
Comparative Analysis of the Growth, Physiological Responses, and Gene Expression of Chinese Soft-Shelled Turtles Cultured in Different Modes
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rice Planting and Turtle Rearing
2.2. Measurement and Sampling
- Specific growth rate (SGR, %/d) = 100 × (lnFBW − lnIBW)/days;
- Hepatosomatic index (HSI, %) = 100 × liver weight/body weight;
- Gonadosomatic index (GSI, %) = 100 × gonad weight/body weight;
- Intestinal somatic index (ISI, %) =100 × intestinal weight/body weight;
- Mesenteric fat index (MFI, %) = 100 × mesenteric fat weight/body weight;
- Limb fat index (LFI, %) = 100 × limb fat weight/body weight.
2.3. Physiological and Biochemical Parameter Analysis
2.4. RNA Extraction and Transcriptome Sequencing Analysis
2.5. Gene Expression Validation by qPCR
2.6. Statistical Analysis
3. Results
3.1. Growth Performance
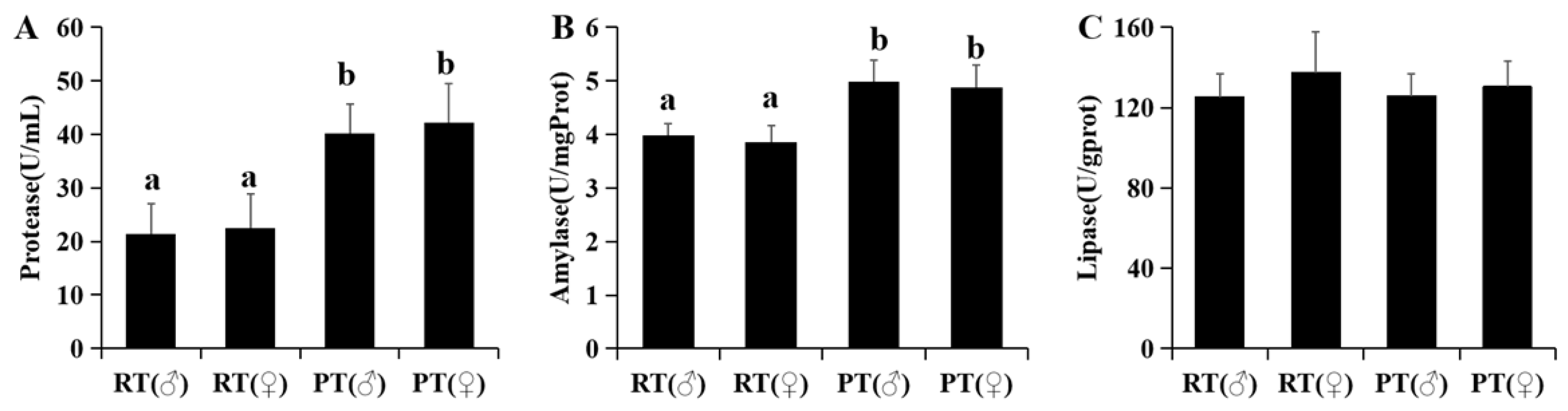
3.2. Physiological and Immune Indicators
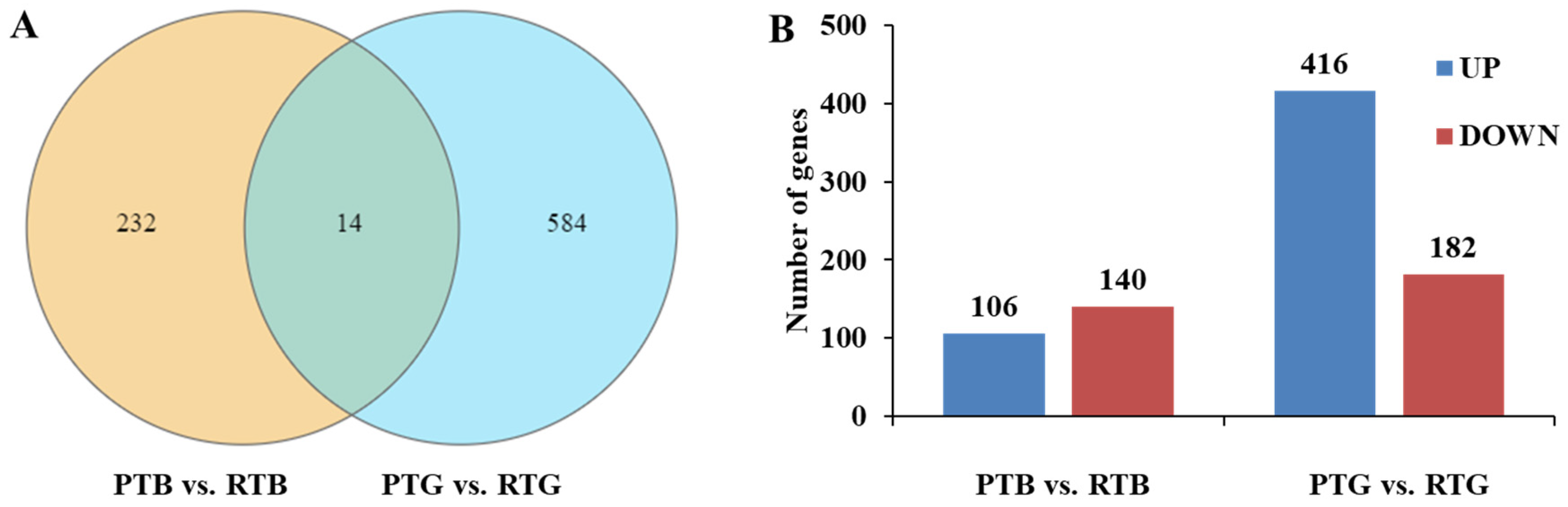
3.3. RNA-Seq Data and Gene Expression Profiles
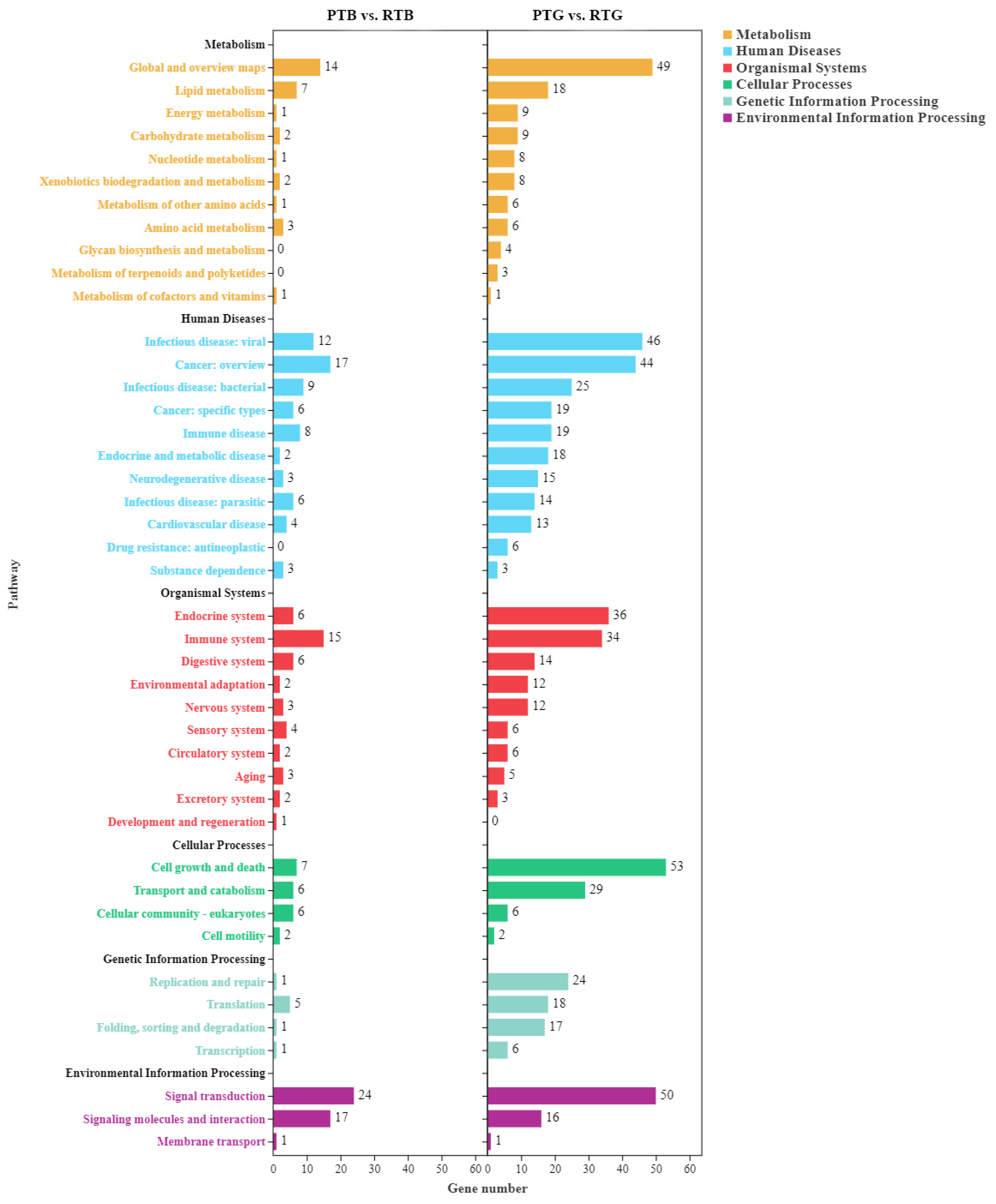
3.4. Functional Classification of DEGs
3.5. Validation of DEGs by qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beale, D.J.; Hillyer, K.; Nilsson, S.; Limpus, D.; Bose, U.; Broadbent, J.A.; Vardy, S. Bioaccumulation and metabolic response of PFAS mixtures in wild-caught freshwater turtles (Emydura macquarii macquarii) using omics-based ecosurveillance techniques. Sci. Total Environ. 2022, 806, 151264. [Google Scholar] [CrossRef] [PubMed]
- Castro Cortés, A.A.; Brieva, C.; Witte, C. Implications of wildlife trafficking on the health and conservation efforts of an endangered turtle species in Colombia. Conserv. Sci. Pract. 2022, 4, e595. [Google Scholar] [CrossRef]
- Wang, F.T.; Lai, N.Y.; Cheng, H.F.; Wu, H.R.; Liang, F.; Ye, T.; Lin, L.; Jiang, S.T.; Lu, J.F. Comparative analysis of the nutritional quality and volatile flavor constituents in the muscle of Chinese soft-shelled turtle from three different environments. Food Ferment. Ind. 2019, 45, 253–261. [Google Scholar]
- Chen, X.R.; Niu, C.J.; Pu, L.J. Effects of stocking density on growth and non-specific immune responses in juvenile soft-shelled turtle, Pelodiscus sinensis. Aquac. Res. 2007, 38, 1380–1386. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, F.; Jiang, Y.L.; Hou, G.J.; Cheng, Y.S.; Chen, H.L.; Li, X. Modern greenhouse culture of juvenile soft-shelled turtle, Pelodiscus sinensis. Aquac. Int. 2017, 25, 1607–1624. [Google Scholar] [CrossRef]
- Zhang, Y.; Fang, X.W.; Chen, X.Y.; Wang, H.K.; Liu, J.; Liang, X.Y.; Gu, Y.F.; Fang, C.; Yang, Y.Y. Bacillus cereus causes fatal disease in soft-shelled turtle (Trionyx sinensis). Aquaculture 2022, 547, 737473. [Google Scholar] [CrossRef]
- Zhou, X.W.; Tian, Y.; Liu, L.; Ge, L.R.; Zhang, H.; Liang, Y.X.; Wang, X.Q.; Hu, Y.Z. Pathologic, transcriptomic and microbiomic insight into the pathogenesis of intestinal parasitic tapeworm in cultured Chinese soft-shelled turtle (Pelodiscus sinensis). Aquaculture 2023, 562, 738788. [Google Scholar] [CrossRef]
- Arena, P.C.; Warwick, C.; Steedman, C. Welfare and environmental implications of farmed sea turtles. J. Agric. Environ. Ethics 2014, 27, 309–330. [Google Scholar] [CrossRef]
- Li, W.; Ding, H.Y.; Zhang, F.Y.; Zhang, T.L.; Liu, J.H.; Li, Z.J. Effects of water spinach Ipomoea aquatica cultivation on water quality and performance of Chinese soft-shelled turtle Pelodiscus sinensis pond culture. Aquacult. Environ. Interact. 2016, 8, 567–574. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Smith, S.A. Fish welfare in public aquariums and zoological collections. Animals 2023, 13, 2548. [Google Scholar] [CrossRef] [PubMed]
- Case, B.C.; Lewbart, G.A.; Doerr, P.D. The physiological and behavioural impacts of and preference for an enriched environment in the eastern box turtle (Terrapene carolina carolina). Appl. Anim. Behav. Sci. 2005, 92, 353–365. [Google Scholar] [CrossRef]
- Diggins, R.; Burrie, R.; Ariel, E.; Ridley, J.; Olsen, J.; Schultz, S.; Pettett-Willmett, A.; Hemming, G.; Lloyd, J. A review of welfare indicators for sea turtles undergoing rehabilitation, with emphasis on environmental enrichment. Anim. Welf. 2022, 31, 219–230. [Google Scholar] [CrossRef]
- Janča, M.; Gvoždík, L. Costly neighbours: Heterospecific competitive interactions increase metabolic rates in dominant species. Sci. Rep. 2017, 7, 5177. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.C.; Kyritsis, N.; Mills, M.; Godfrey, M.H.; Harms, C.A.; Anderson, P.E.; Shedlock, A.M. Tissue and temperature-specific RNA-seq analysis reveals genomic versatility and adaptive potential in wild sea turtle hatchlings (Caretta caretta). Animals 2021, 11, 3013. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Jin, F.; Xu, J.W.; Zhang, S.K.; Chen, D.X.; Hu, B.J.; Hong, Y.J. The impact of aquaculture system on the microbiome and gut metabolome of juvenile Chinese softshell turtle (Pelodiscus sinensis). IMeta 2022, 1, e17. [Google Scholar] [CrossRef]
- Sun, C.X.; Zhang, D.D.; Liu, W.B.; Cai, W.C.; Qian, Y.; Wang, K.Z.; Li, X.F.; Jiang, G.Z.; Xu, W.N. Growth performance, digestion and metabolism to fish meal replacement by rice protein concentrate in Chinese soft-shelled turtle Pelodiscus sinensis. Aquaculture 2018, 492, 321–326. [Google Scholar] [CrossRef]
- Wang, J.; Qi, Z.H.; Yang, Z.C. Evaluation of the protein requirement of juvenile Chinese soft-shelled turtle (Pelodiscus sinensis, Wiegmann) fed with practical diets. Aquaculture 2014, 433, 252–255. [Google Scholar] [CrossRef]
- Wang, Y.B.; Luo, X.Z.; Qu, C.J.; Xu, T.; Zou, G.W.; Liang, H.W. The important role of sex-related Sox family genes in the sex reversal of the Chinese soft-shelled turtle (Pelodiscus sinensis). Biology 2022, 11, 83. [Google Scholar] [CrossRef]
- Wan, N.F.; Li, S.X.; Li, T.; Cavalieri, A.; Weiner, J.; Zheng, X.Q.; Ji, X.Y.; Zhang, J.Q.; Zhang, H.L.; Zhang, H.; et al. Ecological intensification of rice production through rice-fish co-culture. J. Clean. Prod. 2019, 234, 1002–1012. [Google Scholar] [CrossRef]
- Yu, H.Y.; Zhang, X.C.; Shen, W.Y.; Yao, H.Y.; Meng, X.T.; Zeng, J.Y.; Zhang, G.B.; Zamanien, K. A meta-analysis of ecological functions and economic benefits of co-culture models in paddy fields. Agric. Ecosyst. Environ. 2023, 341, 108195. [Google Scholar] [CrossRef]
- Ge, L.; Sun, Y.; Li, Y.J.; Wang, L.Y.; Guo, G.Q.; Song, L.L.; Wang, C.; Wu, G.G.; Zang, X.Y.; Cai, X.M.; et al. Ecosystem sustainability of rice and aquatic animal co-culture systems and a synthesis of its underlying mechanisms. Sci. Total Environ. 2023, 880, 163314. [Google Scholar] [CrossRef]
- Xie, J.; Hu, L.L.; Tang, J.J.; Wu, X.; Li, N.N.; Yuan, Y.G.; Yang, H.S.; Zhang, J.E.; Luo, S.M.; Chen, X. Ecological mechanisms underlying the sustainability of the agricultural heritage rice–fish coculture system. Proc. Natl. Acad. Sci. USA 2011, 108, E1381–E1387. [Google Scholar] [CrossRef]
- Liu, G.B.; Huang, H.; Zhou, J.W. Energy analysis and economic assessment of a rice-turtle-fish co-culture system. Agroecol. Sustain. Food 2019, 43, 299–309. [Google Scholar] [CrossRef]
- Wu, B.L.; Chen, J.; Huang, L.; Zhang, Y.; Fang, T.; He, J.X. Dynamics of soil fertility and microbial community response to stocking density in rice-turtle co-culture. Aquacult. Rep. 2021, 20, 100765. [Google Scholar] [CrossRef]
- Wu, B.L.; Huang, L.; Chen, J.; Zhang, Y.; Wang, J.; He, J.X. Gut microbiota of homologous Chinese soft-shell turtles (Pelodiscus sinensis) in different habitats. BMC Microbiol. 2021, 21, 142. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Zhao, L.F.; Ye, J.; Ji, Z.J.; Tang, J.J.; Bai, K.; Zheng, S.J.; Hu, L.L.; Chen, X. Using aquatic animals as partners to increase yield and maintain soil nitrogen in the paddy ecosystems. Elife 2022, 11, e73869. [Google Scholar] [CrossRef]
- Ahmed, N.; Thompson, S.; Turchini, G.M. Organic aquaculture productivity, environmental sustainability, and food security: Insights from organic agriculture. Food. Sec. 2020, 12, 1253–1267. [Google Scholar] [CrossRef]
- Salmon, M.; Scholl, J. Allometric growth in juvenile marine turtles: Possible role as an antipredator adaptation. Zoology 2014, 117, 131–138. [Google Scholar] [CrossRef]
- Pontzer, H.; Raichlen, D.A.; Shumaker, R.W.; Ocbock, C.; Wich, S.A. Metabolic adaptation for low energy throughput in orangutans. Proc. Natl. Acad. Sci. USA 2010, 107, 14048–14052. [Google Scholar] [CrossRef]
- Rosenfeld, J.; Van Leeuwen, T.; Richards, J.; Allen, D. Relationship between growth and standard metabolic rate: Measurement artefacts and implications for habitat use and life-history adaptation in salmonids. J. Anim. Ecol. 2015, 84, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.Y.; Yao, S.B.; Shi, Y.; Cai, C.F.; Wang, C.R.; Wu, P.; Cao, X.M.; Ye, Y.T. Effects of low or high dosages of dietary sodium butyrate on the growth and health of the liver and intestine of largemouth bass, Micropterus salmoides. Aquacult. Nutr. 2022, 2022, 6173245. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.L.; Li, D.Y.; Zhang, J.M.; He, M.N.; Li, Y.; Liu, R.; Li, M.Z. Transcriptomic and lipidomic profiling of subcutaneous and visceral adipose tissues in 15 vertebrates. Sci. Data 2023, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Pond, C.M. Morphological aspects and the ecological and mechanical consequences of fat deposition in wild vertebrates. Annu. Rev. Ecol. Evol. Syst. 1978, 9, 519–570. [Google Scholar] [CrossRef]
- Snively, E.; O’Brien, H.; Henderson, D.M.; Mallison, H.; Surring, L.A.; Burns, M.E.; Holtz, T.R., Jr.; Russell, A.P.; Witmer, L.M.; Currie, P.J.; et al. Lower rotational inertia and larger leg muscles indicate more rapid turns in tyrannosaurids than in other large theropods. PeerJ 2019, 7, e6432. [Google Scholar] [CrossRef] [PubMed]
- Agha, M.; Ennen, J.R.; Bower, D.S.; Nowakowski, A.J.; Sweat, S.C.; Todd, B.D. Salinity tolerances and use of saline environments by freshwater turtles: Implications of sea level rise. Biol. Rev. 2018, 93, 1634–1648. [Google Scholar] [CrossRef] [PubMed]
- Vistro, W.A.; Zhang, Y.; Azhar, M.; Wu, R.Z.; Yang, S.; Chen, C.; Ding, B.T.; Qu, W.J.; Bai, X.B.; Huang, Y.F.; et al. Hematological and plasma biochemical parameters of Chinese soft-shelled turtle during hibernation and non-hibernation. Int. J. Agric. Biol. 2020, 23, 529–533. [Google Scholar]
- Deem, S.L.; Norton, T.M.; Mitchell, M.; Segars, A.L.; Alleman, A.R.; Cray, C.; Poppenga, R.H.; Dodd, M.; Karesh, W.B. Comparison of blood values in foraging, nesting, and stranded loggerhead turtles (Caretta caretta) along the coast of Georgia, USA. J. Wildl. Dis. 2009, 45, 41–56. [Google Scholar] [CrossRef]
- Qiu, Z.S.; Xu, Q.Y.; Xie, D.Z.; Zhao, J.T.; Yamamoto, F.Y.; Xu, H.; Zhao, J.H. Effects of the replacement of dietary fish meal with poultry by-product meal on growth and intestinal health of Chinese soft-shelled turtle (Pelodiscus sinensis). Animals 2023, 13, 865. [Google Scholar] [CrossRef]
- Dickinson, V.M.; Jarchow, J.L.; Trueblood, M.H. Hematology and plasma biochemistry reference range values for free-ranging desert tortoises in Arizona. J. Wildl. Dis. 2002, 38, 143–153. [Google Scholar] [CrossRef]
- Banaee, M.; Gholamhosseini, A.; Sureda, A.; Soltanian, S.; Fereidouni, M.S.; Ibrahim, A.T.A. Effects of microplastic exposure on the blood biochemical parameters in the pond turtle (Emys orbicularis). Environ. Sci. Pollut. Res. 2021, 28, 9221–9234. [Google Scholar] [CrossRef] [PubMed]
- Stokes, T.; Hector, A.J.; Morton, R.W.; Mcglory, C.; Phillips, S.M. Recent perspectives regarding the role of dietary protein for the promotion of muscle hypertrophy with resistance exercise training. Nutrients 2018, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Y.; Niu, C.J.; Chen, B.J.; Storey, K.B. Digital gene expression profiling reveals transcriptional responses to acute cold stress in Chinese soft-shelled turtle Pelodiscus sinensis juveniles. Cryobiology 2018, 81, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Zhang, Z.; Guan, Y.Q. Physiological and transcriptional analysis of Chinese soft-shelled turtle (Pelodiscus sinensis) in response to acute nitrite stress. Aquat. Toxicol. 2021, 237, 105899. [Google Scholar] [CrossRef]
- Gao, Y.C.; Wei, Y.F.; Cao, D.N.; Ge, Y.; Gong, S.P. Transcriptome analysis reveals decreased immunity under heat stress in Mauremys mutica. Aquaculture 2021, 531, 735894. [Google Scholar] [CrossRef]
- Larhammar, D.; Lagman, D. Turtle ghrelin. Nat. Genet. 2014, 46, 524–525. [Google Scholar] [CrossRef] [PubMed]
- Tine, M.; Kuhl, H.; Teske, P.R.; Tschöp, M.H.; Jastroch, M. Diversification and coevolution of the ghrelin/growth hormone secretagogue receptor system in vertebrates. Ecol. Evol. 2016, 6, 2516–2535. [Google Scholar] [CrossRef]
- Liu, L.; Jiang, G.M.; Peng, Z.T.; Li, Y.L.; Li, J.L.; Zou, L.; He, Z.G.; Wang, X.Q.; Chu, W.Y. The effect of high fat diet on daily rhythm of the core clock genes and muscle functional genes in the skeletal muscle of Chinese soft-shelled turtle (Trionyx sinensis). Comp. Biochem. Phys. 2017, 213, 17–27. [Google Scholar] [CrossRef]
- Zhang, J.; Yu, P.; Zhao, Y.; Zhou, Q.Y.; Yang, J.Y.; Hu, Q.T.; Liu, T.T.; Bao, C.H.; Su, S.P.; Gui, J.F. Global analysis of transcriptome and translatome revealed that coordinated WNT and FGF regulate the carapacial ridge development of Chinese soft-shell turtle. Int. J. Mol. Sci. 2021, 22, 12441. [Google Scholar] [CrossRef]
- Zeng, D.; Xiong, G.; Wang, X.Q.; Zhou, X.W.; Wang, P.; Peng, N.; Luo, Z.J.; Li, X. Transcriptome analysis and identification of genes and single-nucleotide polymorphisms associated with growth traits in the Chinese soft-shelled turtle, Pelodiscus sinensis. J. World Aquac. Soc. 2021, 52, 913–931. [Google Scholar] [CrossRef]
- Kaizuka, T.; Hara, T.; Oshiro, N.; Kikkawa, U.; Yonezawa, K.; Takehana, K.; Iemura, S.; Natsume, T.; Mizushima, N. Tti1 and Tel2 are critical factors in mammalian target of rapamycin complex assembly. J. Biol. Chem. 2010, 285, 20109–20116. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.R.; Huang, X.L.; Zhou, Z.C.; Xin, L. An improvement of the 2(−delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]
- Asad Samani, L.; Ghaedi, K.; Majd, A.; Peymani, M.; Etemadifar, M. Coordinated modification in expression levels of HSPA1A/B, DGKH, and NOTCH2 in Parkinson’s patients’ blood and substantia nigra as a diagnostic sign: The transcriptomes’ relationship. Neurol. Sci. 2023, 44, 2753–2761. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Lu, Y.N.; Li, N.; Shi, H.T.; Ding, L.; Hong, M.L.; Fang, Z.H. Effect of ammonia stress on AMPK regulating-carbohydrate and lipid metabolism in Chinese striped-neck turtle (Mauremys sinensis). Comp. Biochem. Phys. C 2023, 263, 109491. [Google Scholar] [CrossRef] [PubMed]
- Loannilli, L.; Ciccarone, F.; Ciriolo, M.R. Adipose tissue and FoxO1: Bridging physiology and mechanisms. Cells 2020, 9, 849. [Google Scholar] [CrossRef] [PubMed]
- Bentley, B.P.; Haas, B.J.; Tedeschi, J.N.; Berry, O. Loggerhead sea turtle embryos (Caretta caretta) regulate expression of stress response and developmental genes when exposed to a biologically realistic heat stress. Mol. Ecol. 2017, 26, 2978–2992. [Google Scholar] [CrossRef] [PubMed]
- Biggar, K.K.; Zhang, J.; Storey, K.B. Navigating oxygen deprivation: Liver transcriptomic responses of the red eared slider turtle to environmental anoxia. PeerJ 2019, 7, e8144. [Google Scholar] [CrossRef]
- Milton, S.L.; Dirk, L.J.; Kara, L.F.; Prentice, H.M. Adenosine modulates ERK1/2, PI3K/Akt, and p38MAPK activation in the brain of the anoxia-tolerant turtle Trachemys scripta. J. Cerebr. Blood Met. 2008, 28, 1469–1477. [Google Scholar] [CrossRef]
- Lv, Z.; Hu, Y.Z.; Tan, J.; Wang, X.Q.; Liu, X.Y.; Zeng, C. Comparative transcriptome analysis reveals the molecular immunopathogenesis of Chinese Soft-Shelled Turtle (Trionyx sinensis) Infected with Aeromonas hydrophila. Biology 2021, 10, 1218. [Google Scholar] [CrossRef]
- Ouyang, J.H.; Gao, Y.C.; Wei, Y.F.; Huang, H.P.; Ge, Y.; Zhao, J.; Gong, S.P. Transcriptome analysis reveals reduced immunity and metabolic level under cold stress in Mauremys mutica. Front. Mar. Sci. 2023, 10, 1224166. [Google Scholar] [CrossRef]
- Baglia, M.L.; Cai, Q.Y.; Zheng, Y.; Wu, J.; Su, Y.H.; Ye, F.; Bao, P.P.; Cai, H.; Zhao, Z.G.; Balko, J.; et al. Dual specificity phosphatase 4 gene expression in association with triple-negative breast cancer outcome. Breast Cancer Res. Treat. 2014, 148, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Midwood, K.S.; Chiquet, M.; Tucker, R.P.; Orend, G. Tenascin-C at a glance. J. Cell Sci. 2016, 129, 4321–4327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Chen, B.J.; Yuan, L.; Niu, C.J. Acute cold stress improved the transcription of pro-inflammatory cytokines of Chinese soft-shelled turtle against Aeromonas hydrophila. Dev. Comp. Immunol. 2015, 49, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Cocci, P.; Mosconi, G.; Palermo, F.A. Organic UV filters induce toll-like-receptors and related signaling pathways in peripheral blood mononuclear cells of juvenile loggerhead sea turtles (Caretta caretta). Animals 2012, 12, 594. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.J.; Wang, Q.; Wang, F.; Li, X.; Wang, B.K.; Zhou, Y.H.; Zou, P.; Tang, L.; Yu, D.Y.; Li, W.F. Improved immune function of Chinese soft-shelled turtles (Pelodiscus sinensis) through oral probiotics via the TLR signaling pathway. Aquaculture 2022, 555, 738126. [Google Scholar] [CrossRef]
- Zheng, C.; Zhao, Q.Q.; Li, E.C.; Zhao, D.X.; Sun, S.M. Role of hypoxia in the behaviour, physiology, immunity and response mechanisms of crustaceans: A review. Rev. Aquac. 2022, 14, 676–687. [Google Scholar] [CrossRef]
- Qu, D.Q.; Ludwig, D.S.; Gammeltoft, S.; Piper, M.; Pelleymounter, M.A.; Cullen, M.J.; Mathes, W.F.; Przypek, J.; Kanarek, R.; Maratos-Flier, E. A role for melanin-concentrating hormone in the central regulation of feeding behaviour. Nature 1996, 380, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, K.S.; Lauer, L.T.; Hayes, A.M.R.; Décarie-Spain, L.; McBurnett, K.; Nourbash, A.C.; Donohue, K.N.; Kao, A.E.; Bashaw, A.G.; Burdakov, D.; et al. Hypothalamic melanin-concentrating hormone neurons integrate food-motivated appetitive and consummatory processes in rats. Nat. Commun. 2023, 14, 1755. [Google Scholar] [CrossRef]
- Blanco, A.M.; Soengas, J.L. Leptin signalling in teleost fish with emphasis in food intake regulation. Mol. Cell. Endocrinol. 2021, 526, 111209. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Cabrera-Álvarez, M.J.; Maia, C.M.; Saraiva, J.L. Environmental enrichment in fish aquaculture: A review of fundamental and practical aspects. Rev. Aquac. 2022, 14, 704–728. [Google Scholar] [CrossRef]
- Lee, V.E.; Arnott, G.; Turner, S.P. Social behavior in farm animals: Applying fundamental theory to improve animal welfare. Front. Vet. Sci. 2022, 9, 932217. [Google Scholar] [CrossRef]
- Zerr, K.M.; Imlay, T.L.; Horn, A.G.; Slater, K.Y. Sick of attention: The effect of a stress-related disease on juvenile green sea turtle behaviour in the face of intense and prolonged tourism. Aquat. Conserv. 2022, 32, 430–441. [Google Scholar] [CrossRef]
- Dawkins, M.S. Natural behaviour is not enough: Farm animal welfare needs modern answers to Tinbergen’s four questions. Animals 2023, 13, 988. [Google Scholar] [CrossRef]
- Lu, X.; Dawood, M.A.O.; Wu, F.; Wen, H.; Liu, W.; Tian, J.; Jiang, M.; Yu, L.J.; Li, X.; Xu, N.; et al. Effect of Dietary L-Carnitine Supplementation on Growth, Serum Biochemical Indices, Oxidative and Growth-Related Gene Expressions in Chinese Soft-Shelled Turtle (Pelodiscus sinensis). Pak. J. Zool. 2022, 54, 2365–2372. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Gender | FBW (g) | SGR (%/d) | HIS (%) | ISI (%) | GSI (%) | LFI (%) | MFI (%) |
|---|---|---|---|---|---|---|---|---|
| PT | ♂ | 1298.7 ± 68.2 c | 0.60 ± 0.04 d | 3.48 ± 0.47 a | 1.38 ± 0.04 a | 1.65 ± 0.13 b | 6.59 ± 0.39 b | 1.27 ± 0.12 b |
| ♀ | 1073.3 ± 46.2 b | 0.39 ± 0.03 c | 4.47 ± 0.32 b | 1.61 ± 0.11 b | 2.70 ± 0.30 c | 8.48 ± 0.51 d | 1.54 ± 0.10 c | |
| RT | ♂ | 1029 ± 79.8 ab | 0.34 ± 0.01 b | 3.13 ± 0.25 a | 1.61 ± 0.15 b | 1.31 ± 0.14 a | 4.39 ± 0.33 a | 0.81 ± 0.07 a |
| ♀ | 985.2 ± 54.1 a | 0.29 ± 0.01 a | 4.16 ± 0.25 b | 1.68 ± 0.09 b | 2.85 ± 0.32 c | 7.19 ± 0.68 c | 1.17 ± 0.11 b |
| Groups | GLU (mmol/L) | TG (mmol/L) | TCHO (mmol/L) | CRE (μmol/L) | UA (mg/L) | BUN (mmol/L) |
|---|---|---|---|---|---|---|
| RT (♂) | 1.92 ± 0.62 a | 0.88 ± 0.11 a | 4.54 ± 0.18 a | 28.74 ± 3.33 b | 45.22 ± 3.55 c | 4.54 ± 0.23 c |
| RT (♀) | 5.73 ± 1.56 b | 5.6 ± 0.37 b | 8.53 ± 0.43 b | 39.04 ± 5.98 c | 26.92 ± 2.96 ab | 2.60 ± 0.14 a |
| PT (♂) | 9.93 ± 1.99 c | 0.85 ± 0.10 a | 4.66 ± 0.12 a | 20.60 ± 4.23 a | 28.75 ± 3.70 b | 2.91 ± 0.37 b |
| PT (♀) | 10.11 ± 2.84 c | 6.5 ± 0.52 b | 9.35 ± 0.56 b | 31.30 ± 5.50 b | 22.60 ± 3.54 a | 2.41 ± 0.16 a |
| Groups | AKP (U/L) | ACP (U/L) | GOT (U/L) | GPT (U/L) |
|---|---|---|---|---|
| RT (♂) | 1508.7 ± 52.9 d | 74.3 ± 5.3 b | 47.9 ± 5.5 b | 19.5 ± 3.0 a |
| RT (♀) | 1142.8 ± 60.5 b | 69.0 ± 6.0 b | 42.5 ± 3.9 b | 38.8 ± 4.4 d |
| PT (♂) | 1392.3 ± 59.8 c | 47.2 ± 5.9 a | 32.5 ± 4.2 a | 26.5 ± 3.5 b |
| PT (♀) | 1011.6 ± 61.4 a | 46.8 ± 4.6 a | 30.7 ± 5.1 a | 30.1 ± 3.9 c |
| Groups | CAT (U/mL) | GSH-PX (U/mL) | SOD (U/mL) | LZM (U/mL) | C3 (μg/mL) |
|---|---|---|---|---|---|
| RT (♂) | 7.15 ± 1.03 b | 367.9 ± 10.2 | 100.5 ± 5.2 | 698.7 ± 55.8 ab | 5.21 ± 1.11 |
| RT (♀) | 6.64 ± 1.09 b | 361.8 ± 5.6 | 108.2 ± 9.9 | 637.5 ± 37.4 a | 4.96 ± 0.93 |
| PT (♂) | 4.43 ± 0.84 a | 370.0 ± 5.9 | 113.4 ± 11.3 | 728.9 ± 42.1 b | 5.86 ± 1.14 |
| PT (♀) | 4.09 ± 0.89 a | 357.3 ± 12.7 | 101.4 ± 8.6 | 700.3 ± 50.0 ab | 5.66 ± 1.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, B.; Huang, L.; Wu, C.; Chen, J.; Chen, X.; He, J. Comparative Analysis of the Growth, Physiological Responses, and Gene Expression of Chinese Soft-Shelled Turtles Cultured in Different Modes. Animals 2024, 14, 962. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060962
Wu B, Huang L, Wu C, Chen J, Chen X, He J. Comparative Analysis of the Growth, Physiological Responses, and Gene Expression of Chinese Soft-Shelled Turtles Cultured in Different Modes. Animals. 2024; 14(6):962. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060962
Chicago/Turabian StyleWu, Benli, Long Huang, Cangcang Wu, Jing Chen, Xiajun Chen, and Jixiang He. 2024. "Comparative Analysis of the Growth, Physiological Responses, and Gene Expression of Chinese Soft-Shelled Turtles Cultured in Different Modes" Animals 14, no. 6: 962. https://0-doi-org.brum.beds.ac.uk/10.3390/ani14060962