Interactions between Invasive Monk Parakeets (Myiopsitta monachus) and Other Bird Species during Nesting Seasons in Santiago, Chile

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Species
2.3. Study Methods
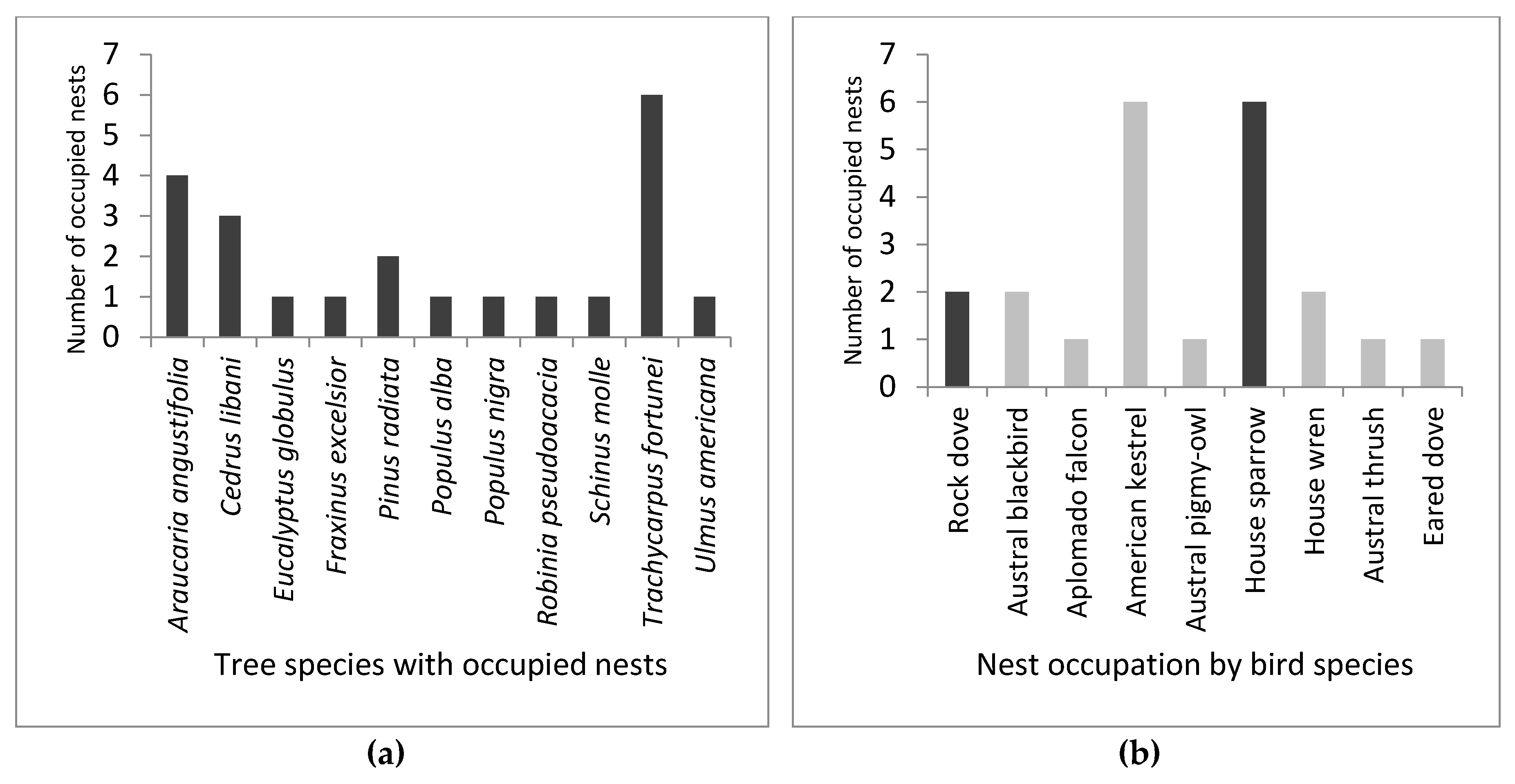
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Edelaar, P.; Roques, S.; Hobson, E.A.; Gonçalves da Silva, A.; Avery, M.L.; Russello, M.A.; Senar, J.C.; Wright, T.F.; Carrete, M.; Tella, J.L. Shared genetic diversity across the global invasive range of the monk parakeet suggests a common restricted geographic origin and the possibility of convergent selection. Mol. Ecol. 2015, 24, 2164–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olrog, C. Las Aves Sudamericanas. Una Guía De Campo. Torno L.; Fundación Instituto M. Lillo. Universidad Nacional de Tucumán: San Miguel de Tucumán, Argentina, 1968. [Google Scholar]
- ISSG Myiopsita monachus (monk parakeet). Global Invasive Species Database (GISD). Available online: http://www.issg.org/database (accessed on 5 May 2019).
- Batllori, X.; Nos, R. Presencia de la Cotorrita gris (Myiopsitta monachus) y de la Cotorrita de collar (Psittacula krameri) en el area metropolitana de Barcelona. Misc. Zool. 1985, 9, 407–411. [Google Scholar]
- Bucher, E.H.; Aramburú, R.M. Land-use changes and monk parakeet expansion in the pampas grasslands of argentina. J. Biogeogr. 2014, 41, 1160–1170. [Google Scholar] [CrossRef]
- Van Ham, C.; Genovesi, P.; Scalera, R. Invasive Alien Species: The Urban Dimension-Case Studies on Strengthening Local Action in Europe; IUCN: Brussels, Belgium, 2013; ISBN 9782831716251. [Google Scholar]
- Avery, M.L.; Tillman, E.A.; Keacher, K.L.; Arnett, J.E.; Lundy, K.J. Biology of Invasive Monk Parakeets in South Florida. Wilson J. Ornithol. 2012, 124, 581–588. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M. Exotic monk parakeets (Myiopsitta monachus) in New Jersey: Nest site selection, rebuilding following removal, and their urban wildlife appeal. Urban. Ecosyst. 2009, 12, 185–196. [Google Scholar] [CrossRef]
- Reed, J.E.; McCleery, R.A.; Silvy, N.J.; Smeins, F.E.; Brightsmith, D.J. Monk parakeet nest-site selection of electric utility structures in Texas. Landsc. Urban. Plan. 2014, 129, 65–72. [Google Scholar] [CrossRef]
- Domènech, J.; Carrillo, J.; Senar, J.C. Population size of the Monk Parakeet (Myiopsitta monachus) in Catalonia. Rev. Catalana d’Ornitologia 2003, 20, 1–9. [Google Scholar]
- Rodríguez-Pastor, R.; Señar, J.C.; Ortega, A.; Faus, J.; Uribe, F.; Montalvo, T. Distribution patterns of invasive Monk parakeets (Myiopsitta monachus) in an urban habitat. Anim. Biodivers. Conserv. 2012, 35, 107–117. [Google Scholar]
- Di Santo, M.; Battisti, C.; Bologna, M.A. Interspecific interactions in nesting and feeding urban sites among introduced Monk Parakeet (Myiopsitta monachus) and syntopic bird species. Ethol. Ecol. Evol. 2017, 29, 138–148. [Google Scholar] [CrossRef]
- Iriarte, A.J.; Lobos, G.A.; Jaksic, F.M. Invasive vertebrate species in Chile and their control and monitoring by governmental agencies. Rev. Chil. Hist. Nat. 2005, 78, 143–154. [Google Scholar]
- Senar, J.C.; Domènech, J.; Arroyo, L.; Torre, I.; Gordo, D. An evaluation of monk parakeet damage to crops in the metropolitan area of Barcelona. Anim. Biodivers. Conserv. 2019, 39, 141–145. [Google Scholar] [CrossRef]
- Briceño, C.; Surot, D.; González-Acuña, D.; Martínez, F.J.; Fredes, F. Parasitic survey on introduced monk parakeets (Myiopsitta monachus) in Santiago, Chile. Rev. Bras. Parasitol. Vet. 2017, 2961, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Tala, C.; Guzmán, P.; González, S. Cotorra Argentina (Myiopsitta monachus) Convidado de Piedra en Nuestras Ciudades Y un Invasor Potencial, Aunque Real, De Sectores Agrícolas; DIPROREN, Servicio Agrícola y Ganadero: Santiago, Chile, 2005. [Google Scholar]
- Davis, A.Y.; Malas, N.; Minor, E.S. Substitutable habitats? The biophysical and anthropogenic drivers of an exotic bird’s distribution. Biol. Invasions 2014, 16, 415–427. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Costa, A.K.F.; Sidrim, J.J.C.; Cordeiro, R.A.; Brilhante, R.S.N.; Monteiro, A.J.; Rocha, M.F.G. Urban pigeons (Columba livia) as a potential source of pathogenic yeasts: A focus on antifungal susceptibility of Cryptococcus strains in Northeast Brazil. Mycopathologia 2010, 169, 207–213. [Google Scholar] [CrossRef]
- Haag-Wackernagel, D.; Moch, H. Health hazards posed by feral pigeons. J. Infect. 2004, 48, 307–313. [Google Scholar] [CrossRef]
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife-threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate warming and disease risks for terrestrial and marine biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef]
- Smith, K.F.; Acevedo-Whitehouse, K.; Pedersen, A.B. The role of infectious diseases in biological conservation. Anim. Conserv. 2009, 12, 1–12. [Google Scholar] [CrossRef]
- Keesing, F.; Belden, L.K.; Daszak, P.; Dobson, A.; Harvell, C.D.; Holt, R.D.; Hudson, P.; Jolles, A.; Jones, K.E.; Mitchell, C.E.; et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature 2010, 468, 647–652. [Google Scholar] [CrossRef]
- Dunn, A.M.; Hatcher, M.J. Parasites and biological invasions: Parallels, interactions, and control. Trends Parasitol. 2015, 31, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Freeland, D.B. Some food preferences and aggressive behavior by monk parakeets. Wilson Bull. 1973, 85, 332–334. [Google Scholar]
- Davis, T.R. The monk parakeet: A potential threat to agriculture. In Proceedings of the 6th Vertebrate Pest Conference, Anaheim, CA, USA, 5–7 March 1974; pp. 253–256. [Google Scholar]
- Port, J.L.; Brewer, G.L. Use of Monk Parakeet (Myiopsitta monachus) nests by Speckled Teal (Anas flavirostris) in eastern Argentina. Ornitol. Neotrop. 2004, 15, 209–218. [Google Scholar]
- Nores, M. Use of Active Monk Parakeet Nests by Common Pigeons and Response by the Host. Wilson J. Ornithol. 2009, 121, 812–815. [Google Scholar] [CrossRef]
- Martella, M.B.; Navarro, J.L.; Bucher, E.H. Vertebrados asociados a los nidos de la cotorra argentina Myiopsitta monachus en Córdoba y La Rioja. Physis 1985, 43, 49–51. [Google Scholar]
- Lucca, D. Nidificación del Halconcito Colorado (Falco sparverius) en nidos de Cotorra (Myiopsitta monachus). Hornero Rev. Ornitol. Neotrop. 1992, 13, 238–240. [Google Scholar]
- Smith, P.; Fretes, A. Two unrelated species of birds (Myiopsitta monachus; Psittacidae and Coryphistera alaudina; Furnariidae) simultaneously building the same nest in the Paraguayan Chaco. Ecol. en Boliv. 2013, 48, 52–56. [Google Scholar]
- Wagner, N. Occupation of monk parakeet (Myiopsitta monachus) nest cavities by house sparrows (Passer domesticus) in Rio Grande do Sul, Brazil. Boletín SAO 2009, 20, 72–78. [Google Scholar]
- Celis-Diez, J.L. Observations on American Kestrels (Falco sparverius) predating on Monk Parakeet chicks (Myopsitta monachus) at urban parks in Santiago. Boletín Chil. Ornitol. 2014, 20, 23–24. [Google Scholar]
- Celis-Diez, J.L.; Muñoz, C.E.; Abades, S.; Marquet, P.A.; Armesto, J.J. Biocultural homogenization in Urban settings: Public knowledge of birds in city parks of Santiago, Chile. Sustainability 2017, 9, 485. [Google Scholar] [CrossRef]
- Jaksic, F.M. Vertebrate invaders and their ecological impacts in Chile. Biodivers. Conserv. 1998, 7, 1427–1445. [Google Scholar] [CrossRef]
- Amigo, J.; Ramírez, C. A Bioclimatic Classification of Chile: Woodland Communities in the Temperate Zone. Plant Ecol. 1998, 136, 9–26. [Google Scholar] [CrossRef]
- Schulz, J.J.; Cayuela, L.; Echeverria, C.; Salas, J.; Rey Benayas, J.M. Monitoring land cover change of the dryland forest landscape of Central Chile (1975–2008). Appl. Geogr. 2010, 30, 436–447. [Google Scholar] [CrossRef]
- Badano, E.I.; Cavieres, L.A.; Molina-Montenegro, M.A.; Quiroz, C.L. Slope aspect influences plant association patterns in the Mediterranean matorral of central Chile. J. Arid Environ. 2005, 62, 93–108. [Google Scholar] [CrossRef]
- INE. Censo de población y vivienda de Chile año 2017; INE: Santiago, Chile, 2017; Available online: http://resultados.censo2017.cl/ (accessed on 20 October 2019).
- Vogiatzakis, I.N.; Mannion, A.M.; Griffiths, G.H. Mediterranean ecosystems: Problems and tools for conservation. Prog. Phys. Geogr. 2006, 30, 175–200. [Google Scholar] [CrossRef]
- Silva Lovera, C.A. Urban sprawl and infrastructural lands: Revamping internal spaces in Santiago de Chile. Geoforum 2015, 67, 36–40. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Eberhard, J.R. Breeding biology of the monk parakeet. Wilson Bull. 1998, 110, 463–473. [Google Scholar]
- MacGregor-Fors, I.; Calserón-Parra, R.; Meléndez-Herrada, A.; López-López, S.; Schondube, J.E. Pretty, but dangerous! Records of non-native Monk Parakeets (Myiopsitta monachus) in Mexico. Rev. Mex. Biodivers. 2011, 81, 1053–1056. [Google Scholar]
- Martin, L.F.; Bucher, E.H. Natal dispersal and first breeding age in monk parakeets. Auk 1993, 110, 930–933. [Google Scholar]
- Strubbe, D.; Matthysen, E. Establishment success of invasive ring-necked and monk parakeets in Europe. J. Biogeogr. 2009, 36, 2264–2278. [Google Scholar] [CrossRef]
- Senar, J.C.; Carrillo-Ortiz, J.G.; Ortega-Segalerva, A.; Dawson Pell, F.S.E.; Pascual, J.; Arroyo, L.; Mazzoni, D.; Montalvo, T.; Hatchwell, B.J. The reproductive capacity of Monk Parakeets Myiopsitta monachus is higher in their invasive range. Bird Study 2019, 66, 136–140. [Google Scholar] [CrossRef]
- Bucher, E.; Martin, L.; Martella, M.; Navarro, J. Social behaviour and population dynamics of the Monk Parakeet. Acta XX Congr. Int. 1991, 2, 681–690. [Google Scholar]
- Spreyer, M.F.; Bucher, E.H. Monk parakeet. In The Birds of North America; Poole, A., Gil, F., Eds.; Cornell Lab of Ornithology: Philadelphia, PA, USA, 1998; pp. 1–23. [Google Scholar]
- Navarro, J.L.; Martella, M.B.; Bucher, E.H.; The, S.; Bulletin, W.; Dec, N. Effects of laying date, clutch size, and communal nest size on the reproductive success of monk parakeets. Wilson Bull. 1995, 107, 742–746. [Google Scholar]
- Navarro, J.L.; Martella, M.B.; Bucher, E.H. Breeding Season and Productivity of Monk Parakeets in Cordoba, Argentina. Wilson Bull. 1992, 104, 413–424. [Google Scholar]
- Aramburú, R.M. Descripción y desarrollo del pichón de la cotorra Myiopsitta monachus monachus (Aves: Psittacidae) en una población silvestre de Argentina. Rev. Chil. Hist. Nat. 1997, 70, 53–58. [Google Scholar]
- Gonçalves da Silva, A.; Eberhard, J.R.; Wright, T.F.; Avery, M.L.; Russello, M.A.; da Silva, A.G.; Eberhard, J.R.; Wright, T.F.; Avery, M.L.; Russello, M.A. Genetic evidence for high propagule pressure and long-distance dispersal in monk parakeet (Myiopsitta monachus) invasive populations. Mol. Ecol. 2010, 19, 3336–3350. [Google Scholar] [CrossRef]
- De Azevedo, C.S.; Rodrigues, L.S.F.; Fontenelle, J.C.R. Important tools for Amazon Parrot reintroduction programs. Rev. Bras. Ornitol. 2017, 25, 1–11. [Google Scholar]
- Mcgarry, J. A proposal for a standard ethogram, exemplified by an ethogram of the bluebrested quail (Coturnix chinensis). Ethology 1984, 64, 193–220. [Google Scholar]
- Brown, E.D.; Veltman, C.J. Ethogram of the Australian Magpie (Gymnorhina tibicen) in Comparison to Other Cracticidae and Corvus Species. Ethology 1987, 76, 309–333. [Google Scholar] [CrossRef]
- Seibert, L.M. Social behavior of psittacine birds. In Manual of Parrot Behavior; Luescher, A.U., Ed.; Blackwell Publishing: London, UK, 2006; pp. 43–48. [Google Scholar]
- Burgiel, S.W.; Muir, A.A. Invasive Species, Climate Change and Ecosystem-Based Adaptation: Addressing Multiple Drivers of Global Change; Global Invasive Species Program (GISP): Washington DC, USA, 2010; ISBN 9789290592877. [Google Scholar]
- Levin, R.N.; Wingfield, J.C.; Levin, R.N.; Wingfield, J.C. The hormonal control of territorial aggression in tropical birds. Ornis Scand. 1991, 23, 284–291. [Google Scholar] [CrossRef]
- South, J.; Pruett-Jones, S. Patterns of Flock Size, Diet, and Vigilance of Naturalized Monk Parakeets in Hyde Park, Chicago. Condor 2000, 102, 848. [Google Scholar] [CrossRef]
- Humphrey, P.S.; Peterson, R.T. Nesting behavior and affinities of Monk Parakeets of Southern Buenos Aires province, Argentina. Wilson Bull. 1978, 90, 544–552. [Google Scholar]
- Hilton, G.M.; Ruxton, G.D.; Reid, J.M.; Monaghan, P. Using Artificial Nests to Test Importance of Nesting Material and Nest Shelter for Incubation Energetics. Auk 2004, 121, 777–787. [Google Scholar] [CrossRef]
- Viana, I.R.; Strubbe, D.; Zocche, J.J. Monk parakeet invasion success: A role for nest thermoregulation and bactericidal potential of plant nest material? Biol. Invasions 2016, 18, 1305–1315. [Google Scholar] [CrossRef]
- Tuero, D.T.; Fiorini, V.D.; Reboreda, J.C. Effects of Shiny Cowbird Molothrus bonariensis parasitism on different components of House Wren Troglodytes aedon reproductive success. Ibis 2007, 149, 521–529. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Positive and Negative Effects of Organisms As Physical Ecosystem Engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Chapman, C.A.; Bonnell, T.R.; Gogarten, J.F.; Lambert, J.E.; Omeja, P.A.; Twinomugisha, D.; Wasserman, M.D.; Rothman, J.M. Are Primates Ecosystem Engineers? Int. J. Primatol. 2013, 34, 1–14. [Google Scholar] [CrossRef]
- Crooks, J.A. Characterizing ecosystem-level consequences of biological invasions: The role of ecosystem engineers. Oikos 2002, 97, 153–166. [Google Scholar] [CrossRef]
- López-García, R. Personal Communication; Scrofa S.L.: Madrid, Spain, 2019. [Google Scholar]
- Myczko, Ł.; Dylewski, Ł.; Sparks, T.H.; Łochyński, M.; Tryjanowski, P. Co-occurrence of birds and bats in natural nest-holes. Ibis 2017, 159, 235–237. [Google Scholar] [CrossRef]
- Aramburú, R.; Calvo, S.; Alzugaray, M.; Cicchino, A. Ectoparasitic load of monk parakeet (Myiopsitta monachus, Psittacidae) nestlings. Ornitol. Neotrop. 2003, 14, 415–418. [Google Scholar]
- Bucher, E.H. Do birds use biological control against nest parasites? Parasitol. Today 1988, 4, 1–3. [Google Scholar] [CrossRef]
- Chiffelle, I.; Huerta Fuentes, A.; Lizana Rojas, D. Physical and Chemical Characterization of Melia azedarach L. Fruit and Leaf for Use as Botanical Insecticide. Chil. J. Agric. Res. 2009, 69, 38–45. [Google Scholar] [CrossRef]
- Chiffelle, I.; Huerta, A.; Puga, K.; Araya, J.E. Antifeeding and Insecticide Properties of Aqueous and ethanolic fruit extracts from Melia azedarach L. on the elm leaf beetle Xanthogaleruca luteola Müller. Chil. J. Agric. Res. 2011, 71, 218. [Google Scholar] [CrossRef]
- Munita, C.; Tadich, T.; Briceño, C. Comparison of two behavioral sampling methods to establish a time budget in a captive female cheetah (Acinonyx jubatus). J. Vet. Behav. Clin. Appl. Res. 2016, 13, 1–5. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Tree Species | Scientific Name | Frequency of Observation (%) |
|---|---|---|
| Paraná pine | Araucaria angustifolia | 33 (25.4) |
| Cedar of Lebanon | Cedrus libani | 18 (13.8) |
| Chinese windmill palm | Trachycarpus fortunei | 18 (13.8) |
| Black locust | Robinia pseudoacacia | 11 (8.5) |
| Tasmanian bluegum | Eucalyptus globulus | 9 (6.9) |
| Monterey pine | Pinus radiata | 6 (4.6) |
| American pepper | Schinus molle | 6 (4.6) |
| Black poplar | Populus nigra | 5 (3.8) |
| Common ash | Fraxinus excelsior | 3 (2.3) |
| Canary palm | Phoenix canariensis | 3 (2.3) |
| Bunya pine | Araucaria bidwillii | 2 (1.5) |
| American elm | Ulmus americana | 2 (1.5) |
| Desert fan palm | Washingtonia filifera | 2 (1.5) |
| Box elder | Acer negundo | 1 (0.8) |
| Norfolk Island pine | Araucaria heterophyla | 1 (0.8) |
| Sweet chestnut | Castanea sativa | 1 (0.8) |
| Atlas cedar | Cedrus atlantica | 1 (0.8) |
| Chilean acorn | Cryptocarya alba | 1 (0.8) |
| Monterey cypress | Cupressus macrocarpa | 1 (0.8) |
| Ceibo | Erythrina umbrosa | 1 (0.8) |
| Chilean wine palm | Jubea chilensis | 1 (0.8) |
| Sweet gum | Liquidambar styraciflua | 1 (0.8) |
| White poplar | Populus alba | 1 (0.8) |
| Water oak | Quercus nigra | 1 (0.8) |
| Weeping willow | Salix babylonica | 1 (0.8) |
| Total | 130 (100) |
| Bird Species | Tree Species | Observation (No. Records) |
|---|---|---|
| Tufted tit-tyrant (Anairetes parulus) | Black locust | Occupying same tree (1) |
| Rock dove (Columba livia) 1 | Box elder; Paraná pine; Bunya pine; Norfolk Island pine; Sweet chestnut; Cedar of Lebanon 2; Monterey cypress; Tasmanian bluegum; Canary palm; Black locust; American pepper; Chinese windmill palm 2; American elm; Desert fan palm | Occupying same tree (30) Occupying nest (2) Forage together (1) |
| Austral blackbird (Curaeus curaeus) | Paraná pine 2; Cedar of Lebanon; Ceibo; Black poplar; Chinese windmill palm | Occupying same tree (14) Occupying nest (2) Attacks (1) |
| Common diuca finch (Diuca diuca) | Paraná pine | Occupying same tree (1) |
| Aplomado falcon (Falco femoralis) | Tasmanian bluegum 2 | Occupying nest (1) |
| American kestrel (Falco sparverius) | Paraná pine 2; Monterey pine 2; White poplar 2; Black locust; American pepper 2; American elm 2 | Occupying same tree (7) Occupying nest (6) |
| Austral pigmy-owl (Glaucidum nanun) | Common ash 2 | Occupying nest (1) |
| Chimango caracara (Milvago chimango) | Box elder; Paraná pine; Bunya pine; Cedar of Lebanon; Tasmanian bluegum; Chilean wine pine; Monterey pine; White poplar; Black locust; Weeping willow; American pepper; American elm | Occupying same tree (18) Attacks (12) |
| Chilean mockingbird (Mimus thenca) | Cedar of Lebanon; Common ash; Black poplar; American pepper | Occupying same tree (4) |
| Shiny cowbird (Molothrous bonariensis) 1 | Cedar of Lebanon; Tasmanian bluegum; Chilean wine palm; Monterey pine | Occupying same tree (5) |
| Harri’s hawk (Parabuteo unicinctus) | Water oak | Attacks (1) Predates (1) |
| House sparrow (Passer domesticus) 1 | Paraná pine; Common ash; Sweet gum; Canary palm; Monterey pine 2; Black poplar 2; Chinese windmill palm 2 | Occupying same tree (11) Occupying nest (6) |
| Rufous-tailed plantcutter (Phytotoma rara) | Ceibo | Occupying same tree (1) |
| Green-backed firecrown (Sephanoides sephaniodes) | Black locust | Occupying same tree (2) |
| Chilean swallow (Tachycineta leucopyga) | Chilean wine palm | Occupying same tree (1) |
| House wren (Troglodytes aedon chilensis) | Cedar of Lebanon 2; Black locust 2; Chinese windmill palm | Occupying same tree (4) Occupying nest (2) |
| Austral thrush (Turdus falcklandii) | Paraná pine; Norfolk Island pine; Sweet chestnut; Atlas cedar; Cedar of Lebanon; Chilean acorn; Canary palm; Black locust; Chinese windmill palm 2; American elm | Occupying same tree (24) Occupying nest (1) Forage together (1) |
| Striped woodpecker (Veniliornis lignarius) | Monterey pine | Occupying same tree (2) |
| Eared dove (Zenaida auriculata) | Paraná pine; Cedar of Lebanon 2; Chilean acorn; Chilean wine palm; Black poplar; Black locust; American pepper; Chinese windmill palm | Occupying same tree (11) Occupying nest (1) Forage together (2) |
| Rufous-collared sparrow (Zonotrichia capensis chilensis) | Sweet gum; Desert fan palm | Occupying same tree (2) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briceño, C.; Sandoval-Rodríguez, A.; Yévenes, K.; Larraechea, M.; Morgado, A.; Chappuzeau, C.; Muñoz, V.; Dufflocq, P.; Olivares, F. Interactions between Invasive Monk Parakeets (Myiopsitta monachus) and Other Bird Species during Nesting Seasons in Santiago, Chile. Animals 2019, 9, 923. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9110923
Briceño C, Sandoval-Rodríguez A, Yévenes K, Larraechea M, Morgado A, Chappuzeau C, Muñoz V, Dufflocq P, Olivares F. Interactions between Invasive Monk Parakeets (Myiopsitta monachus) and Other Bird Species during Nesting Seasons in Santiago, Chile. Animals. 2019; 9(11):923. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9110923
Chicago/Turabian StyleBriceño, Cristóbal, Alejandra Sandoval-Rodríguez, Karina Yévenes, Matilde Larraechea, Angello Morgado, Catalina Chappuzeau, Víctor Muñoz, Pablo Dufflocq, and Florencia Olivares. 2019. "Interactions between Invasive Monk Parakeets (Myiopsitta monachus) and Other Bird Species during Nesting Seasons in Santiago, Chile" Animals 9, no. 11: 923. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9110923