Provision Point-Source Materials Stimulates Play in Sows but Does Not Affect Aggression at Regrouping

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management
2.2. Housing
2.3. Treatment
2.4. Behavioral Observations
2.5. Injury Counts
2.6. Saliva Sample Collection and Analysis
2.7. Reproductive Measures
2.8. Sow Classification Calculation
2.9. Statistical Analyses
3. Results
3.1. Behavioural Observations
3.1.1. Fights and Displacements
3.1.2. Bites and Knocks
3.1.3. Eating and Inactivity
3.1.4. Play and Exploration
3.1.5. Time Budgets
3.2. Injuries
3.3. Salivary Cortisol Concentration
3.4. Sow Weight and Reproduction
3.5. Hierarchy
4. Discussion
4.1. Effect of Pont-Source Materials on Aggression and Injuries
4.2. Days after Mixing
4.3. Success of the Materials in Creating Posative Behaviours
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newberry, R.C.; WoodGush, D.G.M.; Hall, J.W. Playful behaviour of piglets. Behav. Proc. 1988, 17, 205–216. [Google Scholar] [CrossRef]
- Van de Weerd, H.A.; Day, J.E. A review of environmental enrichment for pigs housed in intensive housing systems. App. Anim. Behav. Sci. 2009, 116, 1–20. [Google Scholar] [CrossRef]
- Elmore, M.R.P.; Garner, J.P.; Johnson, A.K.; Kirkden, R.D.; Richert, B.T.; Pajor, E.A. Getting around social status, motivation and enrichment use of dominant and subordinate sows in a group setting. Appl. Anim. Behav. Sci. 2011, 133, 154–163. [Google Scholar] [CrossRef]
- Schaefer, A.; Salomons, M.; Tong, A.; Sather, A.; Lepage, P. The effect of environment enrichment on aggression in newly weaned pigs. Appl. Anim. Behav. Sci. 1990, 27, 41–52. [Google Scholar] [CrossRef]
- Beattie, V.; Walker, N.; Sneddon, I. An investigation of the effect of environmental enrichment and space allowance on the behaviour and production of growing pigs. Appl. Anim. Behav. Sci. 1996, 48, 151–158. [Google Scholar] [CrossRef]
- Blackshaw, J.K.; Thomas, F.J.; Lee, J.-A. The effect of a fixed or free toy on the growth rate and aggressive behaviour of weaned pigs and the influence of hierarchy on initial investigation of the toys. Appl. Anim. Behav. Sci. 1997, 53, 203–212. Available online: https://habricentral.org/resources/9589 (accessed on 1 November 2018). [CrossRef]
- Beattie, V.; O’Connell, N.; Moss, B. Influence of environmental enrichment on the behaviour, performance and meat quality of domestic pigs. Livest. Prod. Sci. 2000, 65, 71–79. Available online: https://pdfs.semanticscholar.org/1403/db920cc8a76111c0b394ef9c9e7a505d05d1.pdf (accessed on 1 November 2018). [CrossRef]
- Olsen, A.; Simonsen, H.; Dybkjraer, L. Effect of access to roughage and shelter on selected behavioural indicators of welfare in pigs housed in a complex environment. Anim. Welf. 2002, 11, 75–87. Available online: https://0-adelaideaus-library-ingentaconnect-com.brum.beds.ac.uk/content/ufaw/aw/2002/00000011/00000001/art00006 (accessed on 6 November 2018).
- Jensen, K.H.; Sørensen, L.S.; Bertelsen, D.; Pedersen, A.R.; Jørgensen, E.; Nielsen, N.P.; Vestergaard, K.S. Management factors affecting activity and aggression in dynamic group-housing systems with electronic sow feeding: A field trial. Anim. Sci. 2000, 71, 535–545. [Google Scholar] [CrossRef]
- Durrell, J.; Sneddon, I.; Beattie, V. Effects of enrichment and floor type on behaviour of cubicle loose-housed dry sows. Anim. Welf. 1997, 6, 297–308. [Google Scholar]
- Andersen, I.; Bøe, K. Straw bedding or concrete floor for loose-housed pregnant sows: Consequences for aggression, production and physical health. Acta Agric. Scand. A Anim. Sci. 1999, 49, 190–195. [Google Scholar] [CrossRef]
- Whittaker, X.; Edwards, S.A.; Spoolder, H.A.M.; Lawrence, A.B.; Corn, S. Effects of straw bedding and high fibre diets on the, behaviour of floor fed group-housed sows. Appl. Anim. Behav. Sci. 1999, 63, 25–39. [Google Scholar] [CrossRef]
- Stewart, C.L.; O’Connell, N.E.; Boyle, L. Influence of access to straw provided in racks on the welfare of sows in large dynamic groups. Appl. Anim. Behav. Sci. 2008, 112, 235–247. [Google Scholar] [CrossRef]
- Studnitz, M.; Jensen, M.B.; Pedersen, L.J. Why do pigs root and in what will they root?: A review on the exploratory behaviour of pigs in relation to environmental enrichment. Appl. Anim. Behav. Sci 2007, 107, 183–197. [Google Scholar] [CrossRef]
- Van de Weerd, H.A.; Docking, C.M.; Day, J.E.; Avery, P.J.; Edwards, S.A. A systematic approach towards developing environmental enrichment for pigs. Appl. Anim. Behav. Sci. 2003, 84, 101–118. [Google Scholar] [CrossRef]
- Boyle, L.; Carroll, C.; McCutcheon, G.; Clarke, S.; McKeon, M.; Lawloe, P.; Ryan, T.; Ryan, T.; Fitzgerald, T.; Quinn, A.; et al. Towards January 2012: Updates Implications and Options for Group Housing Pregnant Sows; Pig Development Department of the Agriculture and Food Development Authority: Carlow, Ireland, 2012. [Google Scholar]
- Day, J.E.L.; Spoolder, H.A.M.; Burfoot, A.; Chamberlain, H.L.; Edwards, S.A. The separate and interactive effects of handling and environmental enrichment on the behaviour and welfare of growing pigs. Appl. Anim. Behav. Sci. 2002, 75, 177–192. [Google Scholar] [CrossRef]
- Muller, T.L.; Callaghan, M.J.; Hewitt, R.J.E.; D’Souza, D.N.; van Barneveld, R.J. A poured block reduces feeding associated aggression in sows during gestation. Anim. Prod. Sci. 2017, 57, 2458. Available online: https://www.publish.csiro.au/an/Fulltext/anv57n12ab061 (accessed on 11 December 2018). [CrossRef]
- Graf, R. Das Visuelle Visuelle Orientierungsvermögen der Schweine in Abhängigkeit von der Beleuchtungsstärke. 1976. Available online: http://rn.ptz.icm.edu.pl/wp-content/uploads/2017/01/11-Klocek-ang.-pdf.pdf (accessed on 1 November 2018).
- Deligeorgis, S.T.; Karalis, K.; Kanzouros, G. The influencs of drinker location and colour on drinking behaviour and water intake of newborn pigs under hot environments. Appl. Anim. Behav. Sci. 2006, 96, 233–244. [Google Scholar] [CrossRef]
- Edge, L.; Varley, A.; Rowlinson, P. The effect of trough colour on the voluntary food intake of postweaned pigs—A short communication. Livest. Prod. Sc. 2004, 88, 193–197. [Google Scholar] [CrossRef]
- Tanida, H.; Senda, K.; Suzuki, S.; Tanaka, T.; Yoshimoto, T. Color discrimination in weaning pigs. Anim. Feed Sci. Technol. 1991, 62, 1029–1034. [Google Scholar]
- Apple, J.; Craig, J. The influence of pen size on toy preference of growing pigs. Appl. Anim. Behav. Sci. 1992, 35, 149–155. [Google Scholar] [CrossRef]
- Feddes, J.; Fraser, D. Non-nutritive chewing by pigs: Implications for tail-biting and behavioral enrichment. Trans. ASAE 1994, 37, 947–950. [Google Scholar] [CrossRef]
- NHMRC. Australian Code of Practice for the Care and Use of Animals for Scientific Purposes. 2013. Available online: https://nhmrc.gov.au/about-us/publications/australian-code-care-and-use-animals-scientific-purposes#block-views-block-file-attachments-content-block-1 (accessed on 6 November 2018).
- Australian Government. Bureau of Meteorology: Climate Data Online. Available online: http://www.bom.gov.au/climate/data/ (accessed on 3 December 2018).
- Karlen, G.A.M.; Hemsworth, P.H.; Gonyou, H.W.; Fabrega, E.; Strom, A.D.; Smits, R.J. The welfare of gestating sows in conventional stalls and large groups on deep litter. Appl. Anim. Behav. Sci. 2007, 105, 87–101. [Google Scholar] [CrossRef]
- Greenwood, E.C.; Plush, K.J.; van Wettere, W.H.; Hughes, P.E. Group and individual sow behavior is altered in early gestation by space allowance in the days immediately following grouping. J. Anim. Sci. 2016, 94, 385–393. [Google Scholar] [CrossRef]
- Greenwood, E.C.; Plush, K.J.; van Wettere, W.H.E.J.; Hughes, P.E. A novel method for the analysis of social structure allows in-depth analysis of sow rank in newly grouped sows. Appl. Anim. Behav. Sci. 2017, 189, 29–35. [Google Scholar] [CrossRef]
- Horback, K.M.; Pierdon, M.K.; Parsons, T.D. Behavioral preference for different enrichment objects in a commercial sow herd. Appl. Anim. Behav. Sci. 2016, 184, 7–15. [Google Scholar] [CrossRef]
- Bulens, A.; Van Beirendonck, S.; Van Thielen, J.; Buys, N.; Driessen, B. Hiding walls for fattening pigs: Do they affect behavior and performance? Appl. Anim. Behav. Sci. 2017, 195, 32–37. [Google Scholar] [CrossRef]
- Waran, N.K.; Broom, D.M. The influence of a barrier on the behaviour and growth of early-weaned piglets. Anim. Sci. 1993, 56, 115–119. [Google Scholar] [CrossRef]
- Pedersen, L.J.; Hansen, S.W.; Munksgaard, L.; Ladewig, J.; Matthews, L. Social isolation affects the motivation to work for food and straw in pigs as measured by operant conditioning techniques. Appl. Anim. Behav. Sci. 2002, 77, 295–309. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Smith, K.; Karlen, M.G.; Arnold, N.A.; Moeller, S.J.; Barnett, J.L. The choice behaviour of pigs in a Y maze: Effects of deprivation of feed, social contact and bedding. Behav. Process. 2011, 87, 210–217. [Google Scholar] [CrossRef]
- De Jong, I.C.; Ekkel, E.D.; van de Burgwal, J.A.; Lambooij, E.; Korte, S.M.; Ruis, M.A.W.; Koolhaas, J.M.; Blokhuis, H.J. Effects of straw bedding on physiological responses to stressors and behavior in growing pigs. Physiol. Behav. 1998, 64, 303–310. [Google Scholar] [CrossRef]
- De Groot, J.I.; Prelle, I.T.; Koolhaas, J.M. Immunity in barren and enriched housed pigs differing in baseline cortisol concentration. Physiol. Behav. 2000, 71, 217–223. [Google Scholar] [CrossRef] [Green Version]
- De Jong, I.C.; Prelle, I.T.; van de Burgwal, J.A.; Lambooij, E.; Korte, S.M.; Blokhuis, H.J.; Koolhaas, J.M. Effects of environmental enrichment on behavioral responses to novelty, learning, and memory, and the circadian rhythm in cortisol in growing pigs. Physiol. Behav. 2000, 68, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Reimert, I.; Bolhuis, J.E.; Kemp, B.; Rodenburg, T.B. Indicators of positive and negative emotions and emotional contagion in pigs. Physiol. Behav. 2013, 109, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Ko, H.L.; Salazar, L.C.; Llonch, L.; Manteca, X.; Camerlink, I.; Llonch, P. Pre-weaning environmental enrichment increases piglets’ object play behaviour on a large scale commercial pig farm. Appl. Anim. Behav. Sci. 2018, 202, 7–12. [Google Scholar] [CrossRef]
- Tilbrook, A.; Turner, A.; Clarke, I. Stress and reproduction: Central mechanisms and sex differences in non-rodent species. Stress 2002, 5, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.I.; Hemsworth, P.H.; Tilbrook, A.J. Susceptibility of reproduction in female pigs to impairment by stress and the role of the hypothalamo-pituitary-adrenal axis. Reprod. Fertil. Dev. 2002, 14, 377–391. [Google Scholar] [CrossRef]
- Turner, A.I.; Keating, C.L.; Tilbrook, A.J. Sex Differences and the Role of Sex Steroids in Sympatho-Adrenal Medullary System and Hypothalamo-Pituitary Adrenal Axis Responses to Stress. Sex Steroids; InTech: Rijeka, Croatia, 2012; pp. 115–136. Available online: http://hdl.handle.net/10536/DRO/DU:30046471 (accessed on 6 November 2018).
- Krackow, S. Potential mechanisms for sex ratio adjustment in mammals and birds. Biol. Rev. 1995, 70, 225–241. [Google Scholar] [CrossRef]
- Mount, N.C.; Seabrook, M.F. A study of aggression when group housed sows are mixed. Appl. Anim. Behav. Sci. 1993, 36, 377–383. [Google Scholar] [CrossRef]
- Moore, A.; Gonyou, H.; Ghent, A. Integration of newly introduced and resident sows following grouping. Appl. Anim. Behav. Sci. 1993, 38, 257–267. [Google Scholar] [CrossRef]
- Arey, D.S. Time course for the formation and disruption of social organisation in group housed sows. Appl. Anim. Behav. Sci. 1999, 62, 199–207. [Google Scholar] [CrossRef]
- Zurbrigg, K.; Blackwell, T. Injuries, lameness and cleanliness of sows in four group housing gestation facilities in Ontario. J. Swine Health Prod. 2005, 14, 202–206. [Google Scholar]
- Greenwood, E.C.; Plush, K.J.; van Wettere, W.H.; Hughes, P.E. Hierarchy formation in newly mixed, group housed sows and management strategies aimed at reducing its impact. Appl. Anim. Behav. Sci. 2014, 160, 1–11. [Google Scholar] [CrossRef]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Mendl, M.; Burman, O.H.P.; Parker, R.M.A.; Paul, E.S. Cognitive bias as an indicator of animal emotion and welfare: Emerging evidence and underlying mechanisms. Appl. Anim. Behav. Sci. 2009, 118, 161–181. [Google Scholar] [CrossRef]
- Hemsworth, P.H.; Mellor, D.J.; Cronin, G.M.; Tilbrook, A.J. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef]
- Held, S.D.E.; Špinka, M. Animal play and animal welfare. Anim. Behav. 2011, 81, 891–899. [Google Scholar] [CrossRef]
- Horback, K. Nosing around: Play in pigs. Anim. Behav. Cogn. 2007, 1, 186–196. [Google Scholar] [CrossRef]
- Ahloy-Dallaire, J.; Espinosa, J.; Mason, G. Play and optimal welfare: Does play indicate the presence of positive affective states? Behav. Process. 2018, 156, 3–15. [Google Scholar] [CrossRef]
- Blois-Heulin, C.; Rochais, C.; Camus, S.; Fureix, C.; Lemasson, A.; Lunel, C.; Bezard, E.; Hausberger, M. Animal welfare: Could adult play be a false friend? Anim. Behav. Cogn. 2015, 2, 156–185. [Google Scholar] [CrossRef]



{kind=link}
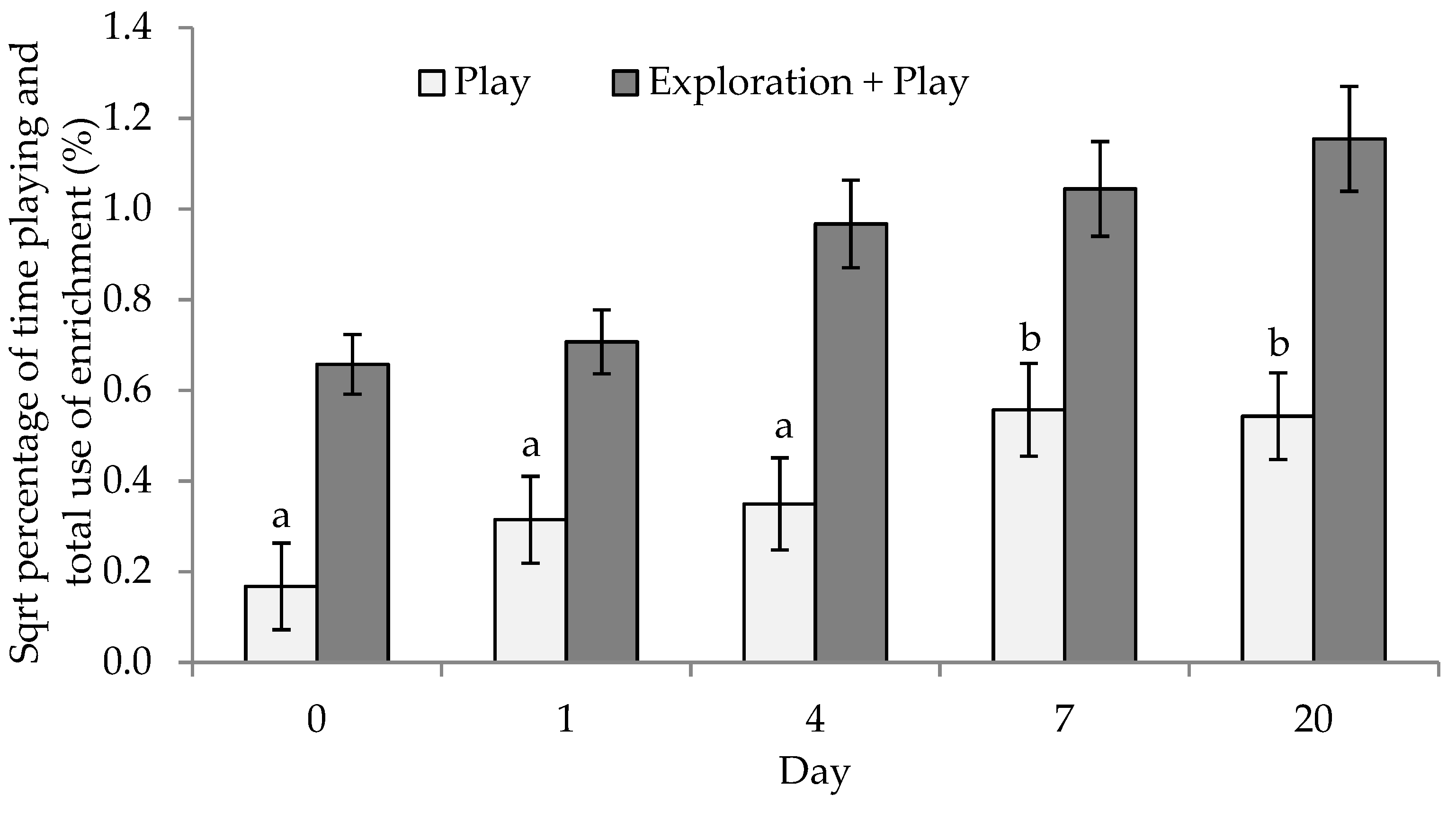{kind=link}
| Active, lying, inactive, eating and drinking | Sows were classed as eating if food was present and she was noted as collecting food from the floor, chewing and/or swallowing. The sow was drinking if her head was in the drinker and she could be seen to swallow and/or actively manipulate the drinker nipple. If it was unclear what the sow was doing and if she was dog-sitting, standing or walking she was considered active. Sows were coded as lying down if lying flat to the floor either on their side or on their belly. Sows were considered inactive if lying down. | Continuous |
| Exploring | Actively manipulating and exploring the surrounding environment, such as rooting, nosing the floor, moving drinkers and chewing fences. When concerning the materials, this was coded when the animal manipulated the rope, disk swing or mat (mainly chewing or nosing the material). Exploration is analysed for separate material use, overall material use, and exploration, except for materials and exploration total. | Continuous |
| Fighting | Aggression including three or more knocks or bites. Aggression can be reciprocal or non-reciprocal and was coded from a sow adopting a parallel pushing defensive posture as well as bite or knock interactions. | Continuous |
| Play | All locomotor and object play were included in one overall heading of ‘play’. Locomotor play included running, hopping (focal animal has either its two front feet or all four feet off the pen floor at one time through an energetic upwards jumping movement.), pivots (twirling of body on the horizontal plane by a minimum of 90° usually associated with jumping on the spot) and head tossing (quick and often erratic shaking of the head back and forth and up and down). Object play is when an animal manipulates an item or securely holds it in its mouth, energetically shaking it [1]. Object play was specifically coded if directed towards a specific object to allow analysis of play around/using the enrichment (rope, yellow disk swing, mat and play not centred on a specific object or other). | Continuous |
| Displacement | Movement of one sow by another, from a valued resource such as food, drinker or lying space (if multiple knocks or bites are required, this is a fight) | Point |
| Knock | One sow knocks another sow using her head and neck, contacting any part of the receiving sow | Point |
| Bite | One single bite delivered from one sow to any part of another | Point |
| Lunge | Sow lunges at another but does not make physical contact | Point |
| Flee | Sow moves herself quickly and as far away as the can get from another sow, in response to an aggressive action | Point |
| Mounting/mounted | One sow mounts another, with her front legs both over the back of the other animal. This behaviour was coded as long as the mounting animal remained on top of the mounted. Mounting and mounted were both scored as separate behaviours. | Point |
| Non-aggressive sow-sow contact | Mutual contact between two sows, which involves exploration of another animal with no aggressive outcomes (does not include lying with another sow) | Point |
| Guarding enrichment | Any form of aggression (knock, bite, lunge) or defensive posturing to stop other sows from the use of enrichment, whether resulting in a fight or displacement from enrichment. This behaviour could fall under displacement or fight if it fit into the below description, but was also specifically allowed for separately, in order to allow analysis of any aggression surrounding the enrichment (sows could be coded as guarding individual enrichment items, mat, disk swing or rope). | Point |
| Day | 0 | 1 | 4 | 7 | 20 | Tran. | p-Value |
|---|---|---|---|---|---|---|---|
| Fight number | 0.4 ± 0.1 a (3.4) | −0.4 ± 0.1 b (0.3) | −0.5 ± 0.1 b (0.3) | −0.7 ± 0.1 b (0.2) | −0.5 ± 0.2 b (0.1) | Log10 | <0.001 |
| Fight duration, sec | 1.9 ± 0.2 a (127.6) | 0.9 ± 0.2 b (13.0) | 0.8 ± 0.2 b (10.8) | 0.7 ± 0.3 b (14.9) | 0.6 ± 0.3 b (9.7) | Log10 | <0.001 |
| Time spent fighting, % | −0.4 ± 0.2 a (0.5) | −1.4 ± 0.2 b (0.06) | −1.8 ± 0.2 b (0.03) | −2.1 ± 0.3 b (0.1) | −1.9 ± 0.3 b (0.1) | Log10 | <0.001 |
| Displacement number | 2.0 ± 0.1 a (4.2) | 1.3 ± 0.1 b,c (1.9) | 1.4 ± 0.1 b (2.2) | ± 0.1 c (1.3) | 1.1 ± 0.1 c (1.1) | SQRT | <0.001 |
| Bite number | 1.3 ± 0.1 a (25.1) | 0.6 ± 0.1 b (4.9) | 0.6 ± 0.1 b (4.5) | 0.4 ± 0.1 c (2.9) | 0.3 ± 0.1 c (2.3) | Log10 | <0.001 |
| Knock number | 1.1 ± 0.1 a (14.5) | 0.6 ± 0.1 bc (4.0) | 0.7 ± 0.1 b (5.4) | 0.7 ± 0.1 b (4.6) | 0.5 ± 0.1 c (3.4) | Log10 | <0.001 |
| Injury count | 4.9 ± 0.2 a (24.7) | 5.5 ± 0.2 b (31.6) | 5.5 ± 0.2 b (31.3) | 5.4 ± 0.3 b (30.9) | 4.1 ± 0.2 c (17.9) | SQRT | <0.05 |
| Behavior | Total Time in Play, % | Total Time Exploring, % |
|---|---|---|
| Rope | 1.4 ± 0.1 (2.4) | 0.9 ± 0.2 (1.1) |
| Mat | 0.2 ± 0.1 (0.3) | 0.5 ± 0.3 (0.5) |
| Yellow disk swing | 0.3 ± 0.1 (0.1) | 0.1 ± 0.1 (0.1) |
| Behavior | Treatment | Day 0 | Day 1 | Day 4 | Day 7 | Day 20 |
|---|---|---|---|---|---|---|
| Active | enhanced | 35.55 ± 2.23 | 20.21 ± 2.11 | 22.64 ± 2.33 | 18.69 ± 2.32 | 27.22 ± 2.11 |
| standard | 33.66 ± 2.16 | 18.31 ± 2.11 | 20.74 ± 2.33 | 16.79 ± 2.32 | 25.32 ± 2.11 | |
| Resting | enhanced | 43.79 ± 2.68 | 62.89 ± 2.54 | 60.06 ± 2.81 | 64.12 ± 2.79 | 55.13 ± 2.54 |
| standard | 46.59 ± 2.60 | 65.69 ± 2.54 | 62.87 ± 2.80 | 66.93 ± 2.79 | 57.93 ± 2.54 | |
| Eating | enhanced | 5.43 ± 0.52 | 6.55 ± 0.49 | 7.09 ± 0.54 | 6.49 ± 0.54 | 6.98 ± 0.49 |
| standard | 5.08 ± 0.51 | 6.21 ± 0.49 | 6.75 ± 0.54 | 6.15 ± 0.54 | 6.64 ± 0.49 | |
| Exploring | enhanced | 3.16 ± 0.13 (10.28) | 2.75 ± 0.12 (7.72) | 2.65 ± 0.14 (7.3) | 2.65 ± 0.14 (7.35) | 2.96 ± 0.12 (8.93) |
| standard | 3.0 ± 0.13 (9.46) | 2.59 ± 0.12 (6.9) | 2.49 ± 0.14 (6.48) | 2.52 ± 0.14 (6.53) | 2.08 ± 0.12 (8.12) | |
| Fighting | enhanced | 0.51 ± 0.07 | 0.08 ± 0.06 | 0.05 ± 0.07 | 0.01 ± 0.07 | 0.03 ± 0.06 |
| standard | 0.46 ± 0.07 | 0.04 ± 0.06 | 0.00 ± 0.07 | 0.01 ± 0.07 | 0.01 ± 0.06 | |
| Mounting | enhanced | 1.42 ± 0.34 | 0.09 ± 0.33 | 0.16 ± 0.35 | 0.53 ± 0.35 | 0.09 ± 0.33 |
| standard | 1.41 ± 0.33 | 0.08 ± 0.33 | 0.16 ± 0.35 | 0.52 ± 0,35 | 0.08 ± 0.33 | |
| Play | enhanced | 0.3 ± 0.11 (1.46) | 0.6 ± 0.12 (1.65) | 1.1 ± 0.12 (2.55) | 0.7 ± 0.12 (2.22) | 1.1 ± 0.12 (2.98) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greenwood, E.C.; Van Wettere, W.H.E.J.; Rayner, J.; Hughes, P.E.; Plush, K.J. Provision Point-Source Materials Stimulates Play in Sows but Does Not Affect Aggression at Regrouping. Animals 2019, 9, 8. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9010008
Greenwood EC, Van Wettere WHEJ, Rayner J, Hughes PE, Plush KJ. Provision Point-Source Materials Stimulates Play in Sows but Does Not Affect Aggression at Regrouping. Animals. 2019; 9(1):8. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9010008
Chicago/Turabian StyleGreenwood, Emma Catharine, William H. E. J. Van Wettere, Jessica Rayner, Paul E. Hughes, and Kate J. Plush. 2019. "Provision Point-Source Materials Stimulates Play in Sows but Does Not Affect Aggression at Regrouping" Animals 9, no. 1: 8. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9010008