.P.; Thomas, G.A.; Kealey, T. The Role of IGF-I in Human Skin and its Appendages: Morphogen as Well as Mitogen? J. Investig. Dermatol. 1997, 109, 770–777. [Google Scholar] [CrossRef]
- Perlmann, T. Retinoid metabolism: A balancing act. Nat. Genet. 2002, 31, 7–8. [Google Scholar] [CrossRef]
- Marceau, G.; Gallot, D.; Borel, V.; Lémery, D.; Dastugue, B.; Dechelotte, P.; Sapin, V. Molecular and metabolic retinoid pathways in human amniotic membranes. Biochem. Biophys. Res. Commun. 2006, 346, 1207–1216. [Google Scholar] [CrossRef]
- Gupta, S.; Masand, P.S. Citalopram and Hair Loss. Prim. Care Companion J. Clin. Psychiatry 2000, 2, 61–62. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Library | Total Reads | Mapped Reads | Mapping Rate |
|---|---|---|---|
| EDAR01 | 47,835,374 | 42,692,760 | 0.8925 |
| EDAR02 | 44,430,710 | 40,183,265 | 0.9044 |
| EDAR03 | 46,326,914 | 41,132,985 | 0.8879 |
| WT01 | 47,459,130 | 41,738,567 | 0.8795 |
| WT02 | 45,578,664 | 40,875,932 | 0.8968 |
| WT03 | 45,630,714 | 40,838,500 | 0.895 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, G.-Z.; Hao, F.; Zhu, L.; Jiang, G.-Q.; Yan, W.; Liu, J.; Liu, D.-J. Combination of Transcriptomics and Proteomics Reveals Differentially Expressed Genes and Proteins in the Skin of EDAR Gene-Targeted and Wildtype Cashmere Goats. Animals 2023, 13, 1452. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13091452
Gao G-Z, Hao F, Zhu L, Jiang G-Q, Yan W, Liu J, Liu D-J. Combination of Transcriptomics and Proteomics Reveals Differentially Expressed Genes and Proteins in the Skin of EDAR Gene-Targeted and Wildtype Cashmere Goats. Animals. 2023; 13(9):1452. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13091452
Chicago/Turabian StyleGao, Gui-Zhen, Fei Hao, Lei Zhu, Guo-Qing Jiang, Wei Yan, Jie Liu, and Dong-Jun Liu. 2023. "Combination of Transcriptomics and Proteomics Reveals Differentially Expressed Genes and Proteins in the Skin of EDAR Gene-Targeted and Wildtype Cashmere Goats" Animals 13, no. 9: 1452. https://0-doi-org.brum.beds.ac.uk/10.3390/ani13091452