
The Link between Occurrence of Class I Integron and Acquired Aminoglycoside Resistance in Clinical MRSA Isolates

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Identification of S. aureus Isolates
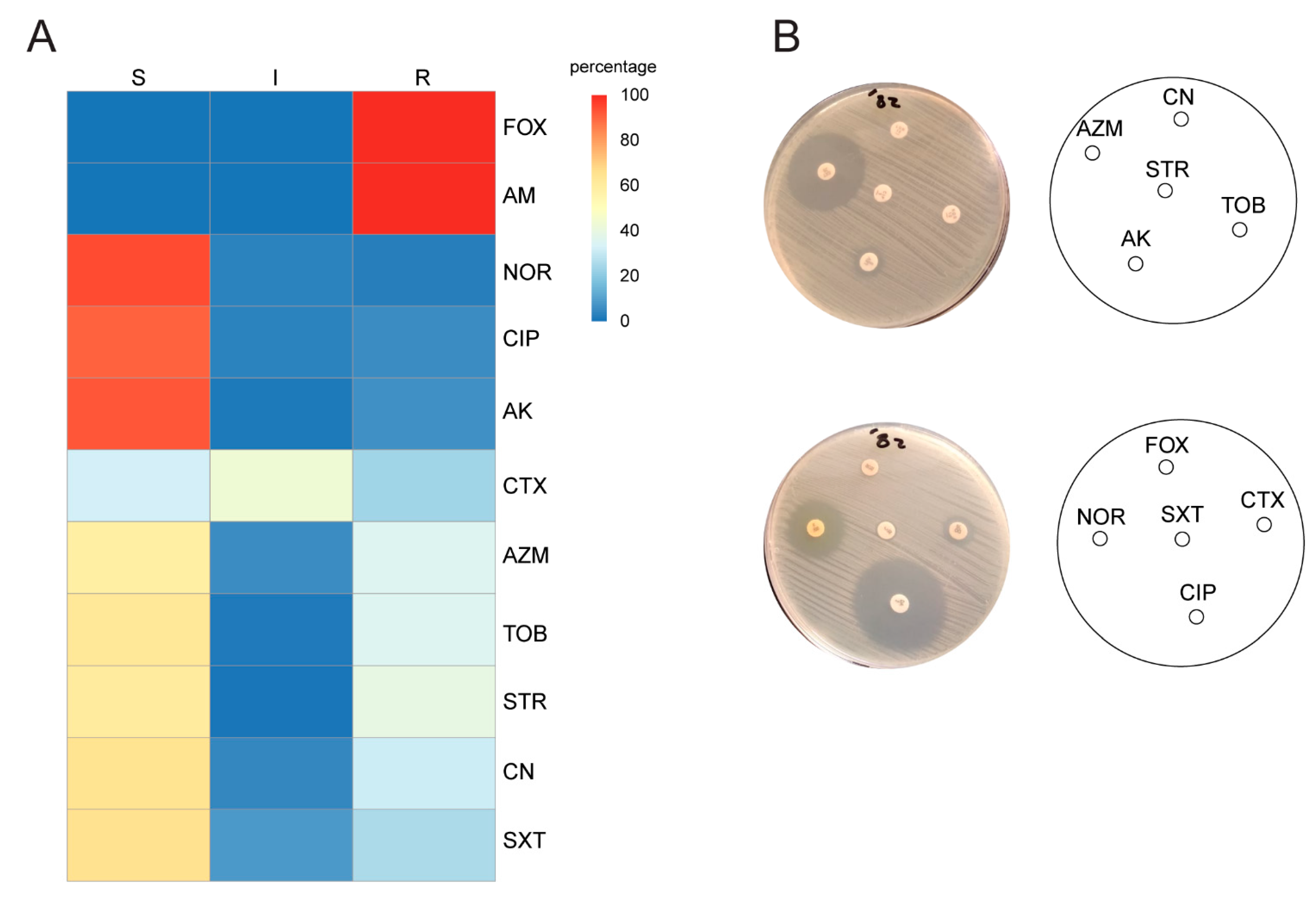
2.2. Antimicrobial Susceptibility Testing
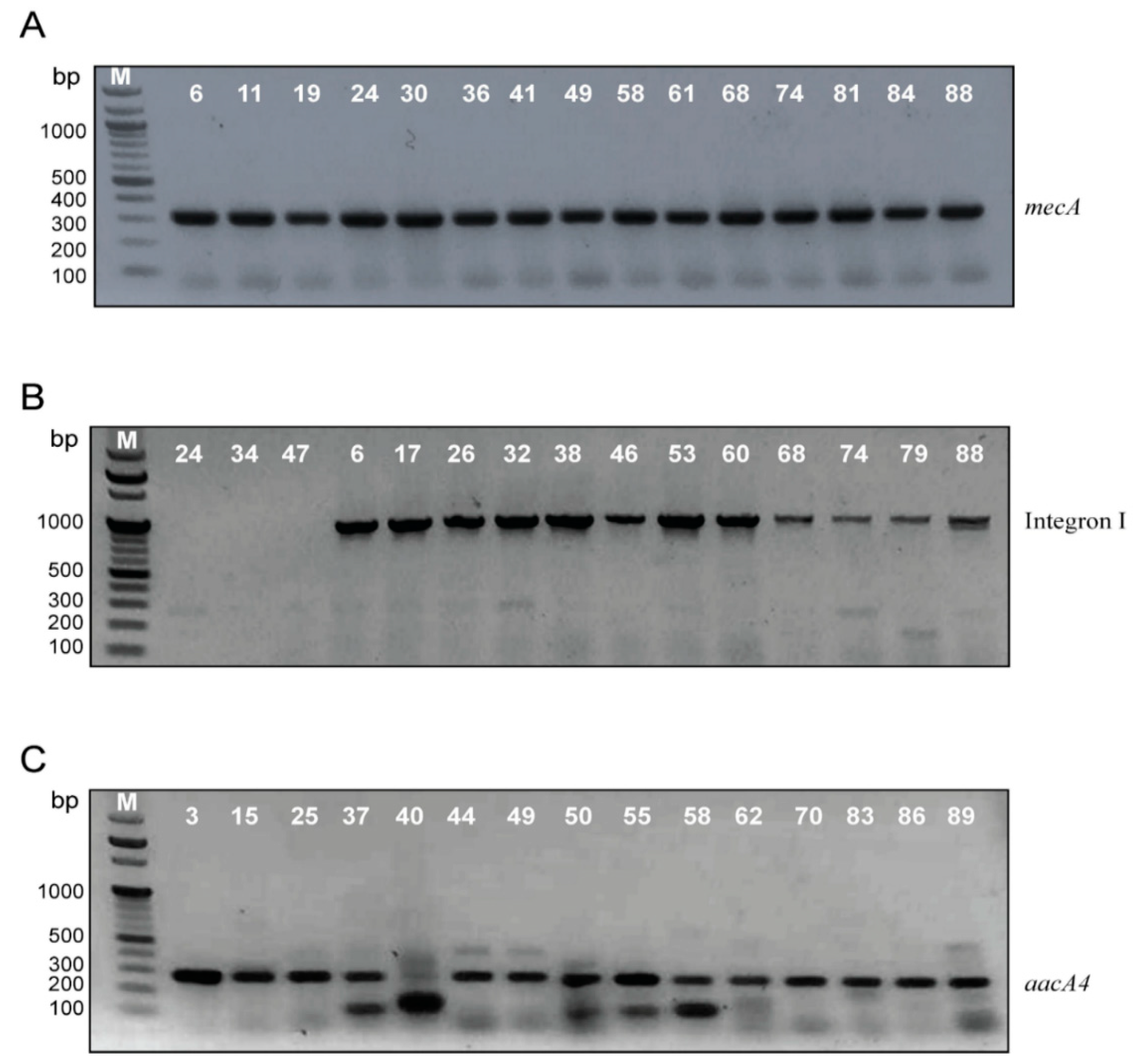
2.3. Confirmation of MRSA Identification, Screening of Integron I, II and aacA4 Genes by Polymerase Chain Reaction (PCR)
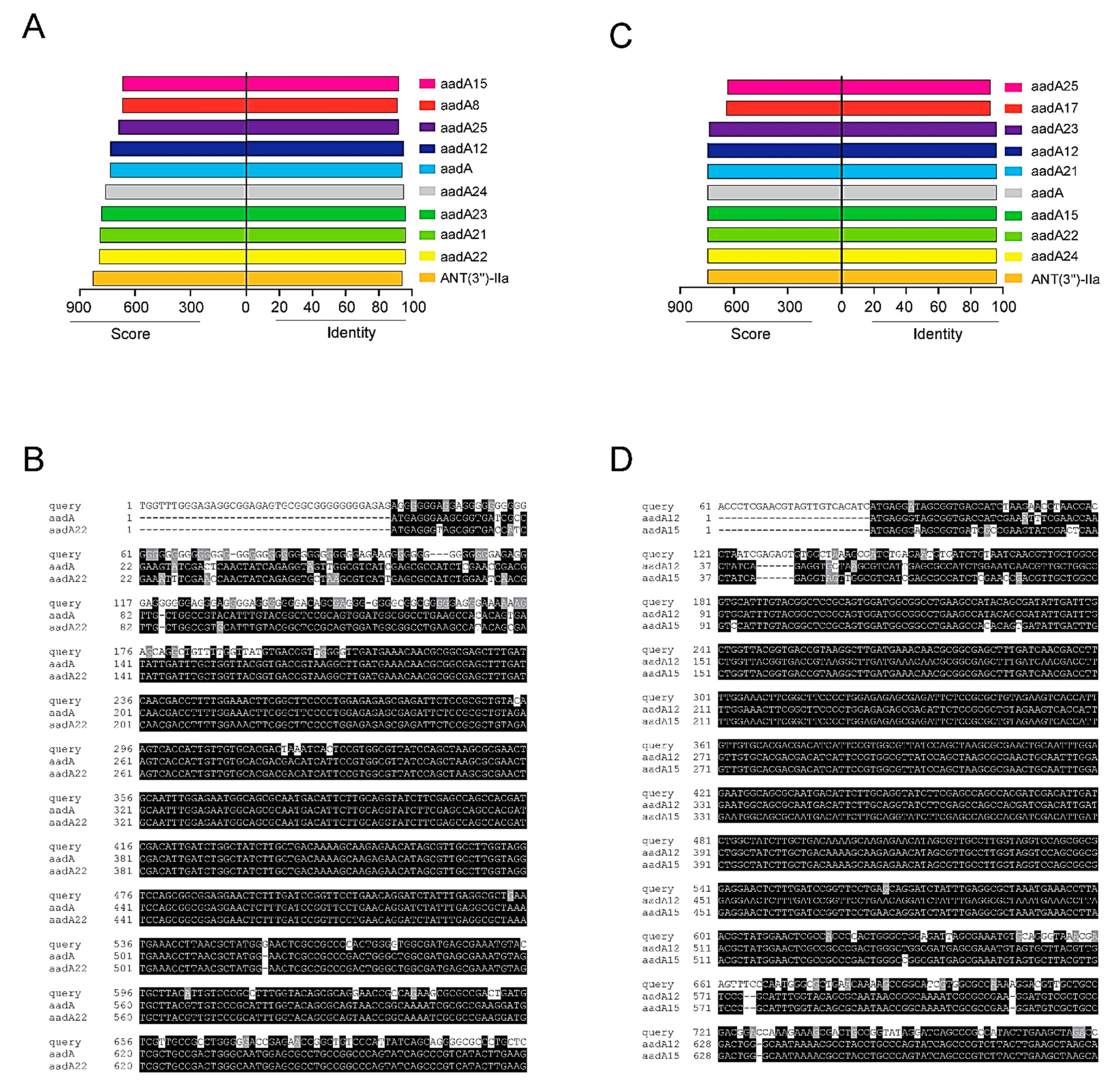
2.4. Sequencing of Integron I-Positive Isolates
2.5. Classification of Isolates According to Resistance Profile, Integron I and aacA4 Presence
3. Discussion
4. Materials and Methods
4.1. Bacteria Isolation and Identifications
4.2. Antimicrobial Susceptibility
4.3. Amplification of Integron I, Integron II, mecA and aacA4 Genes Using PCR
4.4. Sequencing of Amplified Integrons
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pollitt, E.J.; Szkuta, P.T.; Burns, N.; Foster, S.J. Staphylococcus aureus infection dynamics. PLoS Pathog. 2018, 14, e1007112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbial. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.Y.; Mojica, N.; Jiang, W.; Cosgrove, S.E.; Septimus, E.; Morgan, D.J.; Laxminarayan, R. Trends in methicillin-resistant Staphylococcus aureus hospitalizations in the United States, 2010–2014. Clin. Infect. Dis. 2017, 65, 1921–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichereau, S.; Rose, W.E. Invasive community-associated MRSA infections: Epidemiology and antimicrobial management. Expert Opin. Pharmacother. 2010, 11, 3009–3025. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Community-associated MRSA: What makes them special? Int. J. Med. Microbiol. 2013, 303, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassoun, A.; Linden, P.K.; Friedman, B. Incidence, prevalence, and management of MRSA bacteremia across patient populations—A review of recent developments in MRSA management and treatment. Crit. Care 2017, 21, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lakhundi, S.; Zhang, K. Methicillin-resistant Staphylococcus aureus: Molecular characterization, evolution, and epidemiology. Clin. Microbial. Rev. 2018, 31, e00020-18. [Google Scholar] [CrossRef] [Green Version]
- Daef, E.A.; Elsherbiny, N.M.; Ibrahim, M.A.; Ahmed, E.H. Decolonization of methicillin resistant Staphylococcus aureus nasal carriage among health care workers. Life Sci. J. 2012, 9, 223–227. [Google Scholar]
- Abdel-Maksoud, M.; El-Shokry, M.; Ismail, G.; Hafez, S.; El-Kholy, A.; Attia, E.; Talaat, M. Methicillin-resistant Staphylococcus aureus recovered from healthcare-and community-associated infections in Egypt. Int. J Bact. 2016, 2016, 5751785. [Google Scholar] [CrossRef] [Green Version]
- Stokes, H.W.; Gillings, M.R. Gene flow, mobile genetic elements and the recruitment of antibiotic resistance genes into Gram-negative pathogens. FEMS Microbial. Rev. 2011, 35, 790–819. [Google Scholar] [CrossRef]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Kadry, A.A.; Serry, F.M.; El-Ganiny, A.M.; El-Baz, A.M. Integron occurrence is linked to reduced biocide susceptibility in multidrug resistant Pseudomonas aeruginosa. Br. J Biomed. Sci. 2017, 74, 78–84. [Google Scholar] [CrossRef]
- Mostafa, M.; Siadat, S.D.; Shahcheraghi, F.; Vaziri, F.; Japoni-Nejad, A.; Yousefi, J.V.; Rajaei, B.; Mood, E.H.; Moshiri, A.; Siamdoust, S.A.S. Variability in gene cassette patterns of class 1 and 2 integrons associated with multi drug resistance patterns in Staphylococcus aureus clinical isolates in Tehran-Iran. BMC Microbial. 2015, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, C.; Zhao, Y.; Shen, Y. Analysis of the effect of integrons on drug-resistant Staphylococcus aureus by multiplex PCR detection. Mol. Med. Rep. 2013, 7, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, M.; Bahrami, N.; Khajavian, M.; Faghri, J. The Occurrence of Type I, II, and III Integrons in Multi-drug Resistance and Methicillin-Resistant Staphylococcus aureus Isolates in Iran. Curr. Microbial. 2020, 77, 1653–1659. [Google Scholar] [CrossRef]
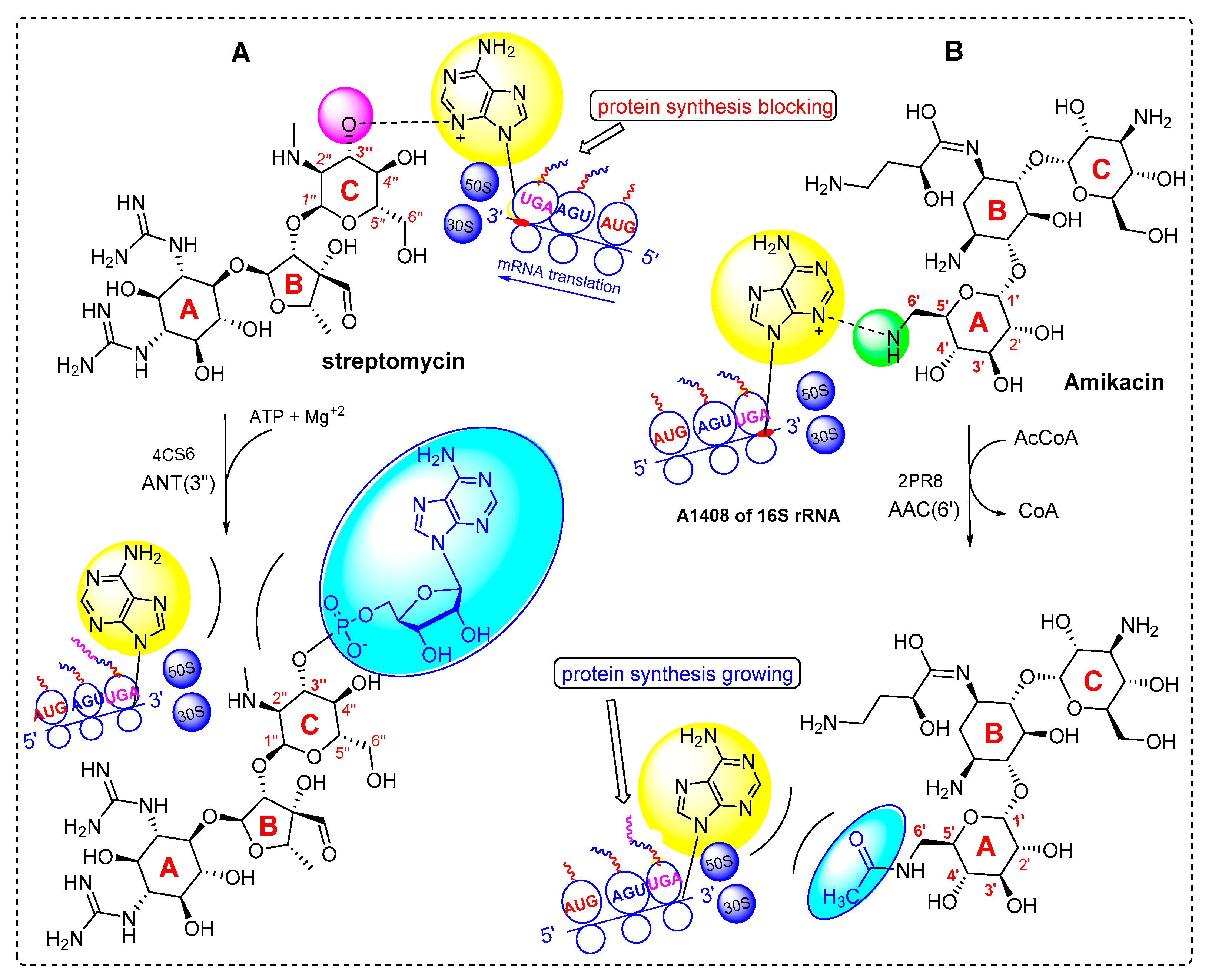
- Zárate, S.G.; De la Cruz Claure, M.L.; Benito-Arenas, R.; Revuelta, J.; Santana, A.G.; Bastida, A. Overcoming aminoglycoside enzymatic resistance: Design of novel antibiotics and inhibitors. Molecules 2018, 23, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitz, F.-J.; Fluit, A.C.; Gondolf, M.; Beyrau, R.; Lindenlauf, E.; Verhoef, J.; Heinz, H.-P.; Jones, M.E. The prevalence of aminoglycoside resistance and corresponding resistance genes in clinical isolates of staphylococci from 19 European hospitals. J. Antimicrob. Chemother. 1999, 43, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Serio, A.; Magalhães, M.; Blanchard, J.; Connolly, L. Aminoglycosides: Mechanisms of Action and Resistance. In Antimicrobial Drug Resistance; National Institute of Allergy and Infectious Diseases: Bethesda, MD, USA, 2017; pp. 213–229. [Google Scholar]
- Ramirez, M.S.; Tolmasky, M.E. Amikacin: Uses, resistance, and prospects for inhibition. Molecules 2017, 22, 2267. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, M.S.; Nikolaidis, N.; Tolmasky, M. Rise and dissemination of aminoglycoside resistance: The aac (6′)-Ib paradigm. Front. Microbiol. 2013, 4, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H.; Chang, W. Computer Software. 2012. Available online: http://ggplot2.org (accessed on 15 January 2021).
- Hofmann, K.; Baron, M.B. Version 3.21 of BOXSHADE. Available online: http://sourceforge.net/projects/boxshade/ (accessed on 15 January 2021).
- Xu, Z.; Li, L.; Shirtliff, M.; Peters, B.; Li, B.; Peng, Y.; Alam, M.J.; Yamasaki, S.; Shi, L. Resistance Class 1 integron in clinical methicillin-resistant Staphylococcus aureus strains in southern China, 2001–2006. Clin. Microbiol. Infect. 2011, 17, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Velasco, D.; del Mar Tomas, M.; Cartelle, M.; Beceiro, A.; Perez, A.; Molina, F.; Moure, R.; Villanueva, R.; Bou, G. Evaluation of different methods for detecting methicillin (oxacillin) resistance in Staphylococcus aureus. J. Antimicrob. Chemother. 2005, 55, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Sousa Júnior, F.C.d.; Néri, G.d.S.; Silva, A.K.; Araújo, B.P.R.C.d.; Guerra, M.J.d.P.D.; Fernandes, M.J.d.B.C.; Milan, E.P.; Melo, M.C.N.d. Evaluation of different methods for detecting methicillin resistance in Staphylococcus aureus isolates in a university hospital located in the Northeast of Brazil. Braz. J. Microbiol. 2010, 41, 316–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadry, A.; Shaker, G.; El-Ganiny, A.; Youssef, C. Phenotypic and Genotypic detection of local MRSA isolates. ZJPS 2016, 25, 39–46. [Google Scholar] [CrossRef]
- Gurung, R.R.; Maharjan, P.; Chhetri, G.G. Antibiotic resistance pattern of Staphylococcus aureus with reference to MRSA isolates from pediatric patients. Future Sci. OA 2020, 6, FSO464. [Google Scholar] [CrossRef] [Green Version]
- Garoy, E.Y.; Gebreab, Y.B.; Achila, O.O.; Tekeste, D.G.; Kesete, R.; Ghirmay, R.; Kiflay, R.; Tesfu, T. Methicillin-resistant Staphylococcus aureus (MRSA): Prevalence and antimicrobial sensitivity pattern among patients—A multicenter study in Asmara, Eritrea. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, 8321834. [Google Scholar] [CrossRef] [Green Version]
- El Aila, N.A.; Al Laham, N.A.; Ayesh, B.M. Nasal carriage of methicillin resistant Staphylococcus aureus among health care workers at Al Shifa hospital in Gaza Strip. BMC Infect. Dis. 2017, 17, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kaur, D.C.; Chate, S.S. Study of antibiotic resistance pattern in methicillin resistant Staphylococcus aureus with special reference to newer antibiotic. J. Glob. Infect. Dis. 2015, 7, 78. [Google Scholar] [CrossRef]
- Li, L.; Zhao, X. Characterization of the resistance class 1 integrons in Staphylococcus aureus isolates from milk of lactating dairy cattle in Northwestern China. BMC Vet. Res. 2018, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.; Yi, M.; Shao, C.; Ma, J.; Zhang, Q.; Shao, S. Novel class 1 integrons in multi-drug resistant isolates from Eastern China. Indian J. Microbiol. 2014, 54, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grape, M.; Farra, A.; Kronvall, G.; Sundström, L. Integrons and gene cassettes in clinical isolates of co-trimoxazole-resistant Gram-negative bacteria. Clin. Microbiol. Infect. 2005, 11, 185–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firoozeh, F.; Mahluji, Z.; Khorshidi, A.; Zibaei, M. Molecular characterization of class 1, 2 and 3 integrons in clinical multi-drug resistant Klebsiella pneumoniae isolates. Antimicrob. Resist. Infect. Control 2019, 8, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrenberg, C.; Catry, B.; Haesebrouck, F.; de Kruif, A.; Schwarz, S. Novel spectinomycin/streptomycin resistance gene, aadA14, from Pasteurella multocida. Antimicrob. Agents Chemother. 2005, 49, 3046–3049. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.; Klima, C.L.; Ha, R.; Gruninger, R.J.; Zaheer, R.; McAllister, T.A. A novel aadA aminoglycoside resistance gene in bovine and porcine pathogens. mSphere 2018, 3, e00568-17. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.; Bertran, S.; Sauca, G.; Julià, A.; Vila, X.; Gómez, E.; Jiménez de Anta, M.; Vila, J. Isolation of an amikacin-resistant Escherichia coli strain after tobramycin treatment of previous recurrent episodes of respiratory tract infections caused by Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2005, 11, 71–73. [Google Scholar] [CrossRef] [Green Version]
- Green, K.D.; Garneau-Tsodikova, S. Domain dissection and characterization of the aminoglycoside resistance enzyme ANT (3″)-Ii/AAC (6′)-IId from Serratia marcescens. Biochimie 2013, 95, 1319–1325. [Google Scholar] [CrossRef] [Green Version]
- Hilal-Dandan, R.; Brunton, L. Aminoglycosides. Goodman and Gilman’s Manual of Pharmacology and Therapeutics, 2nd ed.; McGraw-Hill Education: New York, NY, USA, 2016. [Google Scholar]
- Boerlin, P.; Kuhnert, P.; Hüssy, D.; Schaellibaum, M. Methods for identification of Staphylococcus aureus isolates in cases of bovine mastitis. J. Clin. Microbiol. 2003, 41, 767–771. [Google Scholar] [CrossRef] [Green Version]
- Weinstein, M.P.; Lewis, J.S. The clinical and laboratory standards institute subcommittee on antimicrobial susceptibility testing: Background, organization, functions, and processes. J. Clin. Microbial. 2020, 58, e01864-19. [Google Scholar] [CrossRef] [PubMed]
- Dilhari, A.; Sampath, A.; Gunasekara, C.; Fernando, N.; Weerasekara, D.; Sissons, C.; McBain, A.; Weerasekera, M. Evaluation of the impact of six different DNA extraction methods for the representation of the microbial community associated with human chronic wound infections using a gel-based DNA profiling method. AMB Express 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, M.; Sreevatsan, S.; Davies, P.R. Prevalence and characterization of Staphylococcus aureus in growing pigs in the USA. PLoS ONE 2015, 10, e0143670. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Shi, L.; Yang, L.; Xiao, Z.; Li, X.; Yamasaki, S. Analysis of integrons in clinical isolates of Escherichia coli in China during the last six years. FEMS Microbiol. Lett. 2006, 254, 75–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sacha, P.; Jaworowska, J.; Ojdana, D.; Wieczorek, P.; Czaban, S.; Tryniszewska, E. Occurrence of the aacA4 gene among multidrug resistant strains of Pseudomonas aeruginosa isolated from bronchial secretions obtained from the intensive therapy unit at University hospital in Bialystok, Poland. Folia Histochem. Cytobiol. 2012, 50, 322–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
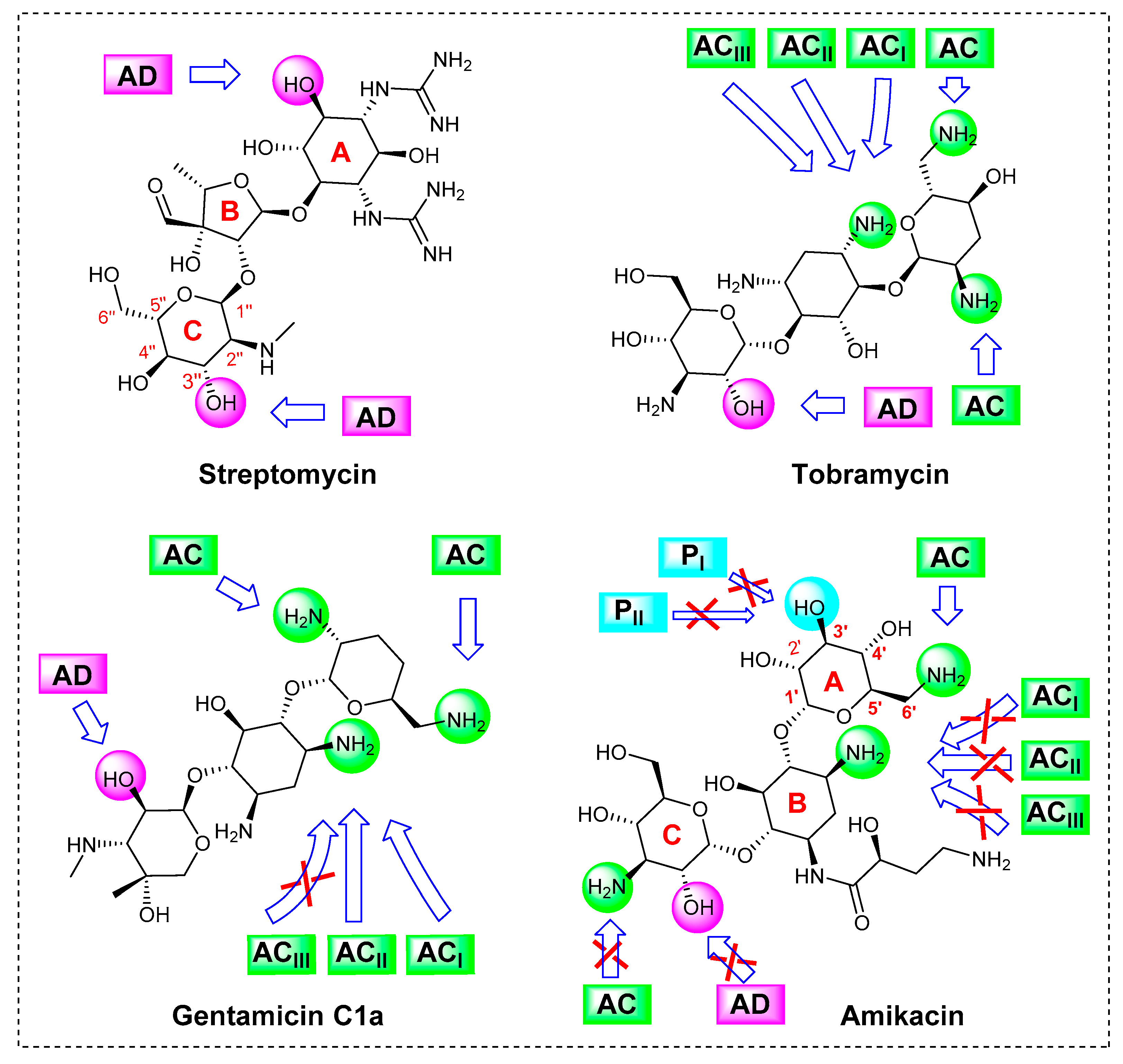{kind=link}
| Isolate No | Integron I (bp) | Sequence Profile of Integron I | aacA4 Gene | Antibiotic Resistance Profile |
|---|---|---|---|---|
| 6 | 1000 | query 2 (aadA12,15) | − | FOX, AM, CTX (I), STR, SXT |
| 8 | 1000 | query 1 (aadA22,23) | − | FOX, AM, STR, CTX(I), SXT(I) |
| 11 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX (I), STR |
| 13 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR AZM, |
| 15 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX(I), STR, TOB (I), CN (I), AK(I) AZM |
| 17 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, SXT(I), AZM |
| 19 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, SXT |
| 20 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX(I), STR, SXT |
| 21 | 1000 | query 1 (aadA22,23) | + | FOX, AM, CTX(I), STR, CN, TOB, AK, AZM |
| 24 | 250 | Empty cassette | − | FOX, AM |
| 25 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX(I), STR, CN, TOB, AK, AZM(I) |
| 26 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, SXT |
| 27 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX (I), STR, AZM |
| 28 | 1000 | query 2 (aadA12,15) | − | FOX, AM, STR |
| 29 | 250 | Empty cassette | − | FOX, AM |
| 30 | 1000 | query 1 (aadA22,23) | − | FOX, AM, STR |
| 31 | 250 | Empty cassette | − | FOX, AM |
| 32 | 1000 | query 2 (aadA12,15) | − | FOX, AM, CTX (I), STR, SXT |
| 34 | 250 | Empty cassette | − | FOX, AM |
| 35 | 1000 | query 1 (aadA22,23) | − | FOX, AM, STR, SXT |
| 36 | 1000 | query 1 (aadA22,23) | − | FOX, AM, STR, AZM |
| 37 | 1000 | query 2 (aadA12,15) | + | FOX, AM, STR, AK, CN, TOB, AZM, SXT |
| 38 | 1000 | query 1 (aadA22,23) | − | FOX, AM, STR, AZM |
| 41 | 250 | Empty cassette | − | FOX, AM |
| 42 | 1000 | query 1 (aadA22,23) | + | FOX, AM, CTX (I), STR, AK, CN, TOB, AZM, SXT |
| 46 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX (I), STR, AK, CN, TOB, AZM |
| 47 | 250 | Empty cassette | − | FOX, AM |
| 49 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX, STR, AK, CN, TOB, NOR, CIP, SXT, AZM |
| 53 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, AZM |
| 58 | 1000 | query 1 (aadA22,23) | + | FOX, AM, CTX(I), STR, CN, TOB, AK, AZM |
| 60 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX(I), CIP, NOR(I), STR, SXT(I) |
| 61 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX(I), STR, CN, TOB, AK(I) |
| 65 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, AZM |
| 68 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, SXT |
| 71 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX, STR, CN, TOB, AK(I), AZM |
| 74 | 1000 | query 1 (aadA22,23) | − | FOX, AM, CTX, STR, SXT |
| 78 | 1000 | query 2 (aadA12,15) | − | FOX, AM, CTX(I), STR, SXT |
| 79 | 1000 | query 1 (aadA22,23) | − | FOX, AM, STR, SXT |
| 81 | 1000 | query 1 (aadA22,23) | + | FOX, AM, STR, AK, CN, TOB, AZM, SXT |
| 84 | 1000 | query 1 (aadA22,23) | + | FOX, AM, CTX(I), STR, AK, CN, TOB, AZM, SXT |
| 88 | 1000 | query 2 (aadA12,15) | + | FOX, AM, CTX, STR, CN, TOB, AK, AZM |
| Isolate No. | Integron I | aacA4 Gene | Antibiotic Resistance Profile |
|---|---|---|---|
| 3 | − | + | FOX, AM, CTX (I), CN, TOB, STR, |
| 4 | − | + | FOX, AM, CTX, CN (I), TOB, STR, SXT |
| 5 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM |
| 23 | − | + | FOX, AM, CTX, CN, TOB, STR, AZM |
| 40 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM (I) |
| 43 | − | + | FOX, AM, CN, TOB, STR, AZM (I) |
| 44 | − | + | FOX, AM, CTX, CIP (I), CN, TOB, STR, AZM |
| 48 | − | + | FOX, AM, CTX (I), CN (I), TOB, STR, AZM |
| 50 | − | + | FOX, AM, CTX, CN, TOB, STR, AZM |
| 51 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM |
| 55 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM |
| 57 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM |
| 62 | − | + | FOX, AM, CTX, CN (I), TOB, STR, SXT |
| 70 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM |
| 85 | − | + | FOX, AM, CN, TOB, STR, AZM (I) |
| 86 | − | + | FOX, AM, CTX, CIP (I), CN, TOB, STR, AZM |
| 87 | − | + | FOX, AM, CTX(I), CIP, NOR, AK, CN, TOB, STR, AZM, SXT |
| 89 | − | + | FOX, AM, CTX (I), CN, TOB, STR, AZM |
| Primer | Sequence (5′-3′) | Amplicon Size | Target | Annealing Temperature | Reference |
|---|---|---|---|---|---|
| mecA-F | GTAGAAATGACTGAACGTCCGATAA | 310 bp | mecA gene | 52 °C | [46] |
| mecA-R | CCAATTCCACATTGTTCGGTCTAA | ||||
| 5′-CS | GGCATACAAGCAGCAAGC | Variable | Intg I gene cassette(s) | 49 °C | [47] |
| 3′-CS | AAGCAGACTTGACCTGAT | ||||
| Ti-F | ACCTTTTTGTCGCATATCCGTG | Variable | Intg II gene cassette(s) | 52 °C | |
| TI-B | CTAACGCTTGAGTTAAGCC | ||||
| aacA4F | GCTCTTGGAAGCGGGGACGG | 300 bp | aacA4 gene | 54 °C | [48] |
| aacA4R | TCGCTCGAATGCCTGGCGTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Baz, A.M.; Yahya, G.; Mansour, B.; El-Sokkary, M.M.A.; Alshaman, R.; Alattar, A.; El-Ganiny, A.M. The Link between Occurrence of Class I Integron and Acquired Aminoglycoside Resistance in Clinical MRSA Isolates. Antibiotics 2021, 10, 488. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050488
El-Baz AM, Yahya G, Mansour B, El-Sokkary MMA, Alshaman R, Alattar A, El-Ganiny AM. The Link between Occurrence of Class I Integron and Acquired Aminoglycoside Resistance in Clinical MRSA Isolates. Antibiotics. 2021; 10(5):488. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050488
Chicago/Turabian StyleEl-Baz, Ahmed M., Galal Yahya, Basem Mansour, Mohamed M. A. El-Sokkary, Reem Alshaman, Abdullah Alattar, and Amira M. El-Ganiny. 2021. "The Link between Occurrence of Class I Integron and Acquired Aminoglycoside Resistance in Clinical MRSA Isolates" Antibiotics 10, no. 5: 488. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050488