
Retrospective Data Insight into the Global Distribution of Carbapenemase-Producing Pseudomonas aeruginosa

 ,
,
Abstract
:1. Introduction
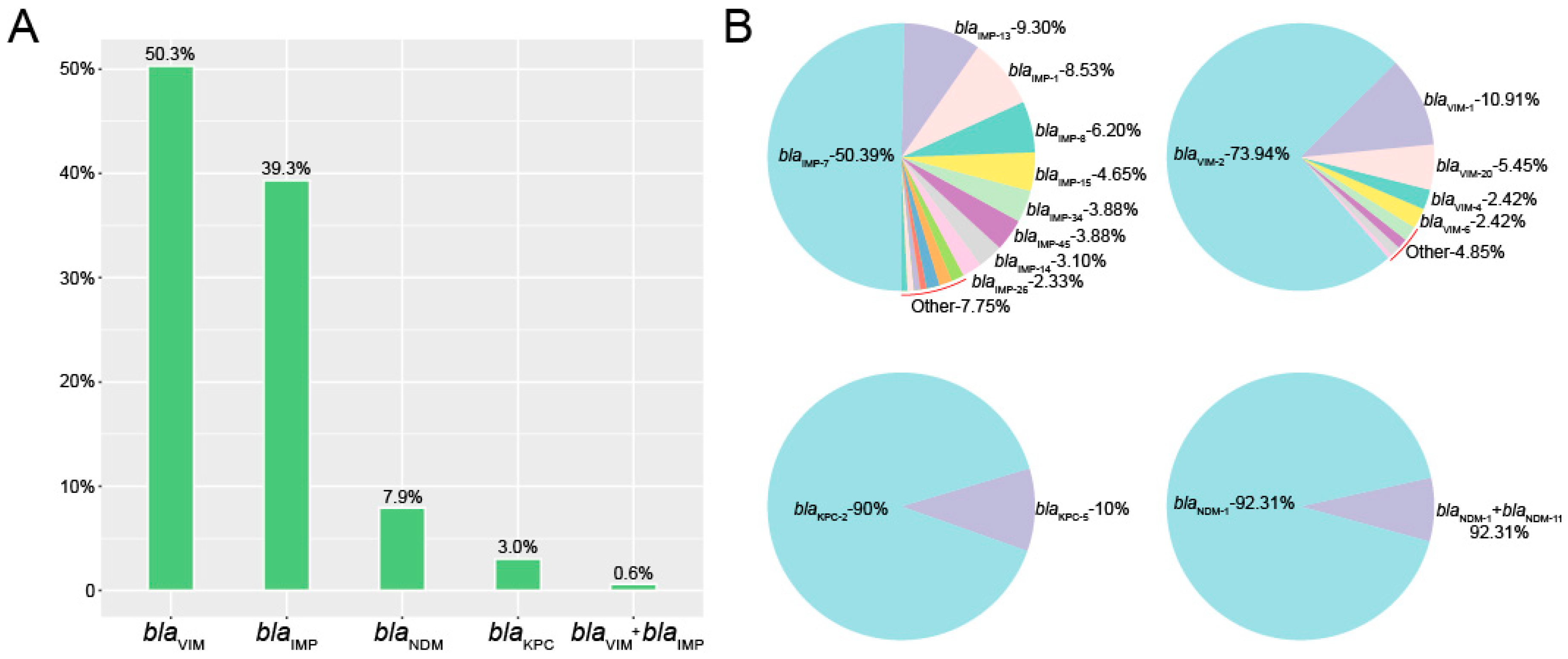
2. Results
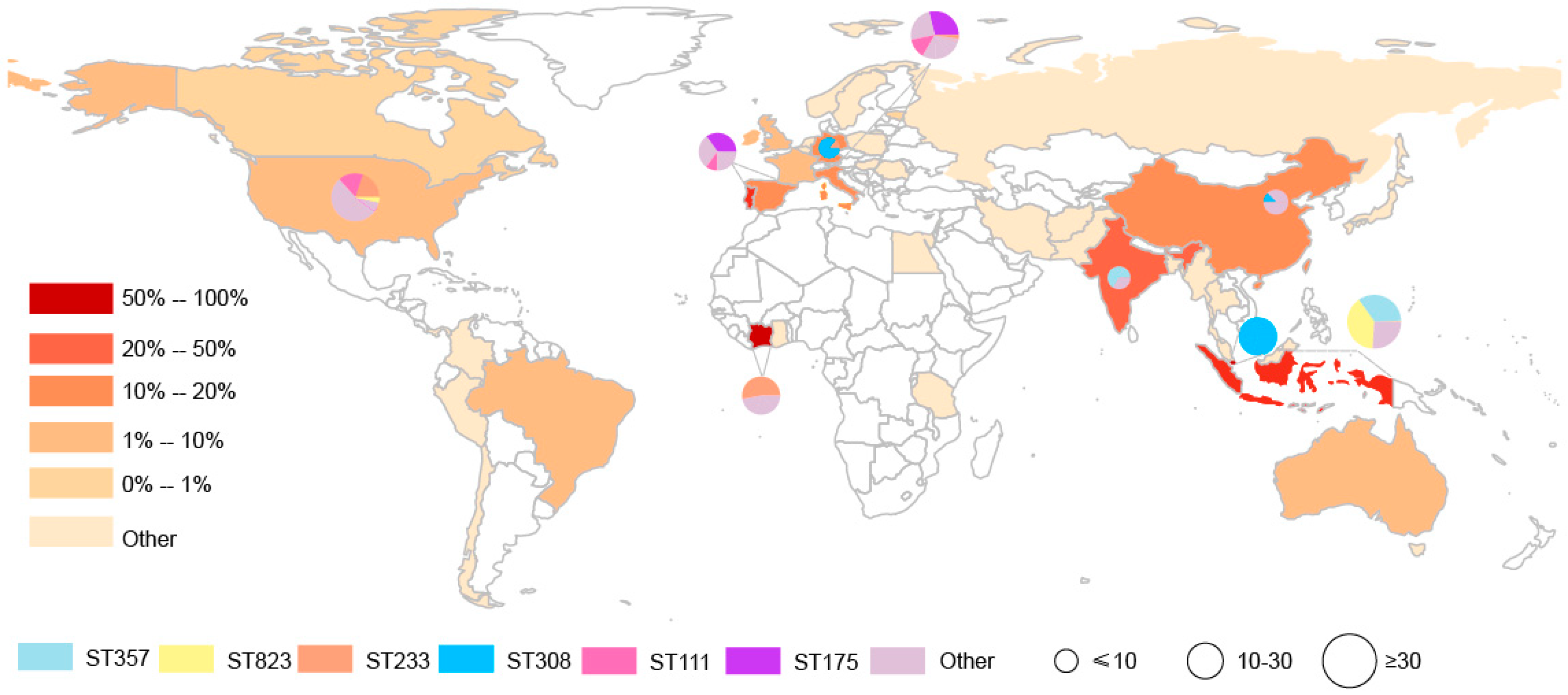
2.1. Geographical Distribution of the Carbapenemase-Producing P. aeruginosa Identified from the Genome Database
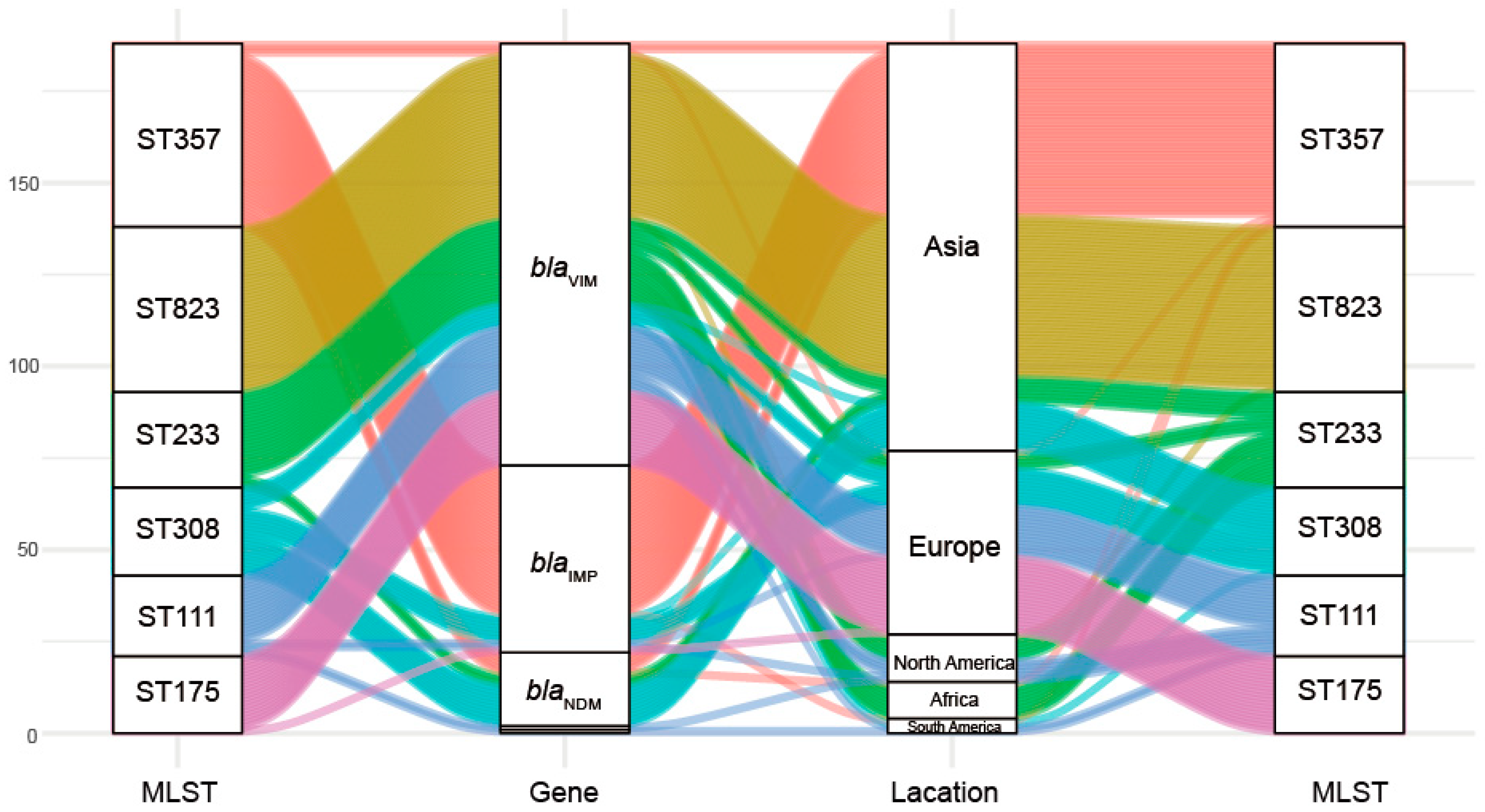
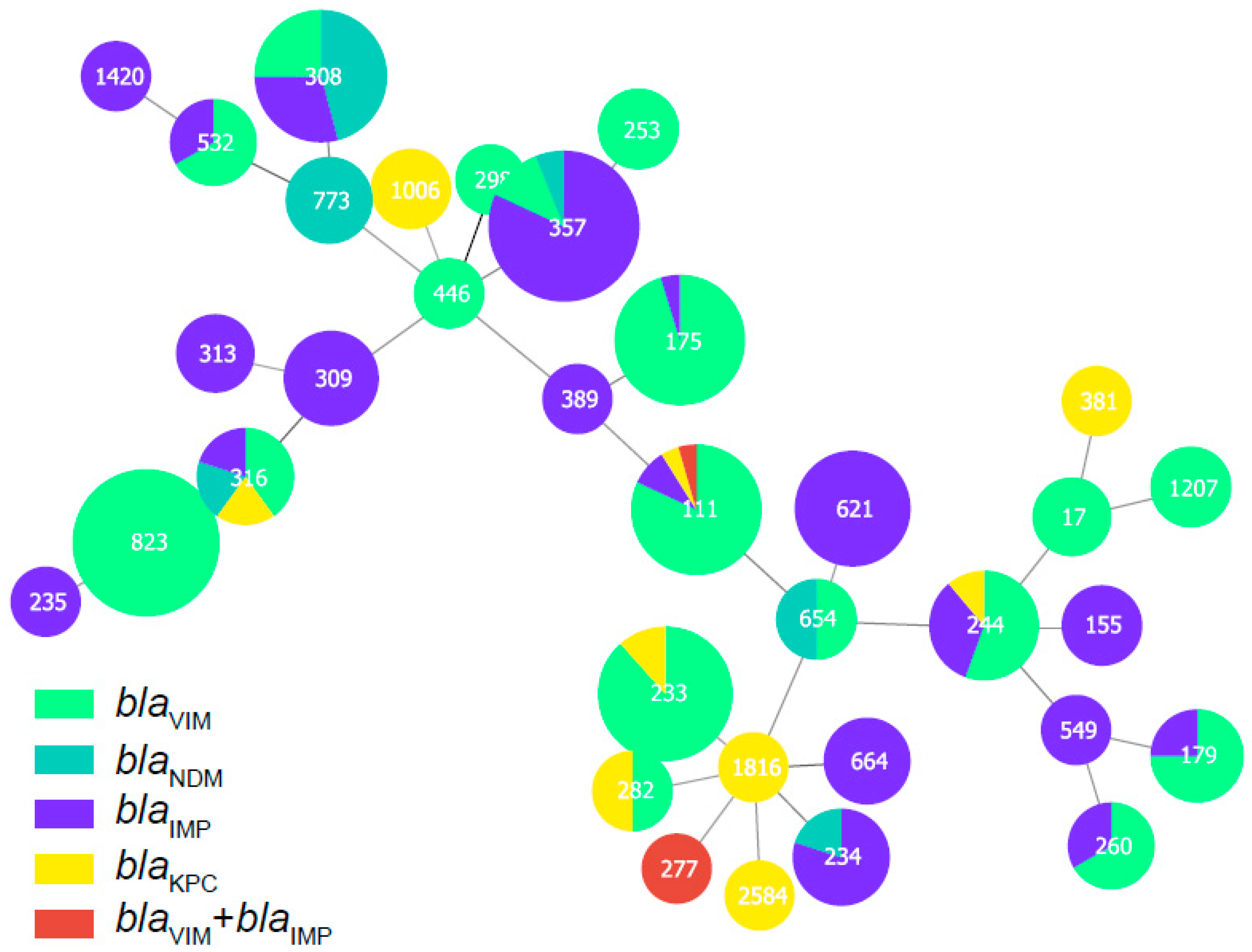
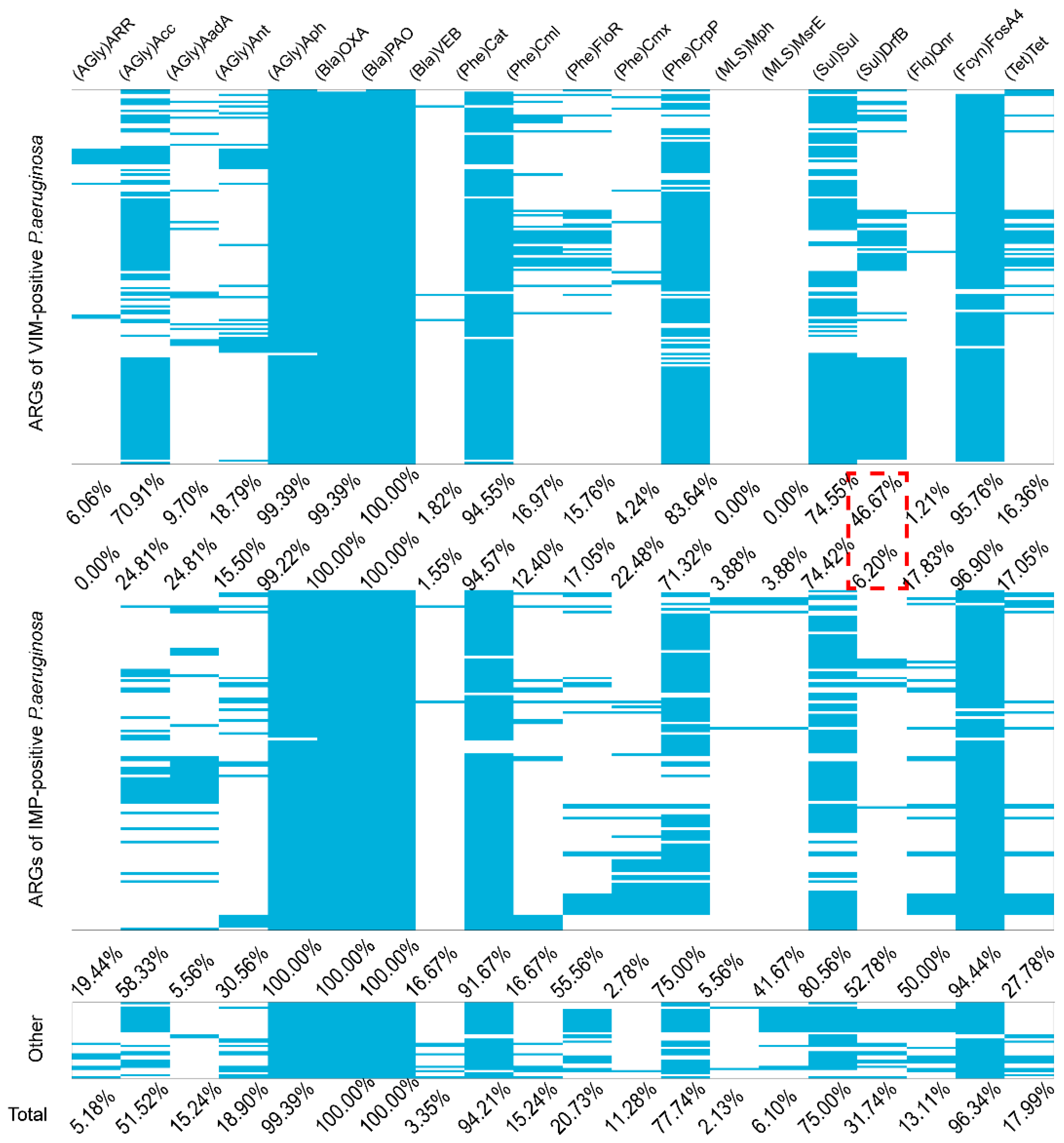
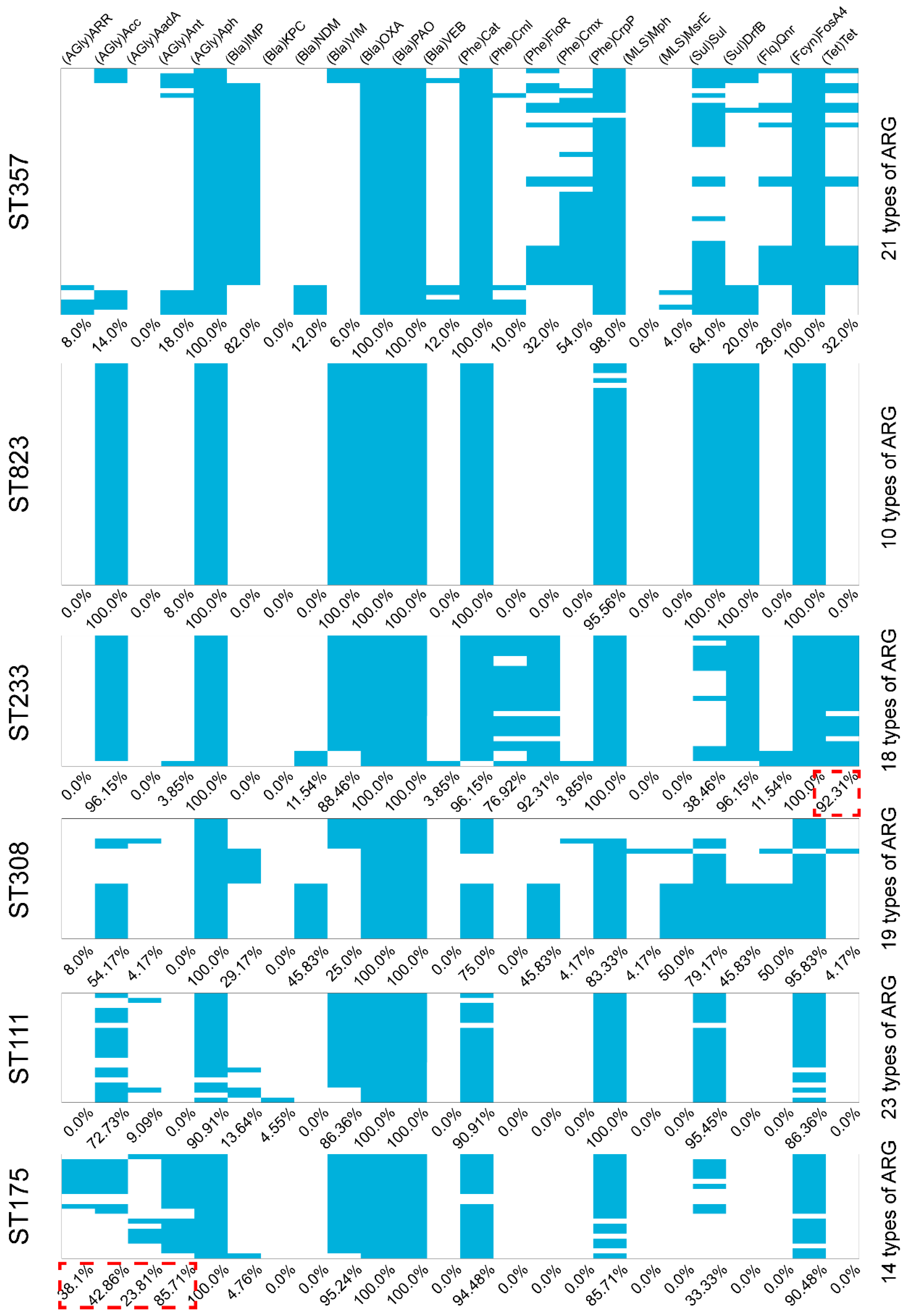
2.2. Molecular Characterization of the Carbapenemase-Producing P. aeruginosa
2.3. Other ARGs
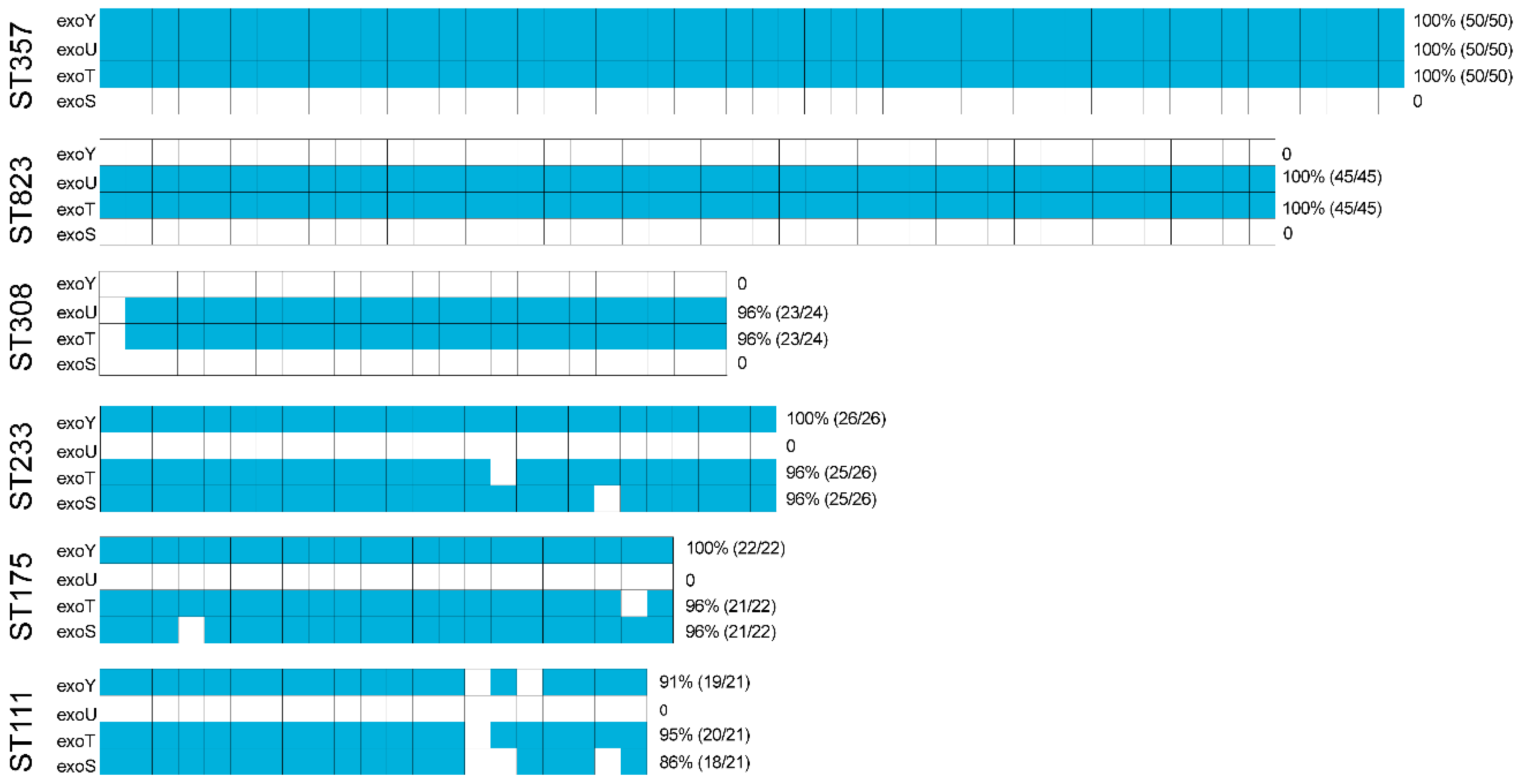
2.4. Virulence Factor
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Continents | Countries | Carbapenemase-Positive P. aeruginosa | Total of P. aeruginosa | Rate |
|---|---|---|---|---|
| Asia (162) | Indonesia | 112 | 222 | 50.45% |
| China | 8 | 54 | 14.81% | |
| Iran | 1 | 1 | ||
| Japan | 3 | 6 | ||
| Afghanistan | 2 | 3 | ||
| India | 9 | 39 | 23.08% | |
| Bangladesh | 1 | 1 | ||
| Myanmar | 1 | 1 | ||
| Turkey | 2 | 2 | ||
| Singapore | 11 | 11 | ||
| Malaysia | 4 | 16 | ||
| Lebanon | 4 | 8 | ||
| Thailand | 3 | 25 | ||
| Pakistan | 1 | 1 | ||
| Europe (101) | Russia | 2 | 9 | |
| Netherlands | 2 | 6 | ||
| Italy | 45 | 254 | 17.72% | |
| Portugal | 9 | 16 | ||
| Germany | 7 | 61 | 11.48% | |
| Norway | 2 | 2 | ||
| Estonia | 1 | 148 | 0.68% | |
| France | 5 | 65 | 7.69% | |
| Hungary | 1 | 1 | ||
| United Kingdom | 3 | 181 | 1.66% | |
| Belgium | 1 | 9 | ||
| Poland | 1 | 2 | ||
| Spain | 20 | 177 | 11.30% | |
| Sweden | 2 | 6 | ||
| America (40) | Brazil | 4 | 87 | 4.60% |
| Colombia | 1 | 3 | ||
| Peru | 2 | 2 | ||
| Chile | 1 | 2 | ||
| Panama | 1 | 1 | ||
| USA | 30 | 767 | 3.91% | |
| Canada | 1 | 497 | 0.20% | |
| Africa (23) | Ghana | 4 | 5 | |
| Cote d’Ivoire | 17 | 18 | ||
| Egypt | 1 | 1 | ||
| Tanzania | 1 | 16 | ||
| Oceania (2) | Australia | 2 | 117 | 1.71% |
| VIM | IMP | ||||
|---|---|---|---|---|---|
| Countries | Numbers | Rate | Countries | Numbers | Rate |
| Indonesia | 45 | 27.3% | Indonesia | 67 | 51.9% |
| Italy | 31 | 18.8% | Italy | 13 | 10.1% |
| USA | 19 | 11.5% | China | 7 | 5.4% |
| Spain | 16 | 9.7% | cote | 6 | 4.7% |
| cote | 11 | 6.7% | Germany | 6 | 4.7% |
| Portugal | 11 | 6.7% | Ghana | 4 | 3.1% |
| India | 5 | 3.0% | Malaysia | 4 | 3.1% |
| Lebanon | 4 | 2.4% | Spain | 4 | 3.1% |
| United Kingdom | 3 | 1.8% | USA | 4 | 3.1% |
| Afghanistan | 2 | 1.2% | France | 3 | 2.3% |
| Netherlands | 2 | 1.2% | Australia | 2 | 1.6% |
| Norway | 2 | 1.2% | Peru | 2 | 1.6% |
| Russia | 2 | 1.2% | Thailand | 2 | 1.6% |
| Brazil | 1 | 0.6% | Iran | 1 | 0.8% |
| Chile | 1 | 0.6% | Sweden | 1 | 0.8% |
| China | 1 | 0.6% | Tokyo | 1 | 0.8% |
| Colombia | 1 | 0.6% | Turkey | 1 | 0.8% |
| Egypt | 1 | 0.6% | Belgium | 1 | 0.8% |
| France | 1 | 0.6% | |||
| Germany | 1 | 0.6% | |||
| Panama | 1 | 0.6% | |||
| Peru | 1 | 0.6% | |||
| Sweden | 1 | 0.6% | |||
| Turkey | 1 | 0.6% | |||
| Bangladesh | 1 | 0.6% | |||
| Body Site | Number of Isolates | Rate |
|---|---|---|
| Throat | 42 | 17.50% |
| Urine | 39 | 16.25% |
| Rectal | 38 | 15.83% |
| Blood | 36 | 15.00% |
| Bronchial | 32 | 13.33% |
| Sputum | 27 | 11.25% |
| Wound | 11 | 4.58% |
| Abscess | 4 | 1.67% |
| Cornea | 2 | 0.83% |
| Feces | 2 | 0.83% |
| Lung | 2 | 0.83% |
| Eye | 1 | 0.42% |
| Groin | 1 | 0.42% |
| Peritoneal fluid | 1 | 0.42% |
| Pleural fluid | 1 | 0.42% |
| Knee | 1 | 0.42% |
| Unknown | 88 | 26.83% |
| MLST | Number of Carbapenemase-Positive P. aeruginosa |
|---|---|
| Unknown | 66 |
| ST357 | 50 |
| ST823 | 45 |
| ST233 | 26 |
| ST308 | 24 |
| ST111 | 22 |
| ST175 | 21 |
| ST621 | 12 |
| ST244 | 9 |
| ST234 | 5 |
| ST316 | 5 |
| ST179 | 4 |
| ST260 | 3 |
| ST309 | 3 |
| ST532 | 3 |
| ST664 | 3 |
| ST773 | 3 |
| ST155 | 2 |
| ST253 | 2 |
| ST282 | 2 |
| ST654 | 2 |
| ST1207 | 2 |
| ST1006 | 2 |
| ST17 | 1 |
| ST235 | 1 |
| ST277 | 1 |
| ST298 | 1 |
| ST313 | 1 |
| ST381 | 1 |
| ST389 | 1 |
| ST446 | 1 |
| ST549 | 1 |
| ST1420 | 1 |
| ST1816 | 1 |
| ST2584 | 1 |




References
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef]
- Hawkey, J.; Le Hello, S.; Doublet, B.; Granier, S.A.; Hendriksen, R.S.; Fricke, W.F.; Ceyssens, P.-J.; Gomart, C.; Billman-Jacobe, H.; Holt, K.E.; et al. Global phylogenomics of multidrug-resistant Salmonella enterica serotype Kentucky ST198. Microb. Genom. 2019, 5, e000269. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Hawkey, J.; Hetland, M.A.K.; Fostervold, A.; Wick, R.R.; Judd, L.M.; Hamidian, M.; Howden, B.P.; Löhr, I.H.; Holt, K.E. Emergence and rapid global dissemination of CTX-M-15-associated Klebsiella pneumoniae strain ST307. J. Antimicrob. Chemother. 2019, 74, 577–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; Chen, Y.; Wang, J.; Xie, X.; Ruan, Z. Emerging challenges of whole-genome-sequencing-powered epidemiological surveillance of globally distributed clonal groups of bacterial infections, giving Acinetobacter baumannii ST195 as an example. Int. J. Med. Microbiol. 2019, 309, 151339. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, X.-L.; Huang, A.-W.; Liu, S.-L.; Liu, W.-J.; Zhang, N.; Lu, X.-Z. Mortality attributable to carbapenem-resistant Pseudomonas aeruginosa bacteremia: A meta-analysis of cohort studies. Emerg. Microbes Infect. 2016, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Papagiannitsis, C.C.; Medvecky, M.; Chudejova, K.; Skalova, A.; Rotova, V.; Spanelova, P.; Jakubu, V.; Zemlickova, H.; Hrabak, J. Molecular Characterization of Carbapenemase-Producing Pseudomonas aeruginosa of Czech Origin and Evidence for Clonal Spread of Extensively Resistant Sequence Type 357 Expressing IMP-7 Metallo-β-Lactamase. Antimicrob. Agents Chemother. 2017, 61, e01811-17. [Google Scholar] [CrossRef] [Green Version]
- Patrice, N.; Poirel, L. Epidemiology and Diagnostics of Carbapenem Resistance in Gram-negative Bacteria. Clin. Infect. Dis. 2019, 69, S521–S528. [Google Scholar] [CrossRef]
- McCracken, M.G.; Adam, H.J.; Blondeau, J.M.; Walkty, A.J.; Karlowsky, J.A.; Hoban, D.J.; Zhanel, G.G.; Mulvey, M.R.; Baxter, M.R.; Nichol, K.A.; et al. Characterization of carbapenem-resistant and XDR Pseudomonas aeruginosa in Canada: Results of the CANWARD 2007–16 study. J. Antimicrob. Chemother. 2019, 74, iv32–iv38. [Google Scholar] [CrossRef]
- Hu, Y.-Y.; Cao, J.-M.; Yang, Q.; Chen, S.; Lv, H.-Y.; Zhou, H.-W.; Wu, Z.; Zhang, R. Risk Factors for Carbapenem-Resistant Pseudomonas aeruginosa, Zhejiang Province, China. Emerg. Infect. Dis. 2019, 25, 1861–1867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, J.W.P.; La, M.-V.; Jureen, R. Emergence of a New Delhi metallo-beta-lactamase-1-producing Pseudomonas aeruginosa in Singapore. Emerg. Microbes Infect. 2015, 4, e72. [Google Scholar] [CrossRef] [PubMed]
- Walkty, A.; Alexander, D.C.; Karlowsky, J.A.; Nichol, K.; Embil, J. Report of a KPC-producing Pseudomonas aeruginosa isolate in Canada. J. Antimicrob. Chemother. 2019, 74, 1748–1749. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, J.B.; Pfennigwerth, N.; Gatermann, S.G.; Von Baum, H.; Essig, A. KPC-2 carbapenemase-producing Pseudomonas aeruginosa reaching Germany. J. Antimicrob. Chemother. 2018, 73, 1812–1814. [Google Scholar] [CrossRef] [PubMed]
- Del Barrio-Tofiño, E.; López-Causapé, C.; Cabot, G.; Rivera, A.; Benito, N.; Segura, C.; Montero, M.M.; Sorlí, L.; Tubau, F.; Gómez-Zorrilla, S.; et al. Genomics and Susceptibility Profiles of Extensively Drug-Resistant Pseudomonas aeruginosa Isolates from Spain. Antimicrob. Agents Chemother. 2017, 61, 01589-17. [Google Scholar] [CrossRef] [Green Version]
- Walters, M.S.; Grass, J.E.; Bulens, S.N.; Hancock, E.B.; Phipps, E.C.; Muleta, D.; Mounsey, J.; Kainer, M.A.; Concannon, C.; Dumyati, G.; et al. Carbapenem-Resistant Pseudomonas aeruginosa at US Emerging Infections Program Sites, 2015. Emerg. Infect. Dis. 2019, 25, 1281–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labarca, J.A.; Salles, M.J.C.; Seas, C.; Guzmán-Blanco, M. Carbapenem resistance in Pseudomonas aeruginosa and Acinetobacter baumanniiin the nosocomial setting in Latin America. Crit. Rev. Microbiol. 2014, 42, 1–17. [Google Scholar] [CrossRef]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and Their Bacterial Producers in Health Care Settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Dong, N.; Chan, E.W.-C.; Zhang, R.; Chen, S. Carbapenem Resistance-Encoding and Virulence-Encoding Conjugative Plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021, 29, 65–83. [Google Scholar] [CrossRef]
- Saharman, Y.R.; Pelegrin, A.C.; Karuniawati, A.; Sedono, R.; Aditianingsih, D.; Goessens, W.H.; Klaassen, C.H.; Van Belkum, A.; Mirande, C.; Verbrugh, H.A.; et al. Epidemiology and characterisation of carbapenem-non-susceptible Pseudomonas aeruginosa in a large intensive care unit in Jakarta, Indonesia. Int. J. Antimicrob. Agents 2019, 54, 655–660. [Google Scholar] [CrossRef]
- Rossolini, G.M.; Riccio, M.L.; Cornaglia, G.; Pagani, L.; Lagatolla, C.; Selan, L.; Fontana, R. Carbapenem-Resistant Pseudomonas aeruginosa with Acquired blavim Metallo-β-Lactamase Determinants, Italy. Emerg. Infect. Dis. 2000, 6, 312–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viedma, E.; Juan, C.; Villa, J.; Barrado, L.; Orellana, M.; Sanz, F.; Otero, J.R.; Oliver, A.; Chaves, F. VIM-2–producing Multidrug-Resistant Pseudomonas aeruginosa ST175 Clone, Spain. Emerg. Infect. Dis. 2012, 18, 1235–1241. [Google Scholar] [CrossRef]
- Oliver, A.; Mulet, X.; López-Causapé, C.; Juan, C. The increasing threat of Pseudomonas aeruginosa high-risk clones. Drug Resist. Updat. 2015, 21–22, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Pelegrin, A.C.; Saharman, Y.R.; Griffon, A.; Palmieri, M.; Mirande, C.; Karuniawati, A.; Sedono, R.; Aditianingsih, D.; Goessens, W.H.F.; Van Belkum, A.; et al. High-Risk International Clones of Carbapenem-Nonsusceptible Pseudomonas aeruginosa Endemic to Indonesian Intensive Care Units: Impact of a Multifaceted Infection Control Intervention Analyzed at the Genomic Level. MBio 2019, 10, 02384-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rimoin, A.W.; Kisalu, N.; Kebela-Ilunga, B.; Mukaba, T.; Wright, L.L.; Formenty, P.; Wolfe, N.D.; Shongo, R.L.; Tshioko, F.; Okitolonda, E.; et al. Endemic Human Monkeypox, Democratic Republic of Congo, 2001–2004. Emerg. Infect. Dis. 2007, 13, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Horcajada, J.P.; Montero, M.; Oliver, A.; Sorlí, L.; Luque, S.; Gómez-Zorrilla, S.; Benito, N.; Grau, S. Epidemiology and Treatment of Multidrug-Resistant and Extensively Drug-Resistant Pseudomonas aeruginosa Infections. Clin. Microbiol. Rev. 2019, 32, 00031-19. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.L.; Octavia, S.; Ng, O.T.; Marimuthu, K.; Venkatachalam, I.; Cheng, B.; Lin, R.T.P.; Teo, J.W.P. Challenge of drug resistance in Pseudomonas aeruginosa: Clonal spread of NDM-1-positive ST-308 within a tertiary hospital. J. Antimicrob. Chemother. 2019, 74, 2220–2224. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Schwengers, O.; Schmiedel, J.; Baars, C.; Lambrecht, O.; Heß, S.; Berendonk, T.U.; Falgenhauer, J.; Chakraborty, T.; Imirzalioglu, C. Multidrug-Resistant and Clinically Relevant Gram-Negative Bacteria Are Present in German Surface Waters. Front. Microbiol. 2019, 10, 2779. [Google Scholar] [CrossRef]
- Zhai, R.; Fu, B.; Shi, X.; Sun, C.; Liu, Z.; Wang, S.; Shen, Z.; Walsh, T.R.; Cai, C.; Wang, Y.; et al. Contaminated in-house environment contributes to the persistence and transmission of NDM-producing bacteria in a Chinese poultry farm. Environ. Int. 2020, 139, 105715. [Google Scholar] [CrossRef]
- Cen, D.-J.; Sun, R.-Y.; Mai, J.-L.; Jiang, Y.-W.; Wang, D.; Guo, W.-Y.; Jiang, Q.; Zhang, H.; Zhang, J.-F.; Zhang, R.-M.; et al. Occurrence and Transmission of blaNDM-Carrying Enterobacteriaceae from Geese and the Surrounding Environment on a Commercial Goose Farm. Appl. Environ. Microbiol. 2021, 87, 00087-21. [Google Scholar] [CrossRef]
- Peña, C.; Cabot, G.; Gómez-Zorrilla, S.; Zamorano, L.; Ocampo-Sosa, A.; Murillas, J.; Almirante, B.; Pomar, V.; Aguilar, M.; Granados, A.; et al. Influence of Virulence Genotype and Resistance Profile in the Mortality of Pseudomonas aeruginosa Bloodstream Infections. Clin. Infect. Dis. 2015, 60, 539–548. [Google Scholar] [CrossRef]
- Lusignani, L.S.; Presterl, E.; Zatorska, B.; Nest, M.V.D.; Diab-Elschahawi, M. Infection control and risk factors for acquisition of carbapenemase-producing enterobacteriaceae. A 5 year (2011–2016) case-control study. Antimicrob. Resist. Infect. Control 2020, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, M.; Sousa, A.; Ramirez, M.; Francisco, A.P.; Carriço, J.A.; Vaz, C. PHYLOViZ 2.0: Providing scalable data integration and visualization for multiple phylogenetic inference methods. Bioinformatics 2017, 33, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.-G.; Liu, Z.-Y.; Liao, X.-P.; Sun, R.-Y.; Li, R.-B.; Liu, Y.; Fang, L.-X.; Sun, J.; Liu, Y.-H.; Zhang, R.-M. Retrospective Data Insight into the Global Distribution of Carbapenemase-Producing Pseudomonas aeruginosa. Antibiotics 2021, 10, 548. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050548
Wang M-G, Liu Z-Y, Liao X-P, Sun R-Y, Li R-B, Liu Y, Fang L-X, Sun J, Liu Y-H, Zhang R-M. Retrospective Data Insight into the Global Distribution of Carbapenemase-Producing Pseudomonas aeruginosa. Antibiotics. 2021; 10(5):548. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050548
Chicago/Turabian StyleWang, Min-Ge, Zhi-Yong Liu, Xiao-Ping Liao, Ruan-Yang Sun, Run-Bo Li, Yan Liu, Liang-Xing Fang, Jian Sun, Ya-Hong Liu, and Rong-Min Zhang. 2021. "Retrospective Data Insight into the Global Distribution of Carbapenemase-Producing Pseudomonas aeruginosa" Antibiotics 10, no. 5: 548. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10050548