Effects of In Ovo Methionine-Cysteine Injection on Embryonic Development, Antioxidant Status, IGF-I and TLR4 Gene Expression, and Jejunum Histomorphometry in Newly Hatched Broiler Chicks Exposed to Heat Stress during Incubation

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Eggs and Incubation Conditions
2.2. Methionine-Cysteine In ovo Injection
2.3. Embryonic Development Index
2.4. Serum Biochemical Indicators
2.5. Jejunum Histomorphometry
2.6. Assay of Antioxidant Biomarkers in the Tissues and Serum
2.7. RNA Isolation and cDNA Synthesis
2.8. Quantitative Real-Time Reverse Transcription-PCR
2.9. Statistical Analysis
3. Results
3.1. Embryonic Development Parameters
3.2. Serum Biochemical Parameters
3.3. Antioxidant Biomarkers
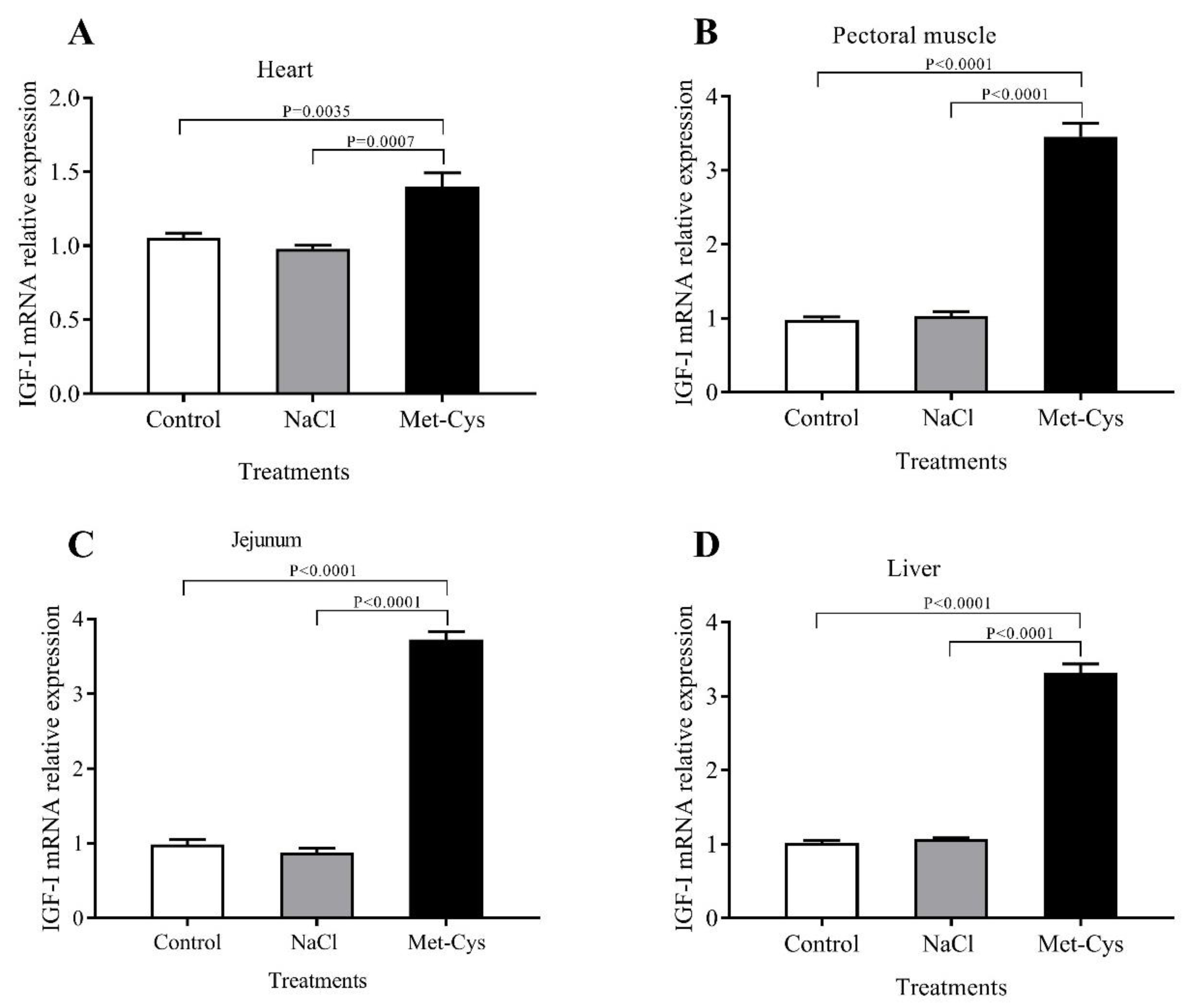
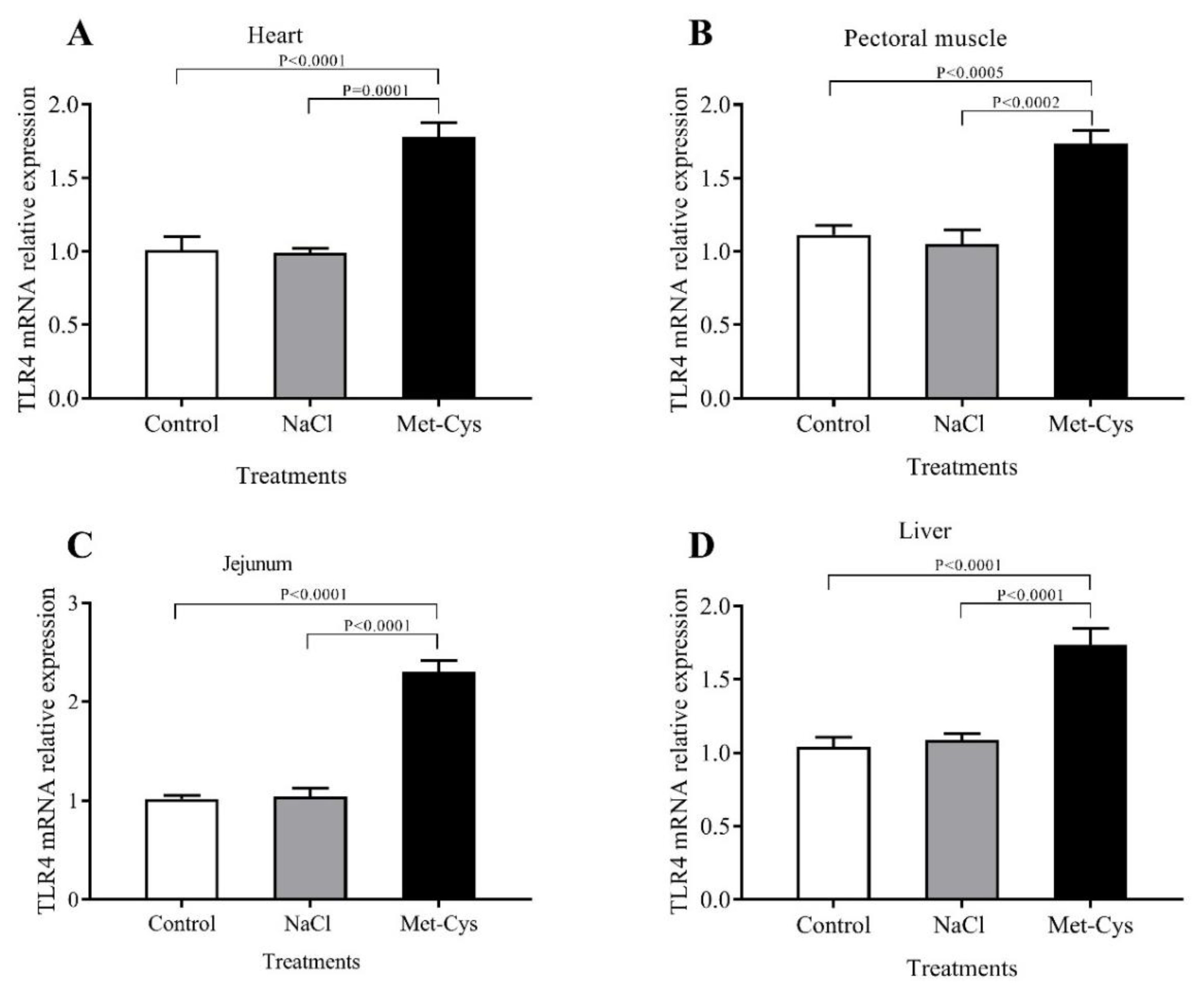
3.4. IGF-I and TLR-4 mRNA Relative Gene Expression
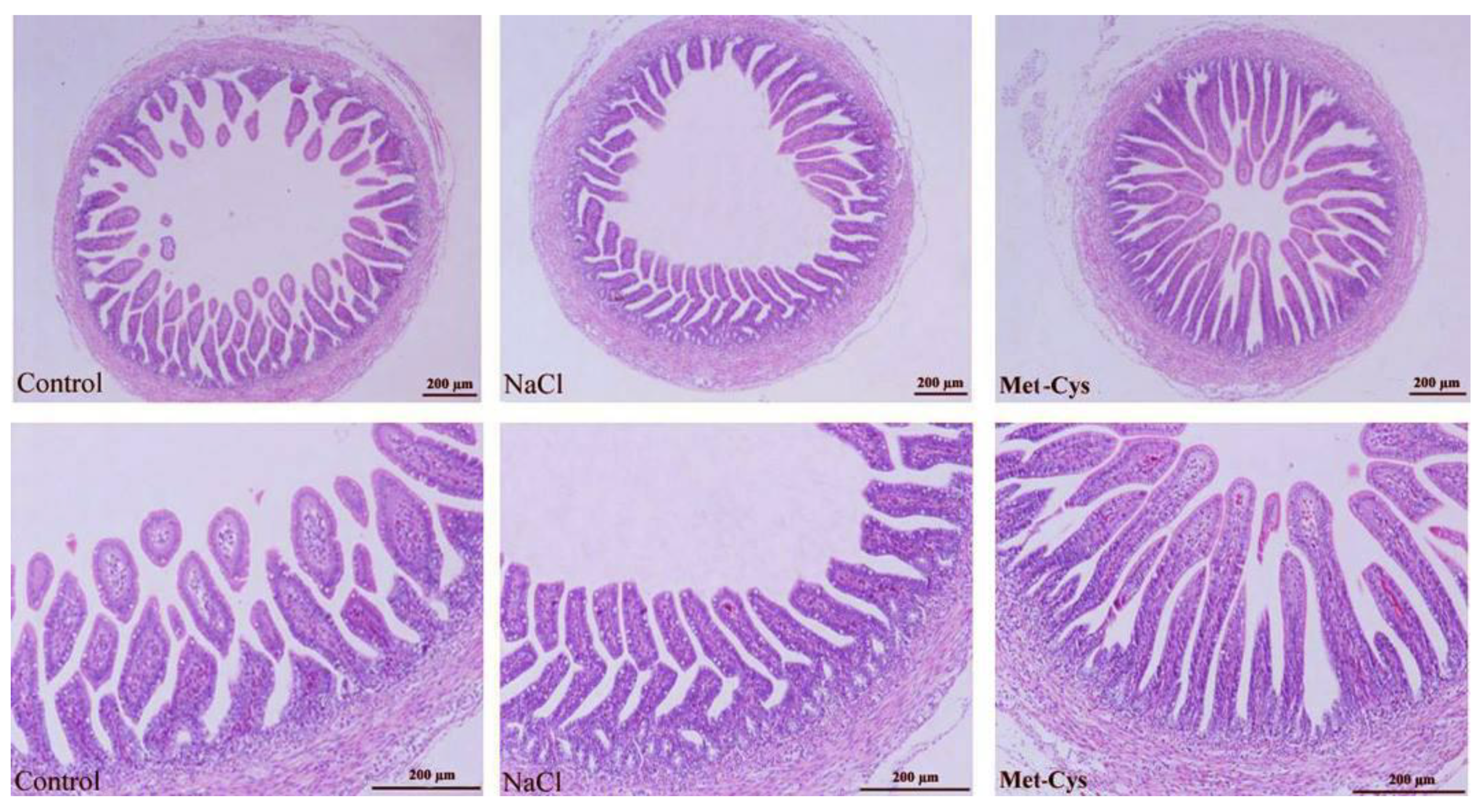
3.5. Jejunum Histomorphometric Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leandro, N.S.M.; Oliveira, A.S.C.; Gonzales, E.; Café, M.B.; Strighini, J.H.; Andrade, M.A. Probiótico na ração ou inoculado em ovos embrionados: Desempenho de pintos de corte desafiados com Salmonella Enteritidis. Rev. Bras. Zootec. 2010, 39, 1509–1516. [Google Scholar] [CrossRef]
- Shafey, T.M.; Mahmoud, A.H.; Alsobayel, A.A.; Abouheif, M.A. Effects of in ovo administration of amino acids on hatchability and performance of meat chickens. S. Afr. J. Anim. Sci. 2014, 44, 123–130. [Google Scholar] [CrossRef]
- Maltby, V.; Somaiya, A.; French, N.A.; Stickland, N.C. In ovo temperature manipulation influences post-hatch muscle growth in the turkey. Br. Poult. Sci. 2004, 45, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Oznurlu, Y.; Celik, I.; Telatar, T.; Sur, E. Histochemical and histological evaluations of the effects of high incubation temperature on embryonic development of thymus and bursa of Fabricius in broiler chickens. Br. Poult. Sci. 2010, 51, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Willemsen, H.; Kamers, B.; Dahlke, F.; Han, H.; Song, Z.; Ansari, P.Z.; Tona, K.; De-Cuypere, E.; Everaert, N. High-and low-temperature manipulation during late incubation: Effects on embryonic development, the hatching process, and metabolism in broilers. Poult. Sci. 2010, 89, 2678–2690. [Google Scholar] [CrossRef] [PubMed]
- Barri, A.; Honaker, C.F.; Sottosanti, J.R.; Hulet, R.M.; McElroy, A.P. Effect of incubation temperature on nutrient transporters and small intestine morphology of broiler chickens. Poult. Sci. 2011, 90, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Al-Zhgoul, M.B.; Dalab, A.E.S.; Ababneh, M.M.; Jawasreh, K.I.; Al-Busadah, K.A.; Ismail, Z.B. Thermal manipulation during chicken embryogenesis results in enhanced Hsp70 gene expression and the acquisition of thermotolerance. Res. Vet. Sci. 2013, 95, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Moseley, P.L. Heat shock proteins and heat adaptation of the whole organism. J. Appl. Psychol. 1997, 83, 1413–1417. [Google Scholar] [CrossRef]
- (NRC) National Research Council. Nutrient Requirements of Poultry, 9th revised ed.; National Academic Press: Washington, DC, USA, 1994. [Google Scholar]
- Kidd, MT. Nutritional Modulation of Immune Function in Broilers. Poult. Sci. 2004, 83, 650–657. [Google Scholar] [CrossRef]
- Stipanuk, M.H. Sulfur amino acid metabolism: Pathways for production and removal of homocysteine and cysteine. Annu. Rev. Nutr. 2004, 24, 539–577. [Google Scholar] [CrossRef]
- Han, G.; Yang, H.; Bahry, M.A.; Tran, P.V.; Do, P.H.; Ikeda, H.; Furuse, M.; Chowdhury, V.S. L-Leucine acts as a potential agent in reducing body temperature at hatching and affords thermotolerance in broiler chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 204, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Bahry, M.A.; Hui, Y.; Furuse, M.; Chowdhury, V.S. Acute heat stress upregulates neuropeptide Y precursor mRNA expression and alters brain and plasma concentrations of free amino acids in chicks. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2015, 187, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ohta, Y.; Kidd, M.T.; Ishibashi, T. Embryo growth and amino acid concentration profiles of broiler breeder eggs, embryos, and chicks after in ovo administration of amino acids. Poult. Sci. 2001, 80, 1430–1436. [Google Scholar] [CrossRef] [PubMed]
- Nazem, M.N.; Sayed, M.S.; Reza, K.; Hamideh, M. Histomorphometric analysis of the small intestine of broiler chick embryos injected in ovo with methionine. Anim. Prod. Sci. 2017, 59, 133–139. [Google Scholar] [CrossRef]
- Foye, O.T.; Uni, Z.; Ferket, P.R. Effect of in ovo feeding egg white protein, β-hydroxy-β-methylbutyrate, and carbohydrates on glycogen status and neonatal growth of turkeys. Poult. Sci. 2006, 85, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, M.; Janmohammadi, H.; Kia, H.D.; Moghaddam, G.; Rajabi, Z.; Rafat, S.A.; Javanmard, A. The effect of L-lysine in ovo feeding on body weight characteristics and small intestine morphology in a day-old Ross broiler chicks. Rev. Med. Vet. 2017, 168, 116–125. [Google Scholar]
- Havenstein, G.B.; Ferket, P.R.; Qureshi, M.A. Carcass composition and yield of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 2003, 82, 1509–1518. [Google Scholar] [CrossRef] [Green Version]
- Uni, Z.; Ferket, P.R.; Tako, E.; Kendar, O. In ovo feeding improves energy status of late-term chicken embryos. Poult. Sci. 2005, 84, 764–770. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, V.S.; Shigemura, A.; Erwan, E.; Ito, K.; Bahry, M.A.; Phuong, T.V.; Furuse, M. Oral administration of l-citrulline, but not l-arginine or l-ornithine, acts as a hypothermic agent in chicks. J. Poult. Sci. 2015, 52, 331–335. [Google Scholar] [CrossRef]
- Sgavioli, S.; Matos Júnior, J.B.; Borges, L.L.; Praes, M.F.F.M.; Morita, V.S.; Zanirato, G.L.; Garcia, R.G.; Boleli, I.C. Effects of Ascorbic Acid Injection in Incubated Eggs Submitted to Heat Stress on Incubation Parameters and Chick Quality. Braz. J. Poult. Sci. 2015, 17, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Bhanja, S.K.; Mandal, A.B.; Johri, T.S. Standardization of injection site, needle length, embryonic age and concentration of amino acids for in ovo injection in broiler breeder eggs. Indian J. Poult. Sci. 2004, 39, 105–111. [Google Scholar]
- Keralapurath, M.M.; Corzo, A.; Pulikanti, R.; Zhai, W.; Peebles, E.D. Effects of in-ovo injection of l–carnitine on hatchability and subsequent broiler performance and slaughter yield. Poult. Sci. 2010, 89, 1497–1501. [Google Scholar] [CrossRef] [PubMed]
- Kisielinski, K.; Willis, S.; Prescher, A.; Klosterhalfen, B.; Schumpelick, V. A simple new method to calculate small intestine absorptive surface in the rat. Clin. Exp. Med. 2002, 2, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.Z.Z.Y.; Jiang, X.X.; Wang, Y.H.; Luo, X.F.; Li, H.L. Role of the insulin-like growth factor system in epiphyseal cartilage on the development of Langshan and Arbor Acres chickens, Gallus domesticus. Poult. Sci. 2010, 89, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.K.; Wu, Y.H.; An, J.; Zheng, S.J. Polymorphisms of chicken Toll-like receptors 4, 15, and 21 in different breeds. Poult. Sci. 2012, 91, 2512–2516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Yang, S.B.; Xu, F.F.; Zhang, J.H. Long noncoding RNA CCAT1 promotes hepatocellular carcinoma progression by functioning as let-7 sponge. J. Exp. Clin. Cancer Res. 2015, 34, 18. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Sukker, H.; Ababneh, M.M. Effect of thermal manipulation of broilers embryos on the response to heat-induced oxidative stress. Poult. Sci. 2018, 1–11. [Google Scholar] [CrossRef]
- Ohta, Y.; Tsushima, N.; Koide, K.; Kidd, M.T.; Ishibashi, T. Effect of amino acid injection in broiler breeder eggs on embryonic growth and hatchability of chicks. Poult. Sci. 1999, 78, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Uni, Z.; Ferket, P.R. Methods for early nutrition and their potential. Worlds Poult. Sci. J. 2004, 60, 101–111. [Google Scholar] [CrossRef]
- Ozaydın, T.; Celik, I. Effects of high incubation temperature on the body weight and yolk consumption of two commercial broiler strain. Acta Sci. Vet. 2014, 42, 1253. [Google Scholar]
- Yahav, S.; Sasson, R.; Shinder, D. The effect of thermal manipulations during embryogenesis of broiler chicks (Gallus domesticus) on hatchability, body weight and thermoregulation after hatch. J. Therm. Biol. 2004, 29, 245–250. [Google Scholar] [CrossRef]
- Lu, J.W.; McMurtry, J.P.; Coon, C.N. Developmental changes of plasma insulin, glucagon, insulin-like growth factors, thyroid hormones, and glucose concentrations in chick embryos and hatched chicks. Poult. Sci. 2007, 86, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Regulation of glutathione synthesis. Mol. Asp. Med. 2009, 30, 42–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.D.; Huang, B.; Tsuji, Y. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2017, 58, 287–293. [Google Scholar] [CrossRef]
- Shen, Y.B.; Ferket, P.; Park, I.; Malheiros, R.D.; Kim, S.W. Effects of feed grade l-methionine on intestinal redox status, intestinal development, and growth performance of young chickens compared with conventional DL-methionine. J. Anim. Sci. 2015, 93, 2977–2986. [Google Scholar] [CrossRef]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F.; Karadas, F.; Sparks, N.H. The importance of antioxidants in poultry. In Proceedings of the Annual North Carolina Poultry Nutrition Conference, NC, USA, 6–9 July 2003; pp. 38–56. [Google Scholar]
- Surai, P.F. Natural Antioxidant in poultry nutrition: New developments. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; pp. 669–676. [Google Scholar]
- Swennen, Q.; Geraert, P.A.; Mercier, Y.; Everaert, N.; Stinckens, A.; Willemsen, H.; Buyse, J. Effects of dietary protein content and 2-hydroxy-4-methylthiobutanoic acid or DL-methionine supplementation on performance and oxidative status of broiler chickens. Br. J. Nutr. 2011, 106, 1845–1854. [Google Scholar] [CrossRef]
- Voet, D.; Voet, J.G. Biochemistry, 2ed.; John Wiley and Sons, Inc.: New York, NY, USA, 1995. [Google Scholar]
- Jastrebski, S.F.; Lamont, S.J.; Schmidt, C.J. Chicken hepatic response to chronic heat stress using integrated transcriptome and metabolome analysis. PLoS ONE 2017, 12, e0181900. [Google Scholar] [CrossRef]
- Chen, Y.P.; Chen, X.; Zhang, H.; Zhou, Y.M. Effects of dietary concentrations of methionine on growth performance and oxidative status of broiler chickens with different hatching weight. Br. Poult. Sci. 2013, 54, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, K.; Mezes, M.; Gaal, T.; Bartos, A.; Balogh, K.; Husveth, F. Effect of supplementation with methionine and different fat sources on the glutathione redox system of growing chickens. Acta Vet. Hung. 2004, 52, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Shoveller, A.K.; Stoll, B.; Ball, R.O.; Burrin, D.G. Nutritional and functional importance of intestinal sulfur amino acid metabolism. J. Nutr. 2005, 135, 1609–1612. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. The sulfur-containing Amino acids: An overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef]
- Straus, D.S.; Takemoto, C.D. Specific decrease in liver insulin-like growth factor-I and brain insulin-like growth factor-II gene expression in energy-restricted rats. J. Nutr. 1991, 121, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Kita, K.; Nagao, K.; Taneda, N.; Inagaki, Y.; Hirano, K.; Shibata, T.; Yaman, M.A.; Conlon, M.A.; Okumura, J. Insulin-like growth factor binding protein-2 gene expression can be regulated by diet manipulation in several tissues of young chickens. J. Nutr. 2002, 132, 145–151. [Google Scholar] [CrossRef]
- Del Vesco, A.P.; Gasparino, E.; Oliveira, N.A.R.; Guimarães, S.E.; Marcato, S.M. Dietary methionine effects on IGF-I and GHR mRNA expression in broilers. Genet. Mol. Res. 2013, 12, 6414–6423. [Google Scholar] [CrossRef]
- Huang, S. Upregulation of TLR4 mRNA Expression Levels in Broiler Chickens Under Acute Heat Stress. Braz. J. Poult. Sci. 2017, 19, 87–94. [Google Scholar] [CrossRef] [Green Version]
- López-Maury, L.; Marguerat, S.; Bähler, J. Tuning gene expression to changing environments: From rapid responses to evolutionary adaptation. Nat. Rev. Genet. 2008, 9, 583–593. [Google Scholar] [CrossRef]
- Zhang, L.J.; Wang, K.F.; Jing, Y.P.; Zhuang, H.M.; Wu, G. Identification of heat shock protein genes HSP70s and HSP70 and their associated mRNA expression under heat stress in insecticide-resistant and susceptible diamondback moth, plutella xylostella (Lepidoptera: Plutellidae). Eur. J. Entomol. 2015, 112, 215–226. [Google Scholar] [CrossRef]
- Ju, X.H.; Xu, H.J.; Yong, Y.H.; An, L.L.; Jiao, P.R.; Liao, M. Heat stress upregulation of toll-like receptors 2/4 and acute inflammatory cytokines in peripheral blood mononuclear cell (PBMC) of bama miniature pigs: An in vivo and in vitro study. Animal 2014, 8, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Michelsen, K.S.; Eri, R.; Thomas, L.S.; Hu, B.; Lukasek, K. Toll like receptor-4 is required for intestinal response to epithelial injury and limiting bacterial translocation in a murine model of acute colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, 423–429. [Google Scholar] [CrossRef]
- Leon, L.R.; Helwig, B.G. Heat stroke: Role of the systemic inflammatory response. J. Appl. Physiol. 2010, 109, 1980–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanya, M.; Yamauchi, K.-E. Histological alterations of intestinal villi in chickens fed dried bacillus subtilis var. Natto. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 95–104. [Google Scholar] [CrossRef]
- Xu, Z.; Hu, C.; Xia, M.; Zhan, X.; Wang, M. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult. Sci. 2003, 82, 1030–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, T.; Tako, E. The in ovo feeding administration (Gallus gallus)—An emerging in vivo approach to assess bioactive compounds with potential nutritional benefits. Nutrients 2018, 10, 418. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Items | Treatments | SEM (Standard Error of the Mean) | p-Value | ||
|---|---|---|---|---|---|
| Control | NaCl | Met-Cys | |||
| Hatchability (%) | 78 | 78 | 80 | 1.3 | 0.758 |
| Hatch time (h) | 493 b | 508 a | 505 a | 1.6 | <0.01 |
| Chick weight (g) | 42.12 b | 42.67 b | 44.66 a | 0.38 | 0.027 |
| Liver (%) | 1.75 | 1.76 | 1.80 | 0.03 | 0.859 |
| Heart (%) | 0.57 | 0.57 | 0.58 | 0.01 | 0.991 |
| Pectoral muscle (%) | 0.71 | 0.72 | 0.81 | 0.02 | 0.146 |
| Small Intestine (%) | 1.97 | 1.99 | 2.07 | 0.06 | 0.779 |
| Left lung (%) | 0.30 | 0.33 | 0.32 | 0.01 | 0.449 |
| Residual yolk (%) | 14.24 | 14.56 | 14.40 | 0.44 | 0.956 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Control | NaCl | Met-Cys | |||
| Total Protein (g/L) | 16.97 | 14.92 | 16.64 | 0.409 | 0.124 |
| Albumin (g/L) | 9.21 | 8.88 | 9.66 | 0.479 | 0.804 |
| Globulin (g/L) | 7.76 | 6.04 | 6.98 | 0.391 | 0.231 |
| Albumin/Globulin ratio | 1.25 | 1.74 | 1.41 | 0.172 | 0.506 |
| Uric acid (µmol/L) | 46.54 b | 46.27 b | 50.54 a | 1.005 | 0.002 |
| Creatine kinase (mmol/L) | 1.28 | 1.19 | 1.28 | 1.253 | 0.786 |
| Triiodothyronine (ng/mL) | 1.09 b | 1.10 b | 1.20 a | 0.013 | 0.007 |
| Thyroxin (ng/mL) | 58.91 b | 52.30 b | 72.18 a | 1.609 | <0.001 |
| Heat shock protein 90 (ng/mL) | 6.54a | 6.52a | 5.24 b | 0.125 | 0.008 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Control | NaCl | Met-Cys | |||
| Serum | |||||
| TAOC (U/mL) | 8.50 b | 8.38 b | 11.13 a | 0.370 | 0.023 |
| GSH (µmol/L) | 23.54 b | 23.45 b | 36.93 a | 0.579 | <0.001 |
| Heart | |||||
| TAOC (U/mg protein) | 1.51 b | 1.65 b | 2.37 a | 0.051 | <0.001 |
| GSH (µmol/g protein) | 12.71 b | 13.39 b | 16.17 a | 0.264 | <0.001 |
| Pectoral muscle | |||||
| TAOC (U/mg protein) | 4.19 b | 4.55 b | 5.80 a | 0.214 | 0.017 |
| GSH (µmol/g protein) | 13.36 b | 12.46 b | 16.08 a | 0.206 | <0.001 |
| Small intestine | |||||
| TAOC (U/mg protein) | 9.27 b | 8.88 b | 10.37 a | 0.175 | 0.009 |
| GSH (µmol/g protein) | 19.58 b | 19.69 b | 21.16 a | 0.214 | 0.015 |
| Liver | |||||
| TAOC (U/mg protein) | 6.24 b | 6.06 b | 7.36 a | 0.181 | 0.018 |
| GSH (µmol/g protein) | 13.81 b | 13.68 b | 15.26 a | 0.183 | 0.005 |
| Kidney | |||||
| TAOC (U/mg protein) | 12.06 b | 12.24 b | 13.63 a | 0.238 | 0.028 |
| GSH (µmol/g protein) | 10.01 b | 9.01 b | 12.73 a | 0.359 | 0.001 |
| Items | Treatments | SEM | p-Value | ||
|---|---|---|---|---|---|
| Control | NaCl | Met-Cys | |||
| Villus height (µm) | 298.6 b | 314.9 b | 335.8 a | 9.14 | <0.001 |
| Villus width (µm) | 53.0 b | 52.8 b | 60.8 a | 2.29 | 0.002 |
| Villus area (µm) | 24,846.5 b | 26,104.0 b | 32,054.1 a | 958.27 | <0.001 |
| Crypt depth (µm) | 44.9 b | 44.9 b | 49.7 a | 0.90 | <0.001 |
| Crypt width (µm) | 25.7 | 24.5 | 22.8 | 0.94 | 0.199 |
| #HASA | 10.8 c | 11.7 b | 12.2 a | 0.34 | 0.001 |
| JMT * (µm) | 124.6 b | 127.5 b | 138.0 a | 3.41 | <0.001 |
© 2019 by the authors. Submitted for possible open access publication under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elwan, H.A.M.; Elnesr, S.S.; Xu, Q.; Xie, C.; Dong, X.; Zou, X. Effects of In Ovo Methionine-Cysteine Injection on Embryonic Development, Antioxidant Status, IGF-I and TLR4 Gene Expression, and Jejunum Histomorphometry in Newly Hatched Broiler Chicks Exposed to Heat Stress during Incubation. Animals 2019, 9, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9010025
Elwan HAM, Elnesr SS, Xu Q, Xie C, Dong X, Zou X. Effects of In Ovo Methionine-Cysteine Injection on Embryonic Development, Antioxidant Status, IGF-I and TLR4 Gene Expression, and Jejunum Histomorphometry in Newly Hatched Broiler Chicks Exposed to Heat Stress during Incubation. Animals. 2019; 9(1):25. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9010025
Chicago/Turabian StyleElwan, Hamada A. M., Shaaban S. Elnesr, Qianqian Xu, Chao Xie, Xinyang Dong, and Xiaoting Zou. 2019. "Effects of In Ovo Methionine-Cysteine Injection on Embryonic Development, Antioxidant Status, IGF-I and TLR4 Gene Expression, and Jejunum Histomorphometry in Newly Hatched Broiler Chicks Exposed to Heat Stress during Incubation" Animals 9, no. 1: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9010025