Protective Effects of Fucoidan against Hydrogen Peroxide-Induced Oxidative Damage in Porcine Intestinal Epithelial Cells



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Establishment of Oxidative Stress
2.3. Fucoidan Treatment
2.4. Assay of Apoptosis and Necrosis
2.5. Measurement of Intracellular ROS
2.6. Determination of Antioxidant-Related Parameters
2.7. RNA Isolation and Reverse Transcription
2.8. Quantitative Real-Time PCR Analysis
2.9. Immunofluorescence Assay of NRF2
2.10. Statistical Analysis
3. Results
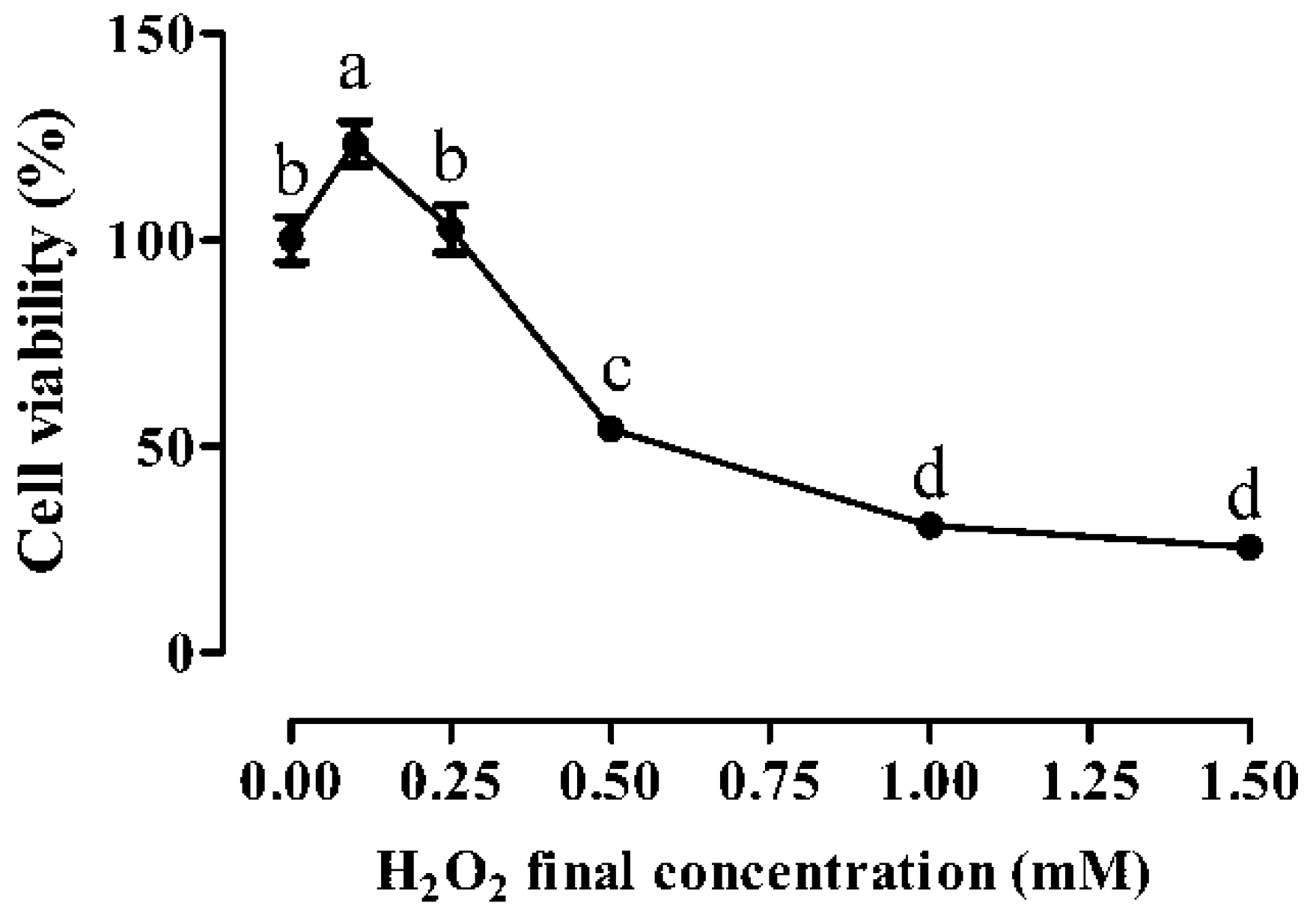
3.1. Cytotoxic Effects of H2O2
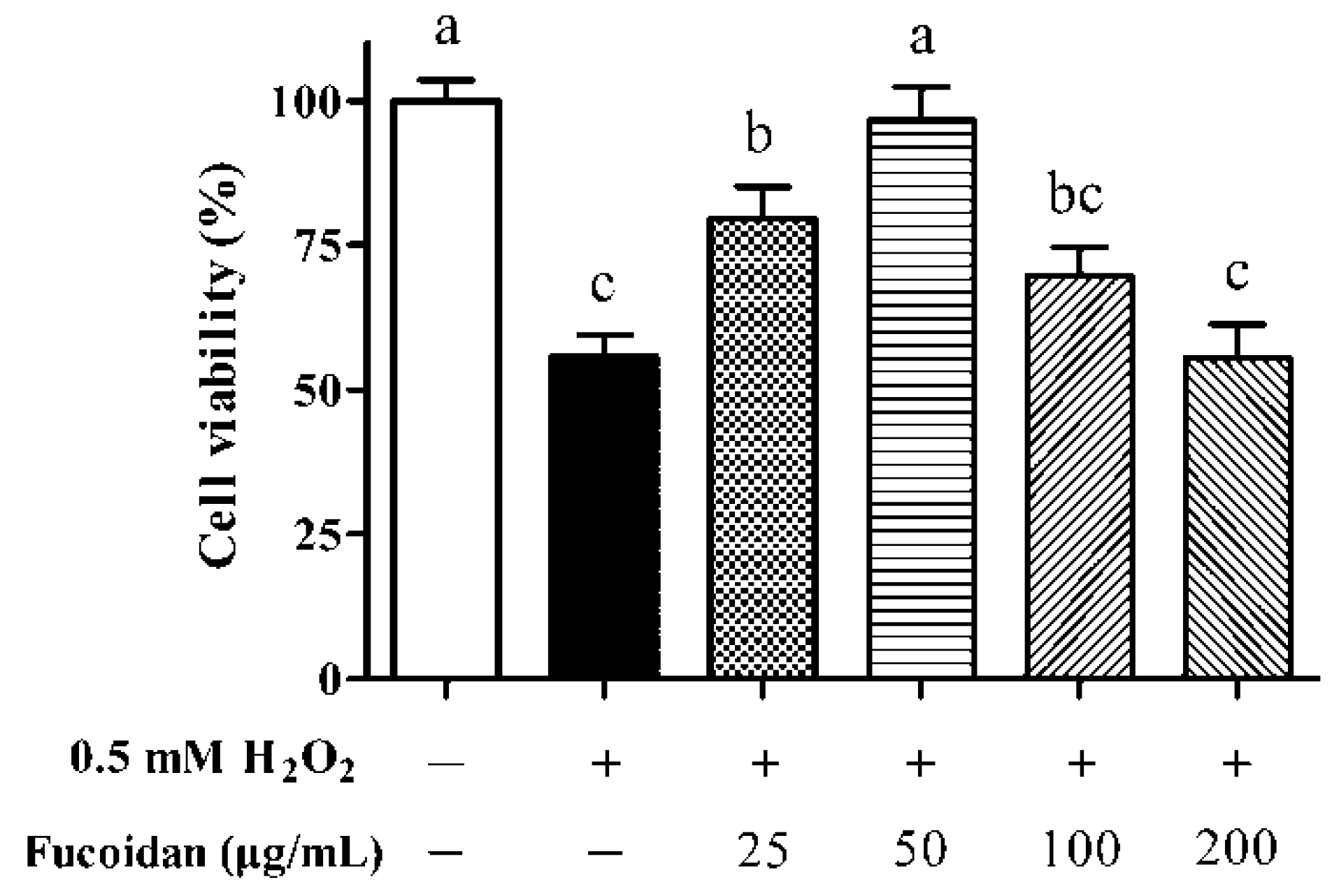
3.2. IPEC-1 Cell Viability in the Presence of Fucoidan
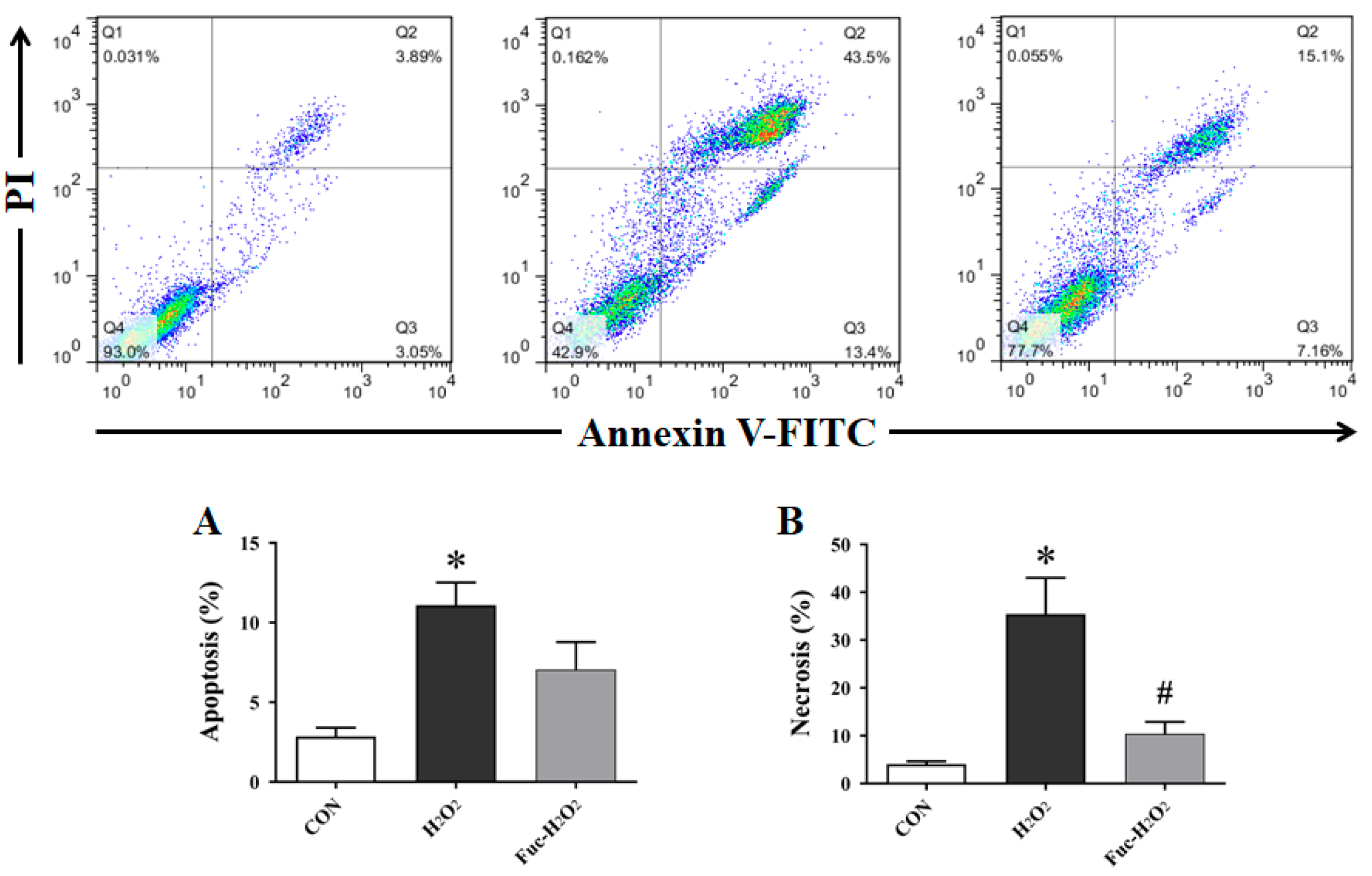
3.3. Cell Apoptosis and Necrosis
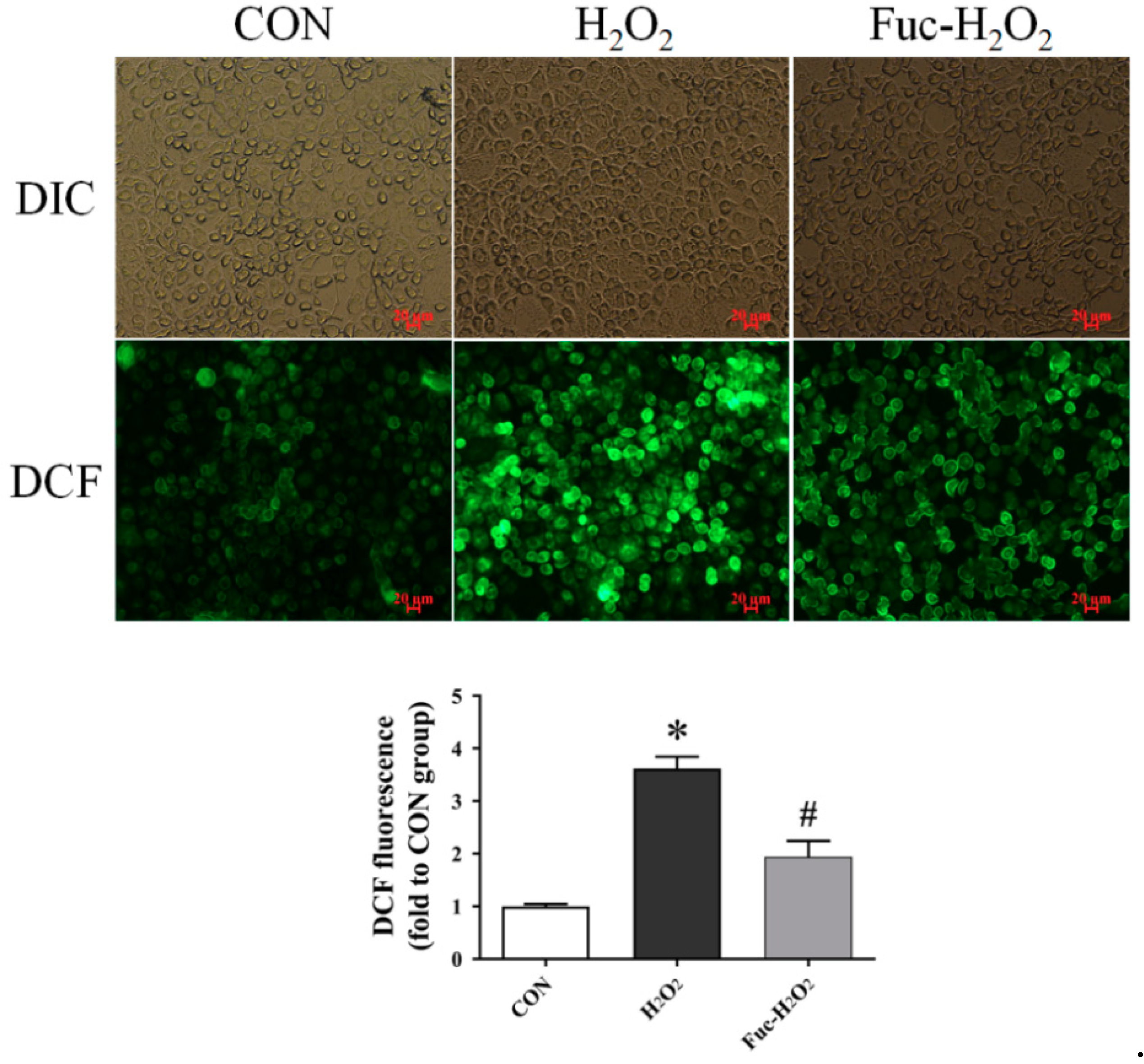
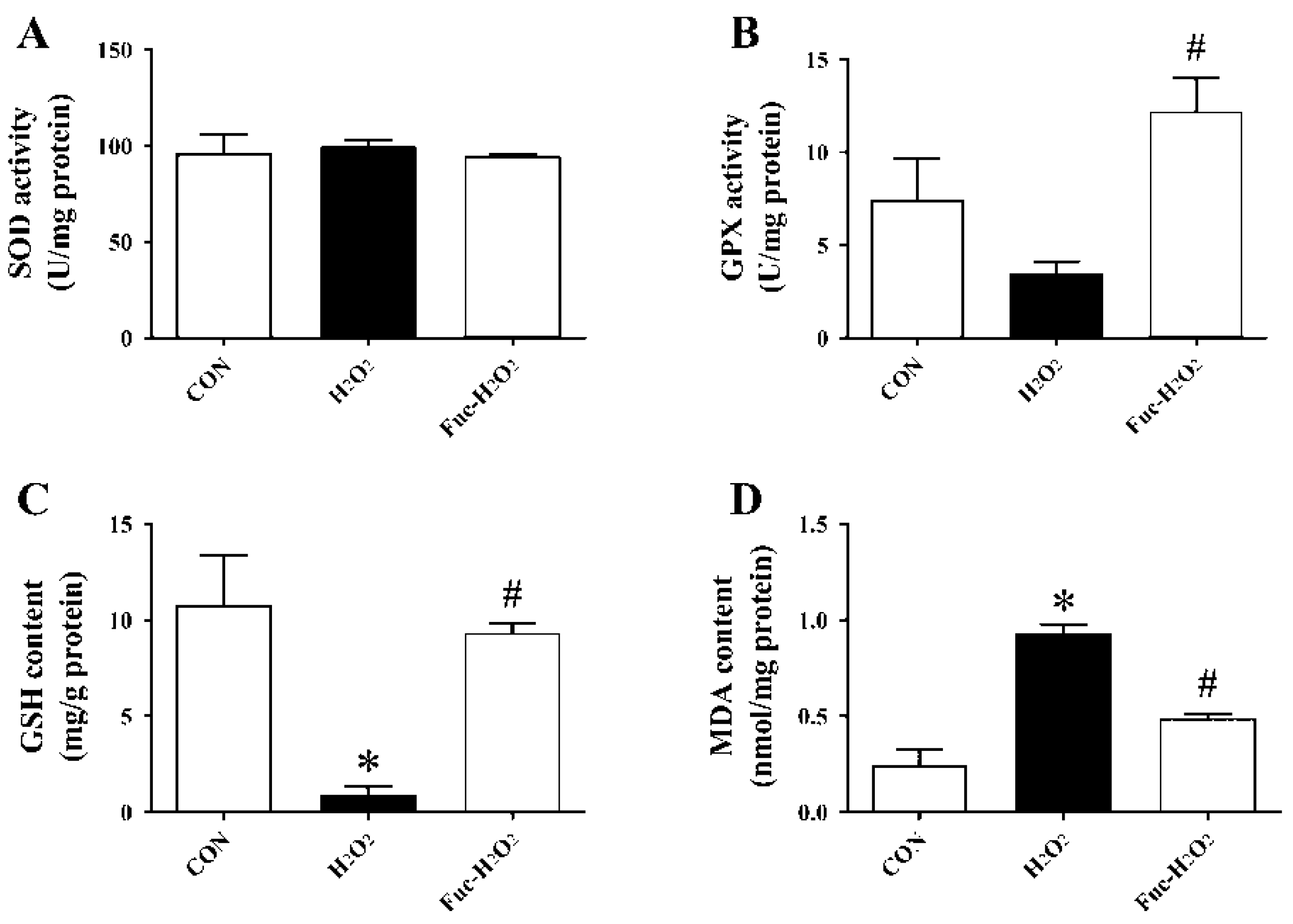
3.4. ROS Level and Antioxidant Capacity
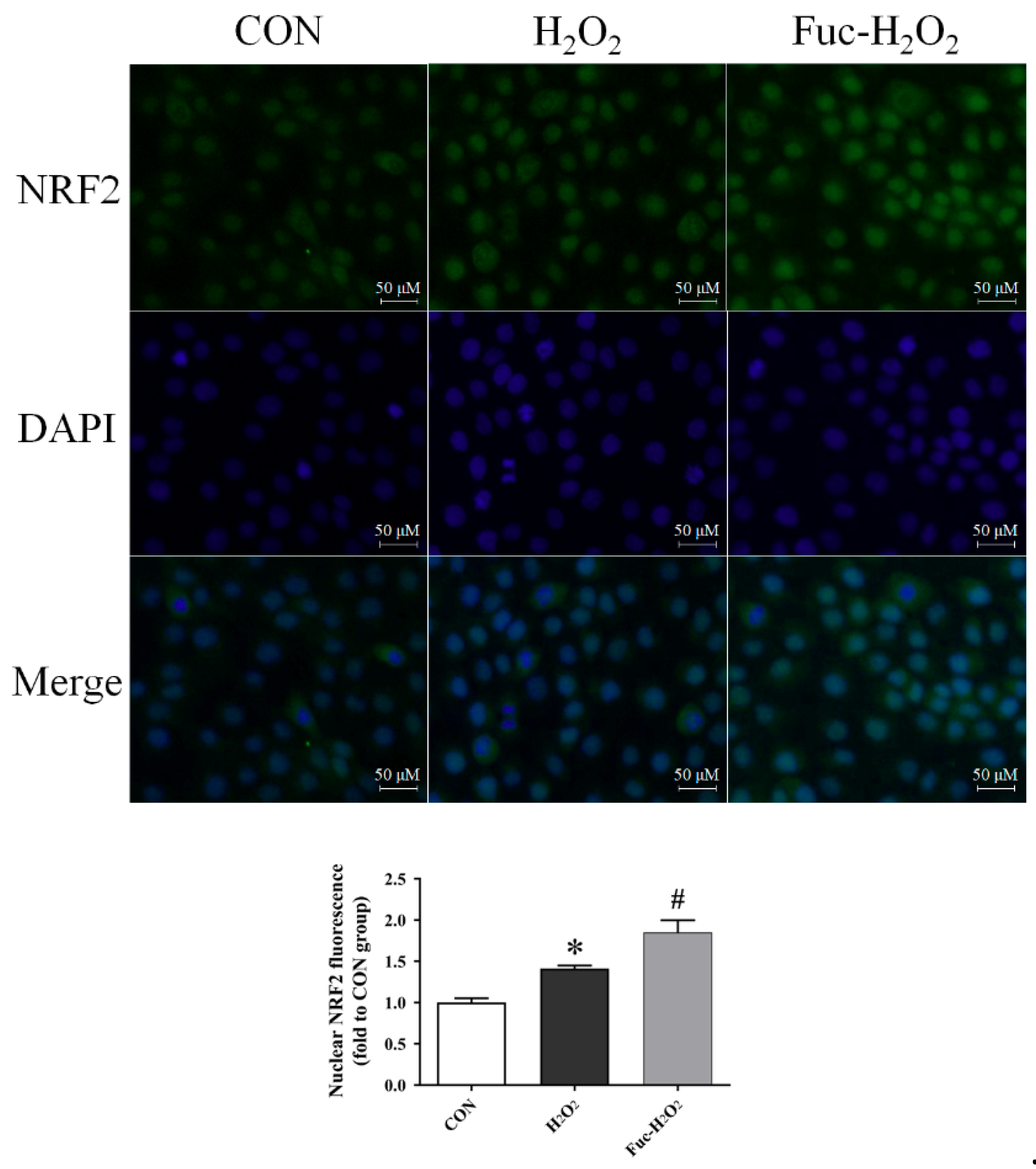
3.5. Gene Expression and the Nuclear Translocation of NRF2
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jones, R.M.; Neish, A.S. Redox signaling mediated by the gut microbiota. Free Radic. Biol. Med. 2017, 105, 41–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Aw, T.Y. Molecular and cellular responses to oxidative stress and changes in oxidation-reduction imbalance in the intestine. Am. J. Clin. Nutr. 1999, 70, 557–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Zhao, K.; Whiteman, M. The gastrointestinal tract: A major site of antioxidant action? Free Radic. Res. 2000, 33, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Pérez, S.; Taléns-Visconti, R.; Rius-Pérez, S.; Finamor, I.; Sastre, J. Redox signaling in the gastrointestinal tract. Free Radic. Biol. Med. 2017, 104, 75–103. [Google Scholar] [CrossRef] [PubMed]
- Blander, J.M. Death in the intestinal epithelium-basic biology and implications for inflammatory bowel disease. FEBS J. 2016, 283, 2720–2730. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative stress: An essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Vergauwen, H.; Tambuyzer, B.; Jennes, K.; Degroote, J.; Wang, W.; De Smet, S.; Michiels, J.; Van Ginneken, C. Trolox and ascorbic acid reduce direct and indirect oxidative stress in the IPEC-J2 cells, an in vitro model for the porcine gastrointestinal tract. PLoS ONE 2015, 10, 0120485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Zhu, L.H.; Chen, X.L.; Sheng, Y.S.; Guo, Q.; Bao, J.; Xu, J.X. X/XO or H2O2 induced IPEC-J2 cell as a new in vitro model for studying apoptosis in post-weaning piglets. Cytotechnology 2016, 68, 713–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Wang, J.; Peng, J.; Wei, H.K. Oregano essential oil induces SOD1 and GSH expression through Nrf2 activation and alleviates hydrogen peroxide-induced oxidative damage in IPEC-J2 cells. Oxid. Med. Cell. Longev. 2016, 2016, 5987183. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.F.; Wei, H.K.; Yu, H.C.; Xing, Q.; Zou, Y.; Zhou, Y.F.; Peng, J. Fish skin gelatin hydrolysate production by ginger powder induces glutathione synthesis to prevent hydrogen peroxide induced intestinal oxidative stress via the Pept1-p62-Nrf2 cascade. J. Agric. Food Chem. 2018, 66, 11601–11611. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Shen, M.Y.; Song, Q.Q.; Xie, J.H. Biological activities and pharmaceutical applications of polysaccharide from natural resources: A review. Carbohydr. Polym. 2018, 183, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.S.; Kim, S.K. Fucoidans as a natural bioactive ingredient for functional foods. J. Funct. Foods 2013, 5, 16–27. [Google Scholar] [CrossRef]
- Li, B.; Lu, F.; Wei, X.J.; Zhao, R.X. Fucoidan: Structure and bioactivity. Molecules 2008, 13, 1671–1695. [Google Scholar] [CrossRef] [Green Version]
- Phull, A.R.; Kim, S.J. Fucoidan as bio-functional molecule: Insights into the anti-inflammatory potential and associated molecular mechanisms. J. Funct. Foods 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Gao, Y.L.; Dong, C.H.; Yin, J.G.; Shen, J.Y.; Tian, J.W.; Li, C.M. Neuroprotective effect of fucoidan on H2O2-induced apoptosis in PC12 cells via activation of PI3K/Akt pathway. Cell. Mol. Neurobiol. 2012, 32, 523–529. [Google Scholar] [CrossRef]
- Roy Chowdhury, S.; Sengupta, S.; Biswas, S.; Sinha, T.K.; Sen, R.; Basak, R.K.; Adhikari, B.; Bhattacharyya, A. Bacterial fucose-rich polysaccharide stabilizes MAPK-mediated Nrf2/Keap1 signaling by directly scavenging reactive oxygen species during hydrogen peroxide-induced apoptosis of human lung fibroblast cells. PLoS ONE 2014, 9, 113663. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.S.; Lee, J.H.; Jung, J.S.; Noh, H.; Baek, M.J.; Ryu, J.M.; Yoon, Y.M.; Han, H.J.; Lee, S.H. Fucoidan protects mesenchymal stem cells against oxidative stress and enhances vascular regeneration in a murine hindlimb ischemia model. Int. J. Cardiol. 2015, 198, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Q.; Wei, J.G.; Tu, M.J.; Gu, J.G.; Zhang, W. Fucoidan alleviates acetaminophen-induced hepatotoxicity via oxidative stress inhibition and Nrf2 translocation. Int. J. Mol. Sci. 2018, 19, 4050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.Y.; Kuo, C.H.; Lee, C.H. Antibacterial and antioxidant capacities and attenuation of lipid accumulation in 3T3-L1 adipocytes by low-molecular-weight fucoidans prepared from compressional-puffing-pretreated Sargassum Crassifolium. Mar. Drugs 2018, 16, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, C.; Lahaye, M.; Bonnet, C.; Mabeau, S.; Barry, J.L. In vitro fermentation by human faecal bacteria of total and purified dietary fibres from brown seaweeds. Br. J. Nutr. 1996, 75, 263–280. [Google Scholar] [CrossRef] [PubMed]
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find. Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a fucoidan-specific antibody and measurement of fucoidan in serum and urine by sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350–357. [Google Scholar] [CrossRef]
- Paszti-Gere, E.; Csibrik-Nemeth, E.; Szeker, K.; Csizinszky, R.; Jakab, C.; Galfi, P. Acute oxidative stress affects IL-8 and TNF-α expression in IPEC-J2 porcine epithelial cells. Inflammation 2012, 35, 994–1004. [Google Scholar] [CrossRef]
- Zhang, H.; Su, W.P.; Ying, Z.X.; Chen, Y.P.; Zhou, L.; Li, Y.; Zhang, J.F.; Zhang, L.L.; Wang, T. N-acetylcysteine attenuates intrauterine growth retardation-induced hepatic damage in suckling piglets by improving glutathione synthesis and cellular homeostasis. Eur. J. Nutr. 2018, 57, 327–338. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Hafeman, D.G.; Sunde, R.A.; Hoekstra, W.G. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutr. 1974, 104, 580–587. [Google Scholar] [CrossRef]
- Owens, C.W.I.; Belcher, R.V. A colorimetric micro-method for the determination of glutathione. Biochem. J. 1965, 94, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 1966, 16, 359–364. [Google Scholar] [CrossRef]
- Kruger, N.J. The Bradford method for protein quantitation. Methods Mol. Biol. 1994, 32, 9–15. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; He, C.; Tian, K.; Li, P.; Su, H.; Wan, J.B. Ginsenoside Rb1 attenuates angiotensin II-induced abdominal aortic aneurysm through inactivation of the JNK and p38 signaling pathways. Vasc. Pharmacol. 2015, 73, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Xie, J.H.; Nie, S.P.; Xie, M.Y. Review on cell models to evaluate the potential antioxidant activity of polysaccharides. Food Funct. 2017, 8, 915–926. [Google Scholar] [CrossRef]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell. Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef]
- Burch, P.M.; Heintz, N.H. Redox regulation of cell-cycle re-entry: Cyclin D1 as a primary target for the mitogenic effects of reactive oxygen and nitrogen species. Antioxid. Redox Signal 2005, 7, 741–751. [Google Scholar] [CrossRef]
- Wang, J.; Liu, L.; Zhang, Q.B.; Zhang, Z.S.; Qi, H.M.; Li, P.C. Synthesized oversulphated, acetylated and benzoylated derivatives of fucoidan extracted from Laminaria japonica and their potential antioxidant activity in vitro. Food Chem. 2009, 114, 1285–1290. [Google Scholar] [CrossRef]
- Lim, S.J.; Aida, W.M.W.; Maskat, M.Y.; Mamot, S.; Ropien, J.; Mohd, D.M. Isolation and antioxidant capacity of fucoidan from selected Malaysian seaweeds. Food Hydrocoll. 2014, 42, 280–288. [Google Scholar] [CrossRef]
- Phull, A.R.; Majid, M.; Haq, I.U.; Khan, M.R.; Kim, S.J. In vitro and in vivo evaluation of anti-arthritic, antioxidant efficacy of fucoidan from Undaria pinnatifida (Harvey) Suringar. Int. J. Biol. Macromol. 2017, 97, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Jeong, B.E.; Ko, E.J.; Joo, H.G. Cytoprotective effects of fucoidan, an algae-derived polysaccharide on 5-fluorouracil-treated dendritic cells. Food Chem. Toxicol. 2012, 50, 1480–1484. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Chen, X.L.; Wang, X.C.; Xu, C.C.; Guo, Q.; Zhu, L.H.; Zhu, S.W.; Xu, J.X. Pre-protective effect of lipoic acid on injury induced by H2O2 in IPEC-J2 cells. Mol. Cell. Biochem. 2013, 378, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Kahlert, S.; Junnikkala, S.; Renner, L.; Hynönen, U.; Hartig, R.; Nossol, C.; Barta-Böszörményi, A.; Dänicke, S.; Souffrant, W.B.; Palva, A.; et al. Physiological concentration of exogenous lactate reduces antimycin a triggered oxidative stress in intestinal epithelial cell line IPEC-1 and IPEC-J2 in vitro. PLoS ONE 2016, 11, 0153135. [Google Scholar] [CrossRef]
- Thomas, L.W.; Lam, C.; Edwards, S.W. Mcl-1; the molecular regulation of protein function. FEBS Lett. 2010, 584, 2981–2989. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2: INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [Green Version]
- Vriend, J.; Reiter, R.J. The Keap1-Nrf2-antioxidant response element pathway: A review of its regulation by melatonin and the proteasome. Mol. Cell. Endocrinol. 2015, 401, 213–220. [Google Scholar] [CrossRef]
- Suzuki, T.; Motohashi, H.; Yamamoto, M. Toward clinical application of the Keap1-Nrf2 pathway. Trends Pharmacol. Sci. 2013, 34, 340–346. [Google Scholar] [CrossRef]
- Zhuang, S.; Yu, R.Y.; Zhong, J.; Liu, P.; Liu, Z.J. Rhein from Rheum rhabarbarum inhibits hydrogen-peroxide-induced oxidative stress in intestinal epithelial cells partly through PI3K/Akt-mediated Nrf2/HO-1 pathways. J. Agric. Food Chem. 2019, 67, 2519–2529. [Google Scholar] [CrossRef]
- Wen, Z.S.; Xue, R.; Du, M.; Tang, Z.; Xiang, X.W.; Zheng, B.; Qu, Y.L. Hemp seed polysaccharides protect intestinal epithelial cells from hydrogen peroxide-induced oxidative stress. Int. J. Biol. Macromol. 2019, 135, 203–211. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Genbank 1 | Sequence (5′→3′) 2 | Length |
|---|---|---|---|
| NRF2 | NM_001185152.1 | GGACAGCAGAAGTGATCCCC | 97 |
| CAAAACCGTATCACTGGCCG | |||
| HO1 | NM_001004027.1 | TGATGGCGTCCTTGTACCAC | 71 |
| GACCGGGTTCTCCTTGTTGT | |||
| NQO1 | NM_001159613.1 | CATGGCGGTCAGAAAAGCAC | 135 |
| ATGGCATACAGGTCCGACAC | |||
| SOD1 | NM_001190422.1 | AAGGCCGTGTGTGTGCTGAA | 118 |
| GATCACCTTCAGCCAGTCCTTT | |||
| SOD2 | NM_214127.2 | GGCCTACGTGAACAACCTGA | 126 |
| TGATTGATGTGGCCTCCACC | |||
| GPX1 | NM_214201.1 | CCTCAAGTACGTCCGACCAG | 85 |
| GTGAGCATTTGCGCCATTCA | |||
| GSTA1 | NM_214389.2 | ACACCCAGGACCAATCTTCTG | 199 |
| AGTCTCAGGTACATTCCGGG | |||
| CAT | NM_214301.2 | TCCAGCCAGTGACCAGATGA | 182 |
| CCCGGTCAAAGTGAGCCATT | |||
| PCNA | NM_001291925.1 | TAGCCGCGTCGTTGTGATTC | 105 |
| GGCCTCGTTGATGAGGTCTT | |||
| CASP3 | NM_214131.1 | GGATTGAGACGGACAGTGGG | 124 |
| CCGTCCTTTGAATTTCGCCA | |||
| CASP9 | XM_003127618.4 | CTGCCAAGCAAATGGTCCAG | 151 |
| ACAGGACATCCATCTGTGCC | |||
| MCL1 | NM_001348806.1 | CGGAGTAACAAACTGGGGCA | 177 |
| AACCCATCCCAGCCTCTTTG | |||
| BCL2 | XM_021099593.1 | GAGTTCGGTGGGGTCATGTG | 152 |
| TACAGCTCCACAAAGGCATCC | |||
| BAX | XM_003127290.5 | GGCCCTTTTGCTTCAGGGTTT | 119 |
| GACACTCGCTCAACTTCTTGG | |||
| GAPDH | NM_001206359.1 | CCAAGGAGTAAGAGCCCCTG | 125 |
| AAGTCAGGAGATGCTCGGTG | |||
| ACTB | XM_003124280.5 | TGGAACGGTGAAGGTGACAG | 176 |
| CTTTTGGGAAGGCAGGGACT |
| Items 1 | CON | H2O2 | Fuc-H2O2 | Contrast | |
|---|---|---|---|---|---|
| CON vs. H2O2 | H2O2 vs. Fuc-H2O2 | ||||
| NRF2 | 1.00 ± 0.14 | 8.10 ± 1.99 | 8.97 ± 2.14 | 0.056 | 0.931 |
| HO1 | 1.00 ± 0.04 | 1.01 ± 0.09 | 1.08 ± 0.12 | 0.997 | 0.821 |
| NQO1 | 1.00 ± 0.07 | 0.22 ± 0.02 * | 0.52 ± 0.07 # | <0.001 | 0.024 |
| SOD1 | 1.00 ± 0.08 | 0.58 ± 0.04 * | 0.91 ± 0.08 # | 0.014 | 0.037 |
| SOD2 | 1.00 ± 0.08 | 0.63 ± 0.07 | 1.06 ± 0.22 | 0.236 | 0.163 |
| GPX1 | 1.00 ± 0.02 | 1.11 ± 0.10 | 1.86 ± 0.18 # | 0.797 | 0.011 |
| GSTA1 | 1.00 ± 0.25 | 1.82 ± 0.42 | 1.75 ± 0.26 | 0.247 | 0.986 |
| CAT | 1.00 ± 0.03 | 0.93 ± 0.06 | 0.97 ± 0.08 | 0.729 | 0.892 |
| PCNA | 1.00 ± 0.06 | 0.97 ± 0.13 | 1.10 ± 0.18 | 0.988 | 0.784 |
| CASP3 | 1.00 ± 0.28 | 1.04 ± 0.41 | 0.97 ± 0.37 | 0.997 | 0.991 |
| CASP9 | 1.00 ± 0.08 | 1.04 ± 0.05 | 0.89 ± 0.07 | 0.920 | 0.341 |
| MCL1 | 1.00 ± 0.09 | 0.60 ± 0.08 * | 0.69 ± 0.04 | 0.020 | 0.668 |
| BCL2 | 1.00 ± 0.12 | 0.68 ± 0.12 | 0.77 ± 0.15 | 0.280 | 0.867 |
| BAX | 1.00 ± 0.16 | 0.99 ± 0.17 | 1.09 ± 0.23 | 0.999 | 0.919 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, W.; Wang, L.; Chen, Y.; Zhang, H.; Wang, T.; Yang, X.; Xing, F.; Yan, J.; Fang, X. Protective Effects of Fucoidan against Hydrogen Peroxide-Induced Oxidative Damage in Porcine Intestinal Epithelial Cells. Animals 2019, 9, 1108. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9121108
Li Y, Zhao W, Wang L, Chen Y, Zhang H, Wang T, Yang X, Xing F, Yan J, Fang X. Protective Effects of Fucoidan against Hydrogen Peroxide-Induced Oxidative Damage in Porcine Intestinal Epithelial Cells. Animals. 2019; 9(12):1108. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9121108
Chicago/Turabian StyleLi, Yue, Weimin Zhao, Li Wang, Yueping Chen, Hao Zhang, Tian Wang, Xiaoyang Yang, Fei Xing, Junshu Yan, and Xiaomin Fang. 2019. "Protective Effects of Fucoidan against Hydrogen Peroxide-Induced Oxidative Damage in Porcine Intestinal Epithelial Cells" Animals 9, no. 12: 1108. https://0-doi-org.brum.beds.ac.uk/10.3390/ani9121108