Genome-Wide Association Study for Milk Fatty Acids in Holstein Cattle Accounting for the DGAT1 Gene Effect

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Datasets
2.2. Variance Components, Breeding Value Estimation and Deregression Analyses
2.3. Genome-Wide Association Analyses
2.4. Candidate Genes, Enrichment and Biology Pathway Analysis
3. Results
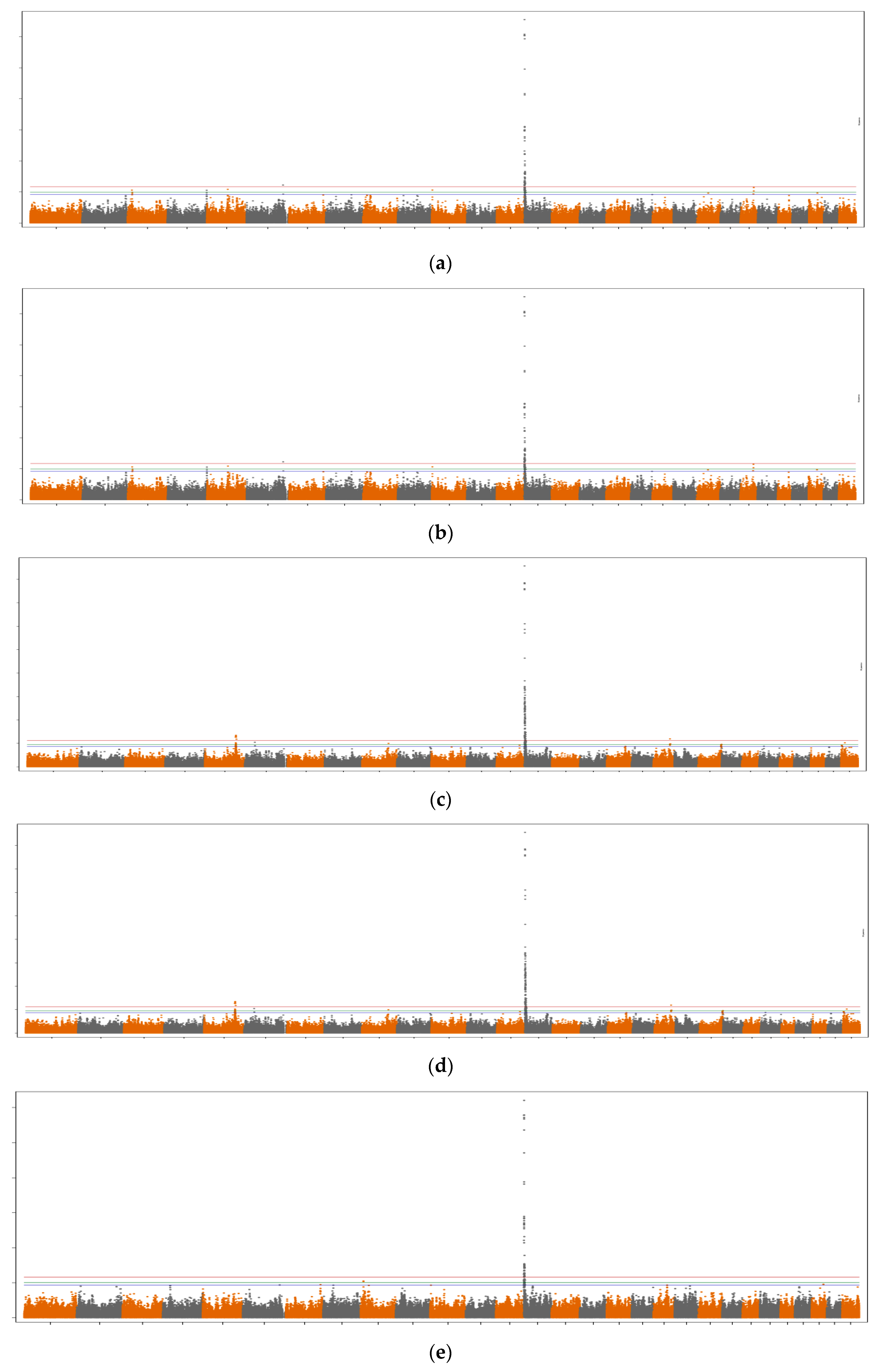
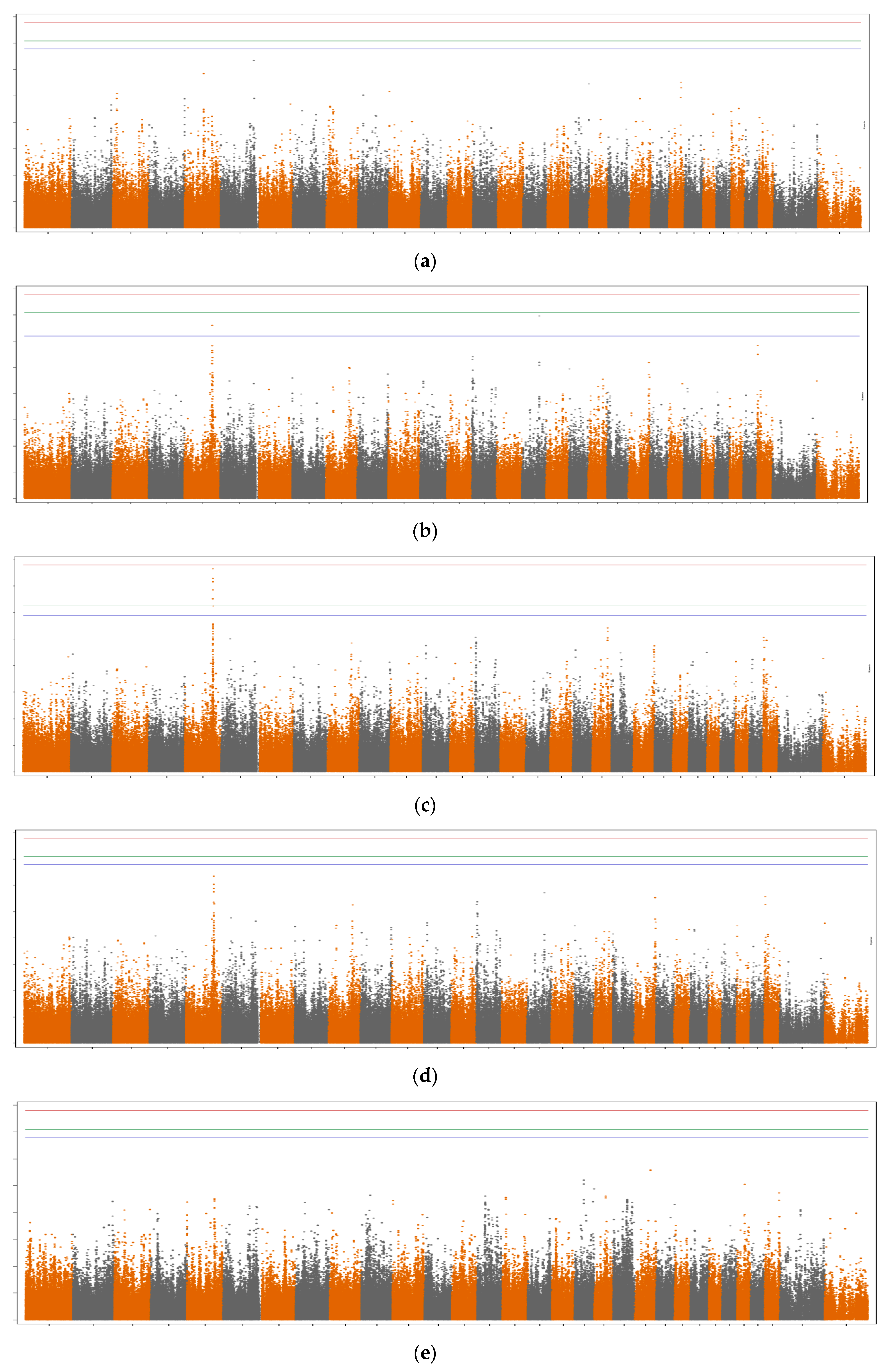
3.1. SNP Variances and Number of Significant SNPs
3.2. Genomic Regions and Candidate Genes
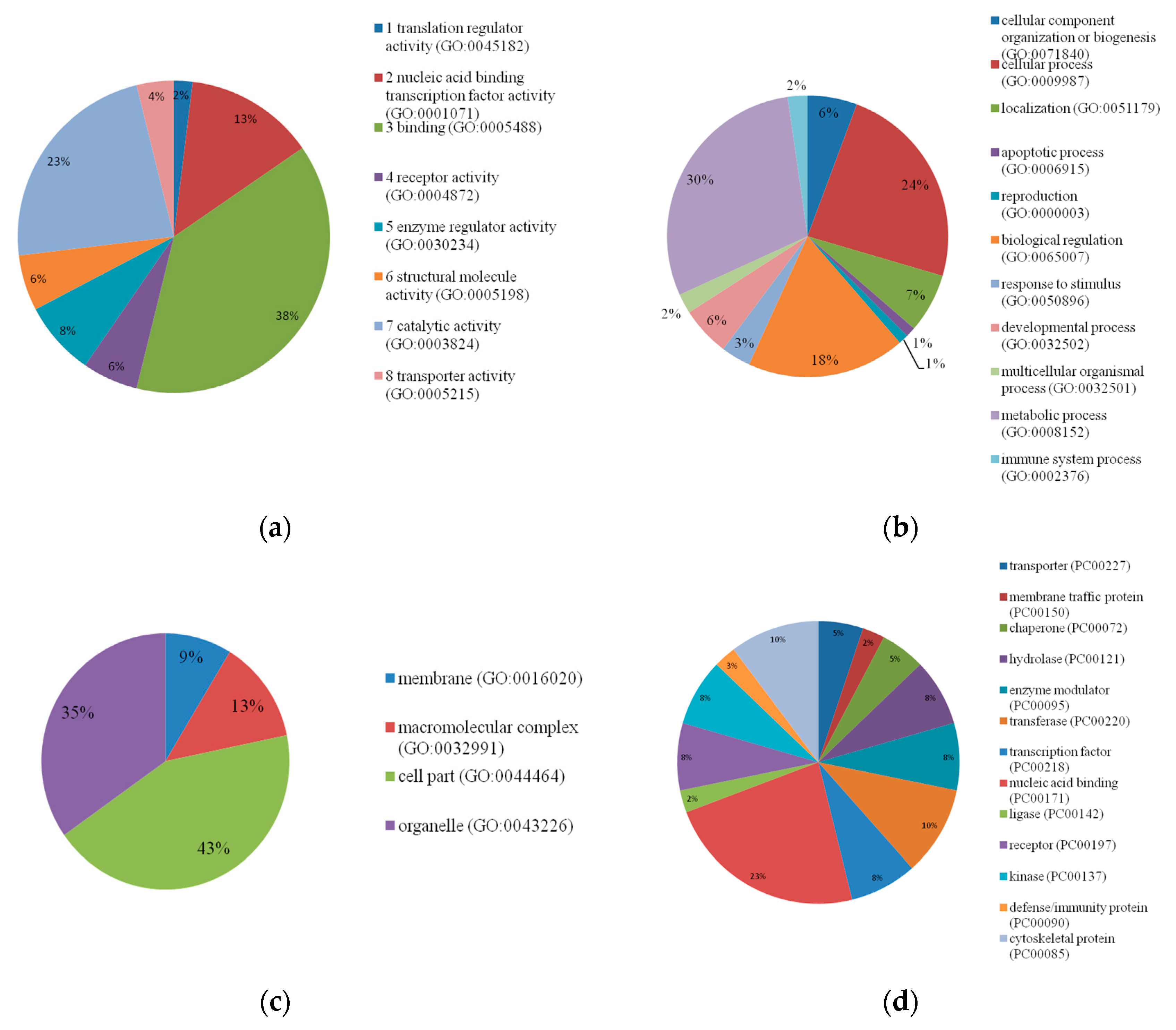
3.3. Candidate Genes and Functional Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability Statement
References
- Mansbridge, R.J.; Blake, J.S. Nutritional factors affecting the fatty acid composition of bovine milk. Br. J. Nutr. 1997, 78, S37–S47. [Google Scholar] [CrossRef]
- Brouwer, I.A.; Wanders, A.J.; Katan, M.B. Trans fatty acids and cardiovascular health: Research completed? Eur. J. Clin. Nutr. 2013, 67, 541. [Google Scholar] [CrossRef]
- Connor, W.E. Importance of n-3 fatty acids in health and disease. Am. J. Clin. Nutr. 2000, 71, 171S–175S. [Google Scholar] [CrossRef]
- FAO-WHO, Fats and Fatty Acids in Human Nutrition, Report of an Expert Consultation. FAO Food Nutrition Paper. # 91. 2008. Available online: http://www.fao.org/3/a-i1953e.pdf (accessed on 14 January 2019).
- Grummer, R.R. Effect of Feed on the Composition of Milk Fat. J. Dairy Sci. 1991, 74, 3244–3257. [Google Scholar] [CrossRef]
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Yorek, M.A. Membrane lipid composition and cellular function. J. Lipid Res. 1985, 26, 1015–1035. [Google Scholar] [PubMed]
- Palombo, V.; Milanesi, M.; Sgorlon, S.; Capomaccio, S.; Mele, M.; Nicolazzi, E.; Ajmone-Marsan, P.; Pilla, F.; Stefanon, B.; D’Andrea, M. Genome-wide association study of milk fatty acid composition in Italian Simmental and Italian Holstein cows using single nucleotide polymorphism arrays. J. Dairy Sci. 2018, 101, 11004–11019. [Google Scholar] [CrossRef]
- Pulina, G.; Francesconi, A.H.D.; Stefanon, B.; Sevi, A.; Calamari, L.; Lacetera, N.; Dell’Orto, V.; Pilla, F.; Marsan, P.A.; Mele, M.; et al. Sustainable ruminant production to help feed the planet. Ital. J. Anim. Sci. 2017, 16, 140–171. [Google Scholar] [CrossRef]
- Buitenhuis, B.; Janss, L.L.; Poulsen, N.A.; Larsen, L.B.; Larsen, M.K.; Sørensen, P. Genome-wide association and biological pathway analysis for milk-fat composition in Danish Holstein and Danish Jersey cattle. BMC Genom. 2014, 15, 1112. [Google Scholar] [CrossRef]
- Narayana, S.G.; Schenkel, F.S.; Fleming, A.; Koeck, A.; Malchiodi, F.; Jamrozik, J.; Johnston, J.; Sargolzaei, M.; Miglior, F. Genetic analysis of groups of mid-infrared predicted fatty acids in milk. J. Dairy Sci. 2017, 100, 4731–4744. [Google Scholar] [CrossRef]
- Tullo, E.; Frigo, E.; Rossoni, A.; Finocchiaro, R.; Serra, M.; Rizzi, N.; Samorè, A.B.; Canavesi, F.; Strillacci, M.G.; Maria Prinsen, R.T.M.; et al. Genetic parameters of fatty acids in Italian Brown Swiss and Holstein cows. Ital. J. Anim. Sci. 2014, 13, 3208. [Google Scholar] [CrossRef]
- Bouwman, A.C.; Visker, M.H.P.W.; van Arendonk, J.A.M.; Bovenhuis, H. Genomic regions associated with bovine milk fatty acids in both summer and winter milk samples. BMC Genet. 2012, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.; Goddard, M. Genome-wide association and genomic selection in animal breeding. Genome 2010, 53, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Marques, E.; Schnabel, R.D.; Stothard, P.; Kolbehdari, D.; Wang, Z.; Taylor, J.F.; Moore, S.S. High density linkage disequilibrium maps of chromosome 14 in Holstein and Angus cattle. BMC Genet. 2008, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Capomaccio, S.; Milanesi, M.; Bomba, L.; Cappelli, K.; Nicolazzi, E.L.; Williams, J.L.; Ajmone-Marsan, P.; Stefanon, B. Searching new signals for production traits through gene-based association analysis in three Italian cattle breeds. Anim. Genet. 2015, 46, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Fleming, A.; Schenkel, F.S.; Chen, J.; Malchiodi, F.; Bonfatti, V.; Ali, R.A.; Mallard, B.; Corredig, M.; Miglior, F. Prediction of milk fatty acid content with mid-infrared spectroscopy in Canadian dairy cattle using differently distributed model development sets. J. Dairy Sci. 2017, 100, 5073–5081. [Google Scholar] [CrossRef]
- Sargolzaei, M.; Chesnais, J.P.; Schenkel, F.S. FImpute—An efficient imputation algorithm for dairy cattle populations. J. Dairy Sci. 2011, 94, 421. [Google Scholar]
- Larmer, S.; Sargolzaei, M.; Schenkel, F. Extent of linkage disequilibrium, consistency of gametic phase, and imputation accuracy within and across Canadian dairy breeds. J. Dairy Sci. 2014, 97, 3128–3141. [Google Scholar] [CrossRef]
- Gilmour, A.R.; Gogel, B.J.; Cullis, B.R.; Thompson, R. ASReml User Guide Release 3.0; VSN Int. Ltd.: Hemel Hempstead, UK, 2009. [Google Scholar] [CrossRef]
- VanRaden, P.M.; Wiggans, G.R. Derivation, Calculation, and Use of National Animal Model Information. J. Dairy Sci. 1991, 74, 2737–2746. [Google Scholar] [CrossRef]
- Oliveira, H.R.; Silva, F.F.; Brito, L.F.; Guarini, A.R.; Jamrozik, J.; Schenkel, F.S. Comparing deregression methods for genomic prediction of test-day traits in dairy cattle. J. Anim. Breed. Genet. 2018, 135, 97–106. [Google Scholar] [CrossRef]
- Sargolzaei, M. SNP1101 User’s Guide. Version 1.0; HiggsGene Solutions Inc.: Guelph, ON, Canada, 2014. [Google Scholar]
- Gianola, D.; Fariello, M.I.; Naya, H.; Schön, C.C. Genome-wide association studies with a genomic relationship matrix: A case study with wheat and arabidopsis. G3: Genes, Genomes, Genet. 2016, 6, 3241–3256. [Google Scholar] [CrossRef] [PubMed]
- VanRaden, P.M. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Sun, D.; Zhang, S.; Wang, S.; Wu, X.; Zhang, Q.; Liu, L.; Li, Y.; Qiao, L. Genome wide association study identifies 20 novel promising genes associated with milk fatty acid traits in Chinese Holstein. PLoS ONE 2014, 9, e96186. [Google Scholar] [CrossRef] [PubMed]
- Nayeri, S.; Sargolzaei, M.; Abo-Ismail, M.K.; May, N.; Miller, S.P.; Schenkel, F.; Moore, S.S.; Stothard, P. Genome-wide association for milk production and female fertility traits in Canadian dairy Holstein cattle. BMC Genet. 2016, 17, 75. [Google Scholar] [CrossRef]
- Oliveira, H.R.; Cant, J.P.; Brito, L.F.; Feitosa, F.L.B.; Chud, T.C.S.; Fonseca, P.A.S.; Jamrozik, J.; Silva, F.F.; Lourenco, D.A.L.; Schenkel, F.S. Genome-wide association for milk production traits and somatic cell score in different lactation stages of Ayrshire, Holstein, and Jersey dairy cattle. J. Dairy Sci. 2019, 102, 8159–8174. [Google Scholar] [CrossRef]
- Oliveira, H.R.; Lourenco, D.A.L.; Masuda, Y.; Misztal, I.; Tsuruta, S.; Jamrozik, J.; Brito, L.F.; Silva, F.F.; Cant, J.P.; Schenkel, F.S. Single-step genome-wide association for longitudinal traits of Canadian Ayrshire, Holstein, and Jersey dairy cattle. J. Dairy Sci. 2019, 102, 9995–10011. [Google Scholar] [CrossRef]
- Kaupe, B.; Kollers, S.; Fries, R.; Erhardt, G. Mapping of CYP11B and a putative CYHR1 paralogous gene to bovine chromosome 14 by FISH. Anim. Genet. 2004, 35, 478–479. [Google Scholar] [CrossRef]
- Maryam, J.; Babar, M.E.; Nadeem, A.; Yaqub, T.; Hashmi, A.S. Identification of functional consequence of a novel selection signature in CYP11b1 gene for milk fat content in Bubalus bubalis. Meta Gene 2015, 6, 85–90. [Google Scholar] [CrossRef]
- Boleckova, J.; Matejickova, J.; Stipkova, M.; Kyselova, J.; Barton, L. The association of five polymorphisms with milk production traits in Czech Fleckvieh cattle. Czech. J. Anim. Sci. 2012, 57, 45–53. [Google Scholar] [CrossRef]
- Hayes, B.J.; Pryce, J.; Chamberlain, A.J.; Bowman, P.J.; Goddard, M.E. Genetic architecture of complex traits and accuracy of genomic Prediction: Coat colour, Milk-fat percentage, and type in Holstein cattle as contrasting model traits. PLoS Genet. 2010, 6, e1001139. [Google Scholar] [CrossRef]
- Moir, R.D.; Willis, I.M. Regulating Maf1 Expression and Its Expanding Biological Functions. PLoS Genet. 2015, 11, e1004896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palian, B.M.; Rohira, A.D.; Johnson, S.A.S.; He, L.; Zheng, N.; Dubeau, L.; Stiles, B.L.; Johnson, D.L. Maf1 Is a Novel Target of PTEN and PI3K Signaling That Negatively Regulates Oncogenesis and Lipid Metabolism. PLoS Genet. 2014, 14, e1004789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuchiya, M.; Yoshida, T.; Taniguchi, S.; Yasuda, K.; Maeda, A.; Hayashi, A.; Tanaka, J.; Shigemoto, M.; Nitta, K.; Tsuchiya, K. In vivo suppression of mafA mRNA with siRNA and analysis of the resulting alteration of the gene expression profile in mouse pancreas by the microarray method. Biochem. Biophys. Res. Commun. 2007, 356, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Morgenstern, R.; Zhang, J.; Johansson, K. Microsomal glutathione transferase 1: Mechanism and functional roles. Drug Metab. Rev. 2011, 43, 300–306. [Google Scholar] [CrossRef]
- Mosialou, E.; Morgenstern, R. Activity of rat liver microsomal glutathione transferase toward products of lipid peroxidation and studies of the effect of inhibitors on glutathione-dependent protection against lipid peroxidation. Arch. Biochem. Biophys. 1989, 275, 289–294. [Google Scholar] [CrossRef]
- Mosialou, E.; Ekström, G.; Adang, A.E.P.; Morgenstern, R. Evidence that rat liver microsomal glutathione transferase is responsible for glutathione-dependent protection against lipid peroxidation. Biochem. Pharmacol. 1993, 45, 1645–1651. [Google Scholar] [CrossRef]
- Mosialou, E.; Piemonte, F.; Andersson, C.; Vos, R.M.E.; van Bladeren, P.J.; Morgenstern, R. Microsomal glutathione transferase: Lipid-derived substrates and lipid dependence. Arch. Biochem. Biophys. 1995, 320, 210–216. [Google Scholar] [CrossRef]
- Repo, H.; Kuokkanen, E.; Oksanen, E.; Goldman, A.; Heikinheimo, P. Is the bovine lysosomal phospholipase B-like protein an amidase? Proteins Struct. Funct. Bioinform. 2014, 82, 300–311. [Google Scholar] [CrossRef]
- Buccioni, A.; Pauselli, M.; Viti, C.; Minieri, S.; Pallara, G.; Roscini, V.; Rapaccini, S.; Marinucci, M.T.; Lupi, P.; Conte, G.; et al. Milk fatty acid composition, rumen microbial population, and animal performances in response to diets rich in linoleic acid supplemented with chestnut or quebracho tannins in dairy ewes. J. Dairy Sci. 2015, 98, 1145–1156. [Google Scholar] [CrossRef] [Green Version]
- Kliem, K.E.; Shingfield, K.J. Manipulation of milk fatty acid composition in lactating cows: Opportunities and challenges. Eur. J. Lipid Sci. Technol. 2016, 118, 1661–1683. [Google Scholar] [CrossRef]
- Meignan, T.; Lechartier, C.; Chesneau, G.; Bareille, N. Effects of feeding extruded linseed on production performance and milk fatty acid profile in dairy cows: A meta-analysis. J. Dairy Sci. 2017, 100, 4394–4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Traits 1 | Not Fitting DGAT1 Gene Effect | Fitting DGAT1 Gene Effect | ||||
|---|---|---|---|---|---|---|
| 1% | 5% | 10% | 1% | 5% | 10% | |
| LCFA | 89 | 134 | 157 | 0 | 0 | 0 |
| MCFA | 204 | 252 | 338 | 0 | 0 | 3 |
| SCFA | 156 | 233 | 273 | 0 | 6 | 7 |
| SFA | 204 | 251 | 334 | 0 | 0 | 0 |
| UFA | 90 | 119 | 132 | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz, V.A.R.; Oliveira, H.R.; Brito, L.F.; Fleming, A.; Larmer, S.; Miglior, F.; Schenkel, F.S. Genome-Wide Association Study for Milk Fatty Acids in Holstein Cattle Accounting for the DGAT1 Gene Effect. Animals 2019, 9, 997. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9110997
Cruz VAR, Oliveira HR, Brito LF, Fleming A, Larmer S, Miglior F, Schenkel FS. Genome-Wide Association Study for Milk Fatty Acids in Holstein Cattle Accounting for the DGAT1 Gene Effect. Animals. 2019; 9(11):997. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9110997
Chicago/Turabian StyleCruz, Valdecy A. R., Hinayah R. Oliveira, Luiz F. Brito, Allison Fleming, Steven Larmer, Filippo Miglior, and Flavio S. Schenkel. 2019. "Genome-Wide Association Study for Milk Fatty Acids in Holstein Cattle Accounting for the DGAT1 Gene Effect" Animals 9, no. 11: 997. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9110997