
Lipid Transfer Proteins (LTPs)—Structure, Diversity and Roles beyond Antimicrobial Activity

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Classification, Nomenclature and Structural Features
2.1. Conserved Domains and Motifs
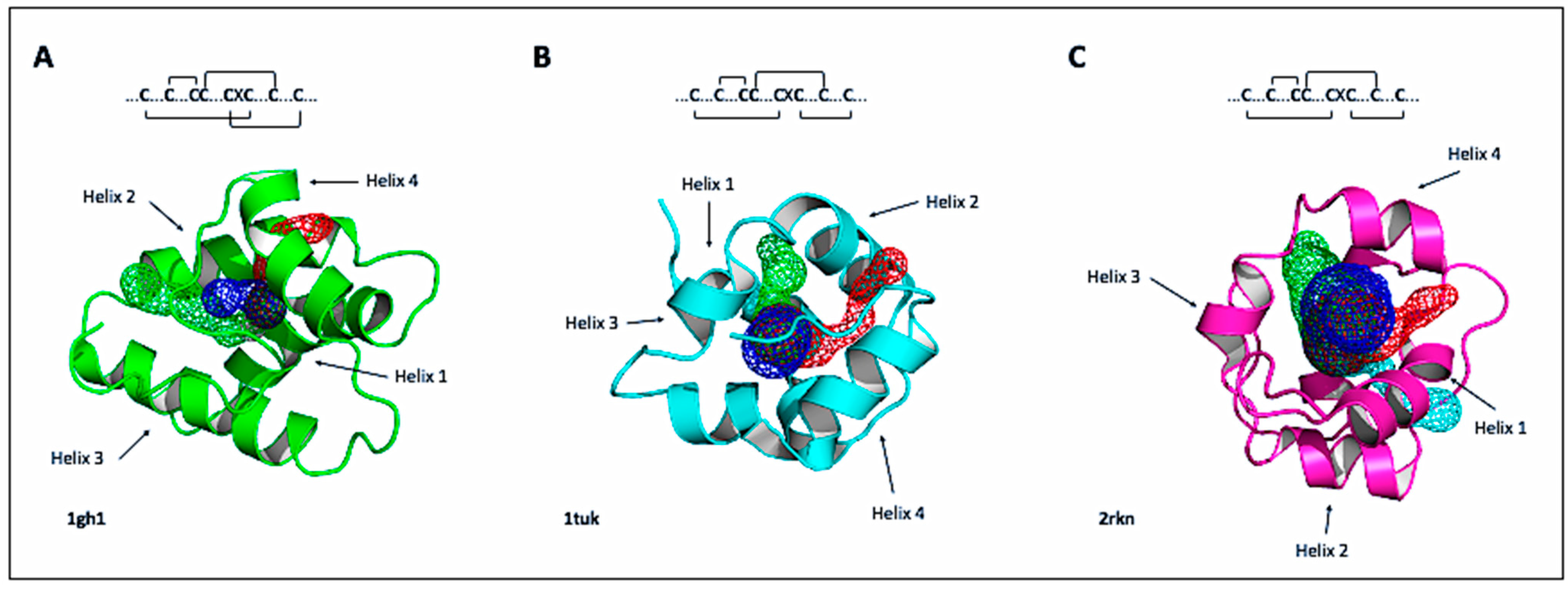
2.2. Tertiary Structure and Available Models
3. Roles of nsLTPs
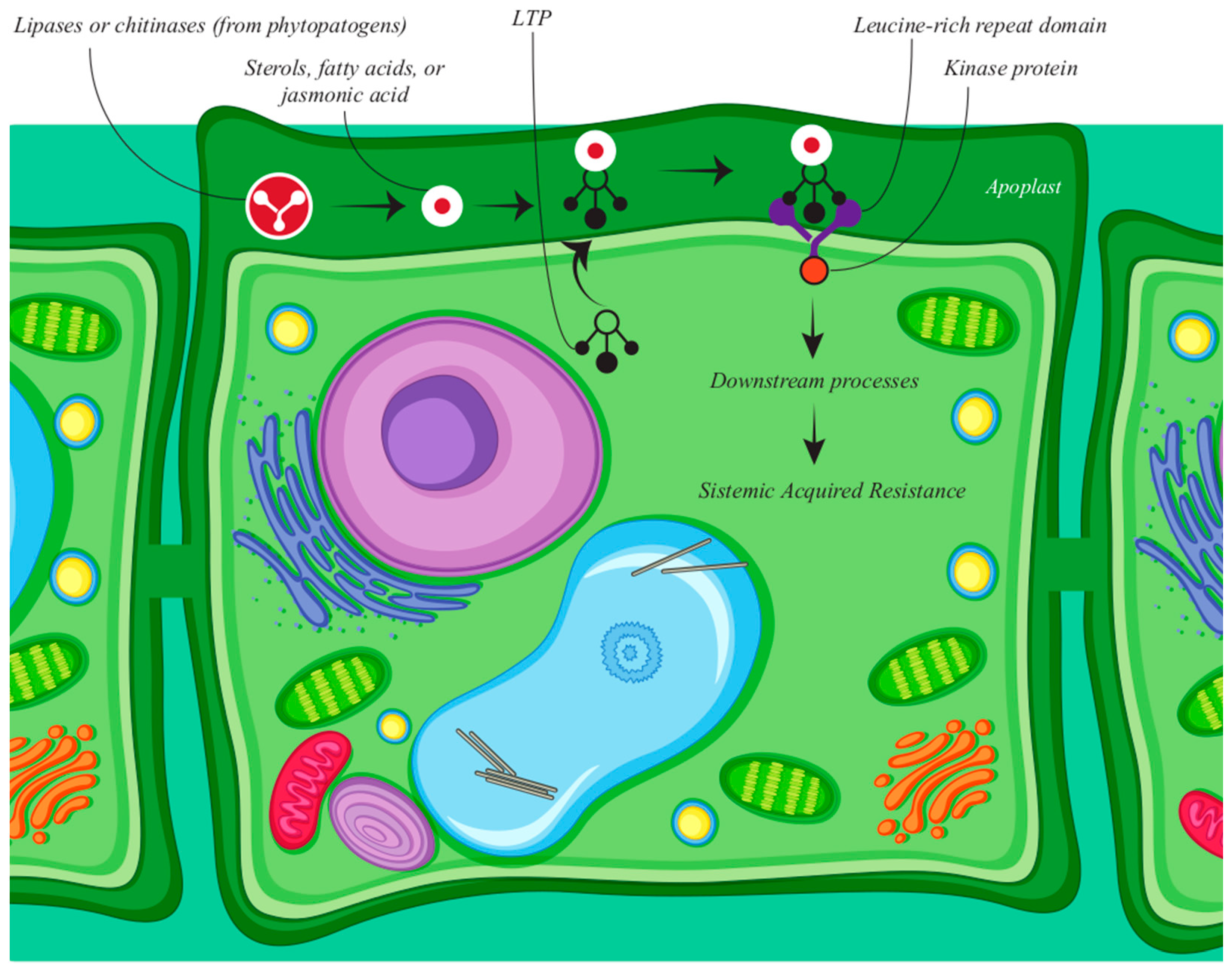
3.1. Defense
3.2. Growth, Development and Environmental Adaptation
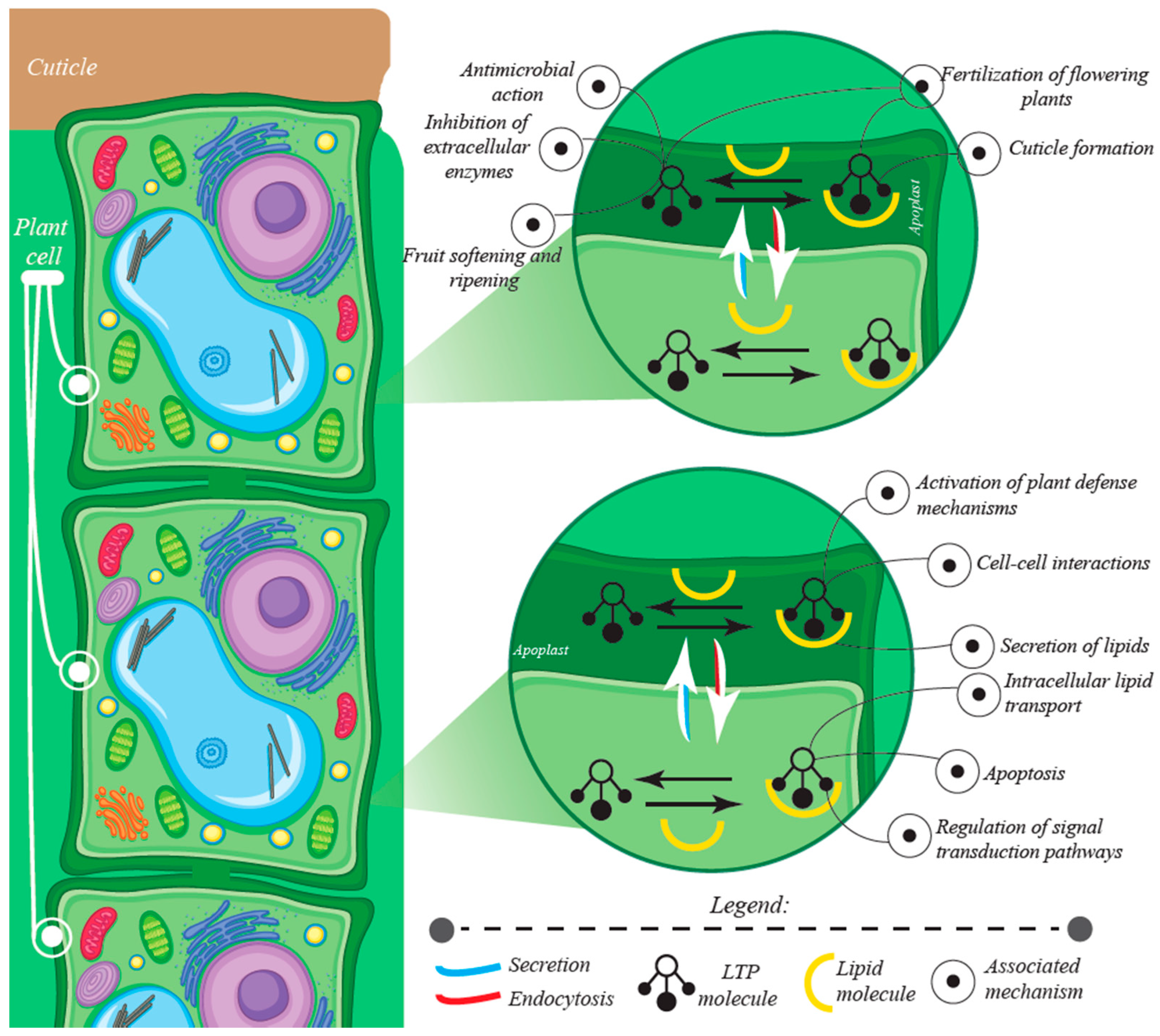
3.3. Other Roles
4. Genomic Distribution and Macrosynteny
5. Transcriptomics and Gene Modulation
6. Identification and Isolation
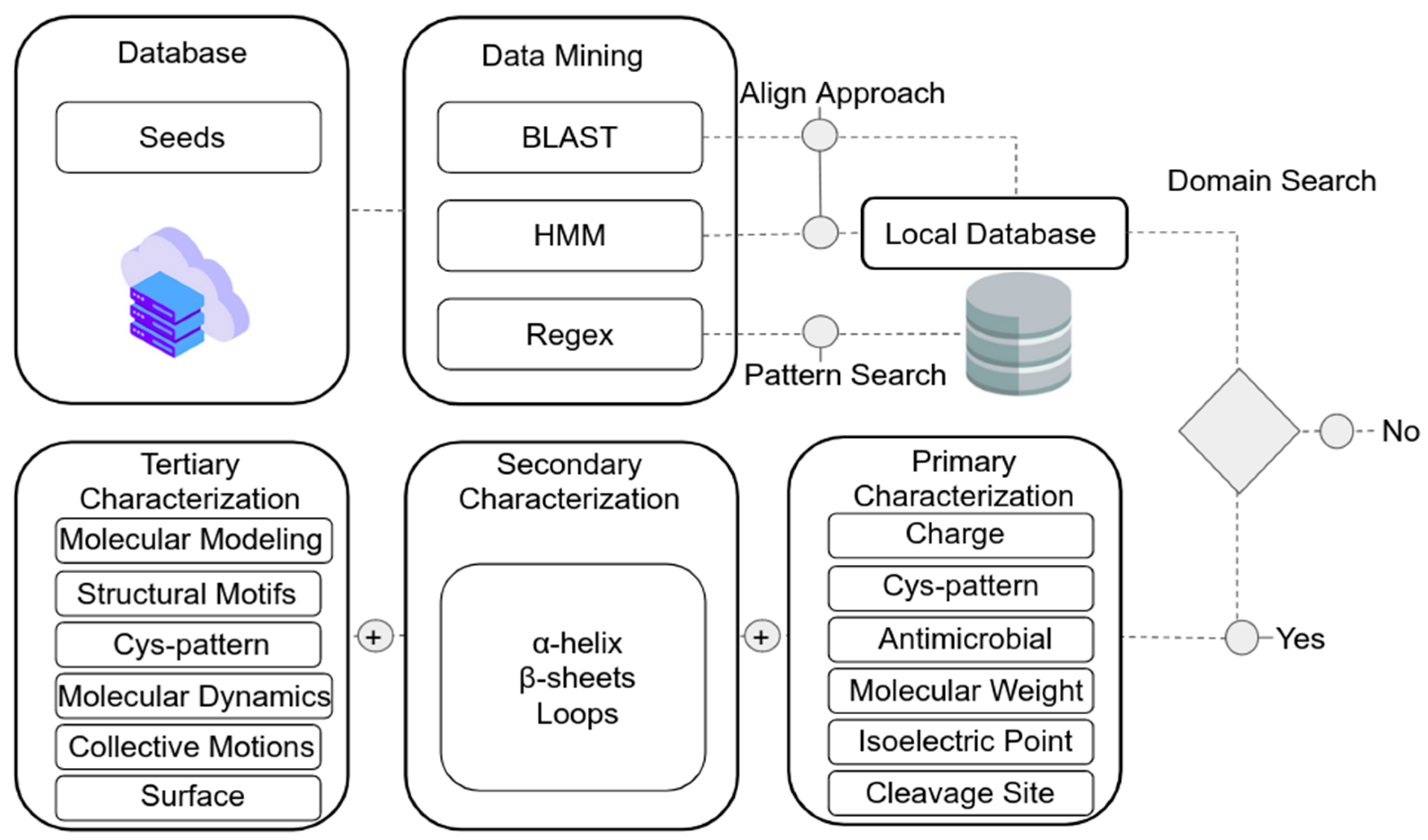
7. In Silico Identification of nsLTPs in Data Banks
8. Heterologous Expression and Biotechnology
8.1. Expression in Plants
8.2. Expression in Bacteria
8.3. Expression in Yeast
8.4. Purification and Yield of Expressed Products
9. PLTPs as Antimicrobial Agents
9.1. Biotechnological Applications on Plant Defense
9.1.1. In Vitro Studies
9.1.2. In Vivo Studies
9.2. Biotechnology Applied to Human Health
10. Toxicity, Allergenicity and Side Effects
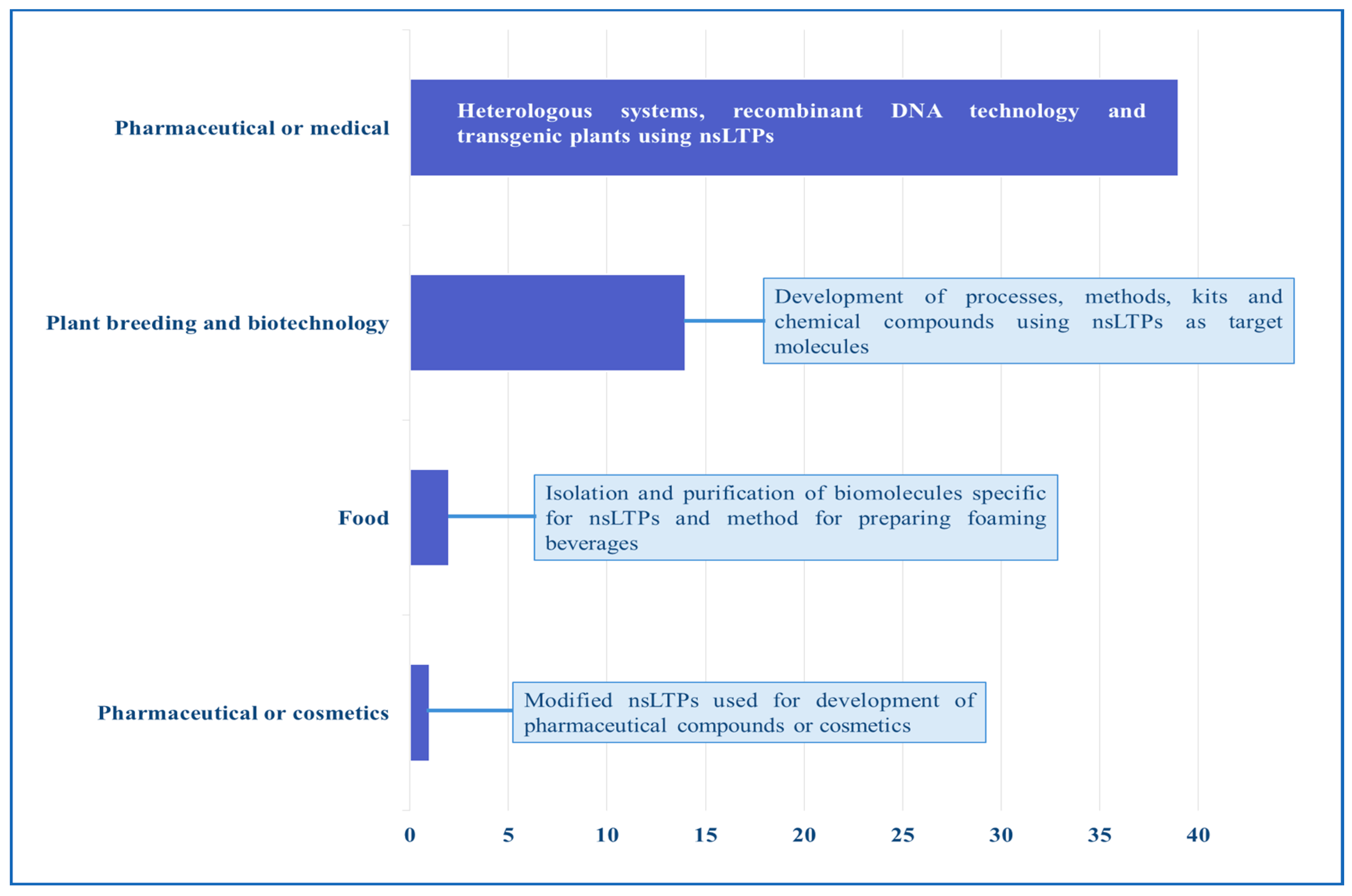
11. Patents and Products
12. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| pLTPs | Plant lipid transfer proteins |
| nsLTPs | Non-specific lipid transfer proteins |
| AMPs | Antimicrobial peptides |
| PR-proteins | Pathogenesis-related proteins |
| LRR | Rich domain in leucine |
| PK | Kinase protein |
| SAR | Systemic acquired resistance |
| BID | Pro-apoptotic protein |
| IAA | Acetic acid-2-indole |
| ABA | Abscisic acid |
| JA | Jasmonic acid |
| kDa | Kilo dalton |
| Cys | Cysteine |
| H | Helix |
| L | Loop |
| PCR | Polymerase chain reaction |
| RP-HPLC | Reverse-phase high-performance liquid chromatography |
| μM | Micromolar |
| pI | Isoelectric point |
| pH | Hydrogen potential |
| MAPK | Mitogen-activated protein kinase |
| ROS | Oxygen-reactive species |
| GST | Glutathione s-transferase |
| MBP | Maltose binding protein |
| NHX1 | Sodium/hydrogen exchanger agent |
| HKT1 | Sodium carrier |
| AINEs | Non-steroidal antibiotics |
| USPTO | United States Patent and Trademark Office |
References
- Kader, J.-C. Proteins and the intracellular exchange of lipids. I. Stimulation of phospholipid exchange between mitochondria and microsomal fractions by proteins isolated from potato tuber. Biochim. Biophys. Acta 1975, 380, 31–44. [Google Scholar] [CrossRef]
- Kader, J.-C. Lipid-Transfer Proteins in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 627–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edqvist, J.; Blomqvist, K.; Nieuwland, J.; Salminen, T.A. Plant Lipid Transfer Proteins: Are We Finally Closing in on the Roles of These Enigmatic Proteins? J. Lipid Res. 2018, 59, 1374–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A.; Korostyleva, T.V.; Kovtun, A.S.; Kasianov, A.S.; Shcherbakova, L.A.; Kudryavtsev, A.M. Non-specific lipid transfer proteins in Triticum kiharae Dorof. et Migush.: Identification, characterization and expression profiling in response to pathogens and resistance inducers. Pathogens 2019, 8, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, T.A.; Blomqvist, K.; Edqvist, J. Lipid Transfer Proteins: Classification, Nomenclature, Structure, and Function. Planta 2016, 244, 971–997. [Google Scholar] [CrossRef] [Green Version]
- José-Estanyol, M.; Gomis-Rüth, F.X.; Puigdomènech, P. The Eight-Cysteine Motif, a Versatile Structure in Plant Proteins. Plant Physiol. Biochem. 2004, 42, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Boutrot, F.; Guirao, A.; Alary, R.; Joudrier, P.; Gautier, M.-F. Wheat Non-Specific Lipid Transfer Protein Genes Display a Complex Pattern of Expression in Developing Seeds. Biochim. Biophys. Acta BBA-Gene Struct. Expr. 2005, 1730, 114–125. [Google Scholar] [CrossRef]
- Boutrot, F.; Chantret, N.; Gautier, M.-F. Genome-Wide Analysis of the Rice and Arabidopsis Non-Specific Lipid Transfer Protein (NsLtp) Gene Families and Identification of Wheat NsLtp Genes by EST Data Mining. BMC Genom. 2008, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Huang, D.; Liu, K.; Hu, S.; Yu, J.; Gao, G.; Song, S. Discovery, Identification and Comparative Analysis of Non-Specific Lipid Transfer Protein (NsLtp) Family in Solanaceae. Genom. Proteom. Bioinform. 2010, 8, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.W.; Hwang, S.-G.; Karuppanapandian, T.; Liu, A.; Kim, W.; Jang, C.S. Insight into the Molecular Evolution of Non-Specific Lipid Transfer Proteins via Comparative Analysis Between Rice and Sorghum. DNA Res. 2012, 19, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Tapia, G.; Morales-Quintana, L.; Parra, C.; Berbel, A.; Alcorta, M. Study of NsLTPs in Lotus japonicus Genome Reveal a Specific Epidermal Cell Member (LjLTP10) Regulated by Drought Stress in Aerial Organs with a Putative Role in Cutin Formation. Plant Mol. Biol. 2013, 82, 485–501. [Google Scholar] [CrossRef]
- Li, J.; Gao, G.; Xu, K.; Chen, B.; Yan, G.; Li, F.; Qiao, J.; Zhang, T.; Wu, X. Genome-Wide Survey and Expression Analysis of the Putative Non-Specific Lipid Transfer Proteins in Brassica rapa L. PLoS ONE 2014, 9, e84556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleury, C.; Gracy, J.; Gautier, M.-F.; Pons, J.-L.; Dufayard, J.-F.; Labesse, G.; Ruiz, M.; de Lamotte, F. Comprehensive Classification of the Plant Non-Specific Lipid Transfer Protein Superfamily towards Its Sequence–Structure–Function Analysis. PeerJ 2019, 7, e7504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edstam, M.M.; Viitanen, L.; Salminen, T.A.; Edqvist, J. Evolutionary History of the Non-Specific Lipid Transfer Proteins. Mol. Plant 2011, 4, 947–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gincel, E.; Simorre, J.-P.; Caille, A.; Marion, D.; Ptak, M.; Vovelle, F. Three-Dimensional Structure in Solution of a Wheat Lipid-Transfer Protein from Multidimensional 1H-NMR Data. Eur. J. Biochem. 1994, 226, 413–422. [Google Scholar] [CrossRef]
- Pons, J.-L.; de Lamotte, F.; Gautier, M.-F.; Delsuc, M.-A. Refined Solution Structure of a Liganded Type 2 Wheat Nonspecific Lipid Transfer Protein *. J. Biol. Chem. 2003, 278, 14249–14256. [Google Scholar] [CrossRef] [Green Version]
- Lascombe, M.-B.; Bakan, B.; Buhot, N.; Marion, D.; Blein, J.-P.; Larue, V.; Lamb, C.; Prangé, T. The Structure of “Defective in Induced Resistance” Protein of Arabidopsis thaliana, DIR1, Reveals a New Type of Lipid Transfer Protein. Protein Sci. 2008, 17, 1522–1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, R.K.; Mattoo, A.K. Plant Antimicrobial Peptides. In Host Defense Peptides and Their Potential as Therapeutic Agents; Epand, R.M., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 111–136. ISBN 978-3-319-32949-9. [Google Scholar]
- Simorre, J.P.; Caille, A.; Marion, D.; Marion, D.; Ptak, M. Two- and Three-Dimensional Proton NMR Studies of a Wheat Phospholipid Transfer Protein: Sequential Resonance Assignments and Secondary Structure. Biochemistry 1991, 30, 11600–11608. [Google Scholar] [CrossRef]
- Chovancova, E.; Pavelka, A.; Benes, P.; Strnad, O.; Brezovsky, J.; Kozlikova, B.; Gora, A.; Sustr, V.; Klvana, M.; Medek, P.; et al. CAVER 3.0: A Tool for the Analysis of Transport Pathways in Dynamic Protein Structures. PLoS Comput. Biol. 2012, 8, e1002708. [Google Scholar] [CrossRef] [Green Version]
- Sels, J.; Mathys, J.; De Coninck, B.M.A.; Cammue, B.P.A.; De Bolle, M.F.C. Plant Pathogenesis-Related (PR) Proteins: A Focus on PR Peptides. Plant Physiol. Biochem. 2008, 46, 941–950. [Google Scholar] [CrossRef]
- van der Weerden, N.L.; Bleackley, M.R.; Anderson, M.A. Properties and Mechanisms of Action of Naturally Occurring Antifungal Peptides. Cell. Mol. Life Sci. 2013, 70, 3545–3570. [Google Scholar] [CrossRef] [PubMed]
- Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Ovchinnikova, T.V. Lipid Transfer Proteins As Components of the Plant Innate Immune System: Structure, Functions, and Applications. Acta Nat. 2016, 8, 47–61. [Google Scholar] [CrossRef]
- Souza, A.A.; Costa, A.S.; Campos, D.C.O.; Batista, A.H.M.; Sales, G.W.P.; Nogueira, N.A.P.; Alves, K.M.M.; Coelho-de-Souza, A.N.; Oliveira, H.D. Lipid Transfer Protein Isolated from Noni Seeds Displays Antibacterial Activity in Vitro and Improves Survival in Lethal Sepsis Induced by CLP in Mice. Biochimie 2018, 149, 9–17. [Google Scholar] [CrossRef]
- Safi, H.; Saibi, W.; Alaoui, M.M.; Hmyene, A.; Masmoudi, K.; Hanin, M.; Brini, F. A Wheat Lipid Transfer Protein (TdLTP4) Promotes Tolerance to Abiotic and Biotic Stress in Arabidopsis thaliana. Plant Physiol. Biochem. 2015, 89, 64–75. [Google Scholar] [CrossRef]
- Tomassen, M.M.M.; Barrett, D.M.; van der Valk, H.C.P.M.; Woltering, E.J. Isolation and Characterization of a Tomato Non-Specific Lipid Transfer Protein Involved in Polygalacturonase-Mediated Pectin Degradation. J. Exp. Bot. 2007, 58, 1151–1160. [Google Scholar] [CrossRef] [Green Version]
- Chebli, Y.; Geitmann, A. Cellular Growth in Plants Requires Regulation of Cell Wall Biochemistry. Curr. Opin. Cell Biol. 2017, 44, 28–35. [Google Scholar] [CrossRef]
- Nieuwland, J.; Feron, R.; Huisman, B.A.H.; Fasolino, A.; Hilbers, C.W.; Derksen, J.; Mariani, C. Lipid Transfer Proteins Enhance Cell Wall Extension in Tobacco. Plant Cell 2005, 17, 2009–2019. [Google Scholar] [CrossRef] [Green Version]
- Hairat, S.; Baranwal, V.K.; Khurana, P. Identification of Triticum aestivum NsLTPs and Functional Validation of Two Members in Development and Stress Mitigation Roles. Plant Physiol. Biochem. 2018, 130, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Hou, W.; Du, X.; Wang, L.; Wu, H.; Zhao, L.; Kong, L.; Wang, H. Identification of Wheat Non-Specific Lipid Transfer Proteins Involved in Chilling Tolerance. Plant Cell Rep. 2014, 33, 1757–1766. [Google Scholar] [CrossRef]
- Fang, Z.; He, Y.; Liu, Y.; Jiang, W.; Song, J.; Wang, S.; Ma, D.; Yin, J. Bioinformatic Identification and Analyses of the Non-Specific Lipid Transfer Proteins in Wheat. J. Integr. Agric. 2020, 19, 1170–1185. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, X.; Song, Y.; Zhu, L.; Yu, Z.; Gan, L.; Zhou, S.; Liu, H.; Wen, F.; Zhu, C. NtLTP4, a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum. Sci. Rep. 2018, 8, 8873. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-Specific Lipid Transfer Proteins in Plants: Presenting New Advances and an Integrated Functional Analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edstam, M.M.; Edqvist, J. Involvement of GPI-Anchored Lipid Transfer Proteins in the Development of Seed Coats and Pollen in Arabidopsis thaliana. Physiol. Plant. 2014, 152, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Douliez, J.-P.; Michon, T.; Elmorjani, K.; Marion, D. Mini Review: Structure, Biological and Technological Functions of Lipid Transfer Proteins and Indolines, the Major Lipid Binding Proteins from Cereal Kernels. J. Cereal Sci. 2000, 32, 1–20. [Google Scholar] [CrossRef]
- Crimi, M.; Astegno, A.; Zoccatelli, G.; Esposti, M.D. Pro-Apoptotic Effect of Maize Lipid Transfer Protein on Mammalian Mitochondria. Arch. Biochem. Biophys. 2006, 445, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Degli Esposti, M. Sequence and Functional Similarities between Pro-Apoptotic Bid and Plant Lipid Transfer Proteins. Biochim. Biophys. Acta BBA-Bioenerg. 2002, 1553, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Blein, J.-P.; Coutos-Thévenot, P.; Marion, D.; Ponchet, M. From Elicitins to Lipid-Transfer Proteins: A New Insight in Cell Signalling Involved in Plant Defence Mechanisms. Trends Plant Sci. 2002, 7, 293–296. [Google Scholar] [CrossRef]
- Buhot, N.; Gomès, E.; Milat, M.-L.; Ponchet, M.; Marion, D.; Lequeu, J.; Delrot, S.; Coutos-Thévenot, P.; Blein, J.-P. Modulation of the Biological Activity of a Tobacco LTP1 by Lipid Complexation. Mol. Biol. Cell 2004, 15, 5047–5052. [Google Scholar] [CrossRef] [Green Version]
- Kouidri, A.; Baumann, U.; Okada, T.; Baes, M.; Tucker, E.J.; Whitford, R. Wheat TaMs1 Is a Glycosylphosphatidylinositol-Anchored Lipid Transfer Protein Necessary for Pollen Development. BMC Plant Biol. 2018, 18, 332. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Burns, J.K. Isolation and Characterization of a CDNA Encoding a Lipid Transfer Protein Expressed in ‘Valencia’ Orange during Abscission. J. Exp. Bot. 2003, 54, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Ohno, S. The Creation of a New Gene from a Redundant Duplicate of an Old Gene. In Evolution by Gene Duplication; Ohno, S., Ed.; Springer: Berlin/Heidelberg, Germany, 1970; pp. 71–82. ISBN 978-3-642-86659-3. [Google Scholar]
- Rensing, S.A. Gene Duplication as a Driver of Plant Morphogenetic Evolution. Curr. Opin. Plant Biol. 2014, 17, 43–48. [Google Scholar] [CrossRef]
- Desta, Z.A.; Ortiz, R. Genomic Selection: Genome-Wide Prediction in Plant Improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef]
- Arondel, V.; Vergnolle, C.; Cantrel, C.; Kader, J.-C. Lipid Transfer Proteins Are Encoded by a Small Multigene Family in Arabidopsis thaliana. Plant Sci. 2000, 157, 1–12. [Google Scholar] [CrossRef]
- Jang, C.S.; Jung, J.H.; Yim, W.C.; Lee, B.-M.; Seo, Y.W.; Kim, W. Divergence of Genes Encoding Non-Specific Lipid Transfer Proteins in the Poaceae Family. Mol. Cells 2007, 24, 215–223. [Google Scholar]
- Meng, C.; Yan, Y.; Liu, Z.; Chen, L.; Zhang, Y.; Li, X.; Wu, L.; Zhang, G.; Wang, X.; Ma, Z. Systematic Analysis of Cotton Non-Specific Lipid Transfer Protein Family Revealed a Special Group That Is Involved in Fiber Elongation. Front. Plant Sci. 2018, 9, 1285. [Google Scholar] [CrossRef]
- Jiao, Y.; Wickett, N.J.; Ayyampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral Polyploidy in Seed Plants and Angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef]
- Irigoyen, M.L.; Garceau, D.C.; Bohorquez-Chaux, A.; Lopez-Lavalle, L.A.B.; Perez-Fons, L.; Fraser, P.D.; Walling, L.L. Genome-Wide Analyses of Cassava Pathogenesis-Related (PR) Gene Families Reveal Core Transcriptome Responses to Whitefly Infestation, Salicylic Acid and Jasmonic Acid. BMC Genom. 2020, 21, 93. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, K.A.T.; Graham, M.A.; Paape, T.D.; VandenBosch, K.A. Genome Organization of More than 300 Defensin-like Genes in Arabidopsis. Plant Physiol. 2005, 138, 600–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Lv, Y.; Hou, Z.; Li, X.; Ding, L. Expansion and Evolution of Thaumatin-like Protein (TLP) Gene Family in Six Plants. Plant Growth Regul. 2016, 79, 299–307. [Google Scholar] [CrossRef]
- Wanderley-Nogueira, A.C.; Belarmino, L.C.; Soares-Cavalcanti, N.D.M.; Bezerra-Neto, J.P.; Kido, E.A.; Pandolfi, V.; Abdelnoor, R.V.; Binneck, E.; Carazzole, M.F.; Benko-Iseppon, A.M. An Overall Evaluation of the Resistance (R) and Pathogenesis-Related (PR) Superfamilies in Soybean, as Compared with Medicago and Arabidopsis. Genet. Mol. Biol. 2012, 35, 260–271. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, N.; Buonanno, M.; Ayoub, J.; Barone, A.; Monti, S.M.; Rigano, M.M. Identification of Non-Specific Lipid Transfer Protein Gene Family Members in Solanum lycopersicum and Insights into the Features of Sola l 3 Protein. Sci. Rep. 2019, 9, 1607. [Google Scholar] [CrossRef]
- Ji, J.; Lv, H.; Yang, L.; Fang, Z.; Zhuang, M.; Zhang, Y.; Liu, Y.; Li, Z. Genome-Wide Identification and Characterization of Non-Specific Lipid Transfer Proteins in Cabbage. PeerJ 2018, 6, e5379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Kim, Y.; Zong, J.; Lin, H.; Dievart, A.; Li, H.; Zhang, D.; Liang, W. Genome-wide analysis of the barley non-specific lipid transfer protein gene family. Crop J. 2019, 7, 65–76. [Google Scholar] [CrossRef]
- Zhu, X.; Li, Z.; Xu, H.; Zhou, M.; Du, L.; Zhang, Z. Overexpression of Wheat Lipid Transfer Protein Gene TaLTP5 Increases Resistances to Cochliobolus sativus and Fusarium graminearum in Transgenic Wheat. Funct. Integr. Genom. 2012, 12, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Kido, E.A.; Pandolfi, V.; Houllou-Kido, L.M.; Andrade, P.P.; Marcelino, F.C.; Nepomuceno, A.L.; Abdelnoor, R.V.; Benko-Iseppon, W.L.B.; Benko-Iseppon, A.M. Plant Antimicrobial Peptides: An Overview of SuperSAGE Transcriptional Profile and a Functional Review. Available online: https://www.eurekaselect.com/85038/article (accessed on 28 February 2021).
- Pii, Y.; Pandolfini, T.; Crimi, M. Signaling LTPs: A New Plant LTPs Sub-Family? Plant Signal. Behav. 2010, 5, 594–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Ma, J.; Zhang, X.; Yang, Y.; Zhou, D.; Yu, Q.; Que, Y.; Xu, L.; Guo, J. A Novel Non-Specific Lipid Transfer Protein Gene from Sugarcane (NsLTPs), Obviously Responded to Abiotic Stresses and Signaling Molecules of SA and MeJA. Sugar Tech. 2017, 19, 17–25. [Google Scholar] [CrossRef]
- Wei, K.; Zhong, X. Non-Specific Lipid Transfer Proteins in Maize. BMC Plant Biol. 2014, 14, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edstam, M.M.; Blomqvist, K.; Eklöf, A.; Wennergren, U.; Edqvist, J. Coexpression Patterns Indicate That GPI-Anchored Non-Specific Lipid Transfer Proteins Are Involved in Accumulation of Cuticular Wax, Suberin and Sporopollenin. Plant Mol. Biol. 2013, 83, 625–649. [Google Scholar] [CrossRef]
- Patkar, R.N.; Chattoo, B.B. Transgenic Indica Rice Expressing Ns-LTP-Like Protein Shows Enhanced Resistance to Both Fungal and Bacterial Pathogens. Mol. Breed. 2006, 17, 159–171. [Google Scholar] [CrossRef]
- Sohal, A.K.; Pallas, J.A.; Jenkins, G.I. The Promoter of a Brassica napus Lipid Transfer Protein Gene Is Active in a Range of Tissues and Stimulated by Light and Viral Infection in Transgenic Arabidopsis. Plant Mol. Biol. 1999, 41, 75–87. [Google Scholar] [CrossRef]
- Wang, X.; Li, Q.; Cheng, C.; Zhang, K.; Lou, Q.; Li, J.; Chen, J. Genome-Wide Analysis of a Putative Lipid Transfer Protein LTP_2 Gene Family Reveals CsLTP_2 Genes Involved in Response of Cucumber against Root-Knot Nematode (Meloidogyne incognita). Genome 2020, 63, 225–238. [Google Scholar] [CrossRef] [PubMed]
- da Silva, F.C.V.; do Nascimento, V.V.; Fernandes, K.V.; Machado, O.L.T.; da Silva Pereira, L.; Gomes, V.M.; de Oliveira Carvalho, A. Recombinant Production and α-Amylase Inhibitory Activity of the Lipid Transfer Protein from Vigna unguiculata (L. Walp.) Seeds. Process Biochem. 2018, 65, 205–212. [Google Scholar] [CrossRef]
- Van loon, L.C.; Van strien, E.A. The Families of Pathogenesis-Related Proteins, Their Activities, and Comparative Analysis of PR-1 Type Proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Sarowar, S.; Kim, Y.J.; Kim, K.D.; Hwang, B.K.; Ok, S.H.; Shin, J.S. Overexpression of Lipid Transfer Protein (LTP) Genes Enhances Resistance to Plant Pathogens and LTP Functions in Long-Distance Systemic Signaling in Tobacco. Plant Cell Rep. 2009, 28, 419–427. [Google Scholar] [CrossRef]
- Ke, T.; Cao, H.; Huang, J.; Hu, F.; Huang, J.; Dong, C.; Ma, X.; Yu, J.; Mao, H.; Wang, X.; et al. EST-Based in Silico Identification and in Vitro Test of Antimicrobial Peptides in brassica Napus. BMC Genom. 2015, 16, 653. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.S. Structural studies of antimicrobial peptides provide insight into their mechanisms of action. In Antimicrobial Peptides: Discovery, Design and Novel Therapeutic Strategies; Wang, G., Ed.; CABI: Wallingford, CT, USA, 2010; pp. 141–168. ISBN 978-1-84593-657-0. [Google Scholar]
- Jang, C.S.; Yim, W.C.; Moon, J.-C.; Jung, J.H.; Lee, T.G.; Lim, S.D.; Cho, S.H.; Lee, K.K.; Kim, W.; Seo, Y.W.; et al. Evolution of Non-Specific Lipid Transfer Protein (NsLTP) Genes in the Poaceae Family: Their Duplication and Diversity. Mol. Genet. Genom. 2008, 279, 481–497. [Google Scholar] [CrossRef]
- Noonan, J.; Williams, W.P.; Shan, X. Investigation of Antimicrobial Peptide Genes Associated with Fungus and Insect Resistance in Maize. Int. J. Mol. Sci. 2017, 18, 1938. [Google Scholar] [CrossRef] [Green Version]
- Jülke, S.; Ludwig-Müller, J. Response of Arabidopsis thaliana Roots with Altered Lipid Transfer Protein (LTP) Gene Expression to the Clubroot Disease and Salt Stress. Plants 2016, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, G.; Hao, X.; Cao, B.; Chen, Q.; Liu, S.; Lei, J. CaMF2, an Anther-Specific Lipid Transfer Protein (LTP) Gene, Affects Pollen Development in Capsicum annuum L. Plant Sci. 2011, 181, 439–448. [Google Scholar] [CrossRef]
- Jung, H.W.; Kim, W.; Hwang, B.K. Three Pathogen-Inducible Genes Encoding Lipid Transfer Protein from Pepper Are Differentially Activated by Pathogens, Abiotic, and Environmental Stresses*. Plant Cell Environ. 2003, 26, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Moraes, G.P.; Benitez, L.C.; do Amaral, M.N.; Vighi, I.L.; Auler, P.A.; da Maia, L.C.; Bianchi, V.J.; Braga, E.J.B. Expression of LTP Genes in Response to Saline Stress in Rice Seedlings. Genet. Mol. Res. 2015, 14, 8294–8305. [Google Scholar] [CrossRef]
- Carvalho, A.O.; Souza-Filho, G.A.; Ferreira, B.S.; Branco, A.T.; Araújo, I.S.; Fernandes, K.V.S.; Retamal, C.A.; Gomes, V.M. Cloning and Characterization of a Cowpea Seed Lipid Transfer Protein CDNA: Expression Analysis during Seed Development and under Fungal and Cold Stresses in Seedlings’ Tissues. Plant Physiol. Biochem. 2006, 44, 732–742. [Google Scholar] [CrossRef]
- Cammue, B.P.A.; Thevissen, K.; Hendriks, M.; Eggermont, K.; Goderis, I.J.; Proost, P.; Van Damme, J.; Osborn, R.W.; Guerbette, F.; Kader, J.C.; et al. A Potent Antimicrobial Protein from Onion Seeds Showing Sequence Homology to Plant Lipid Transfer Proteins. Plant Physiol. 1995, 109, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnikova, D.N.; Mineev, K.S.; Finkina, E.I.; Arseniev, A.S.; Ovchinnikova, T.V. A Novel Lipid Transfer Protein from the Dill Anethum graveolens L.: Isolation, Structure, Heterologous Expression, and Functional Characteristics. J. Pept. Sci. 2016, 22, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, A.K.; Brunstedt, J.; Nielsen, K.K.; Roepstorff, P.; Mikkelsen, J.D. Characterization of a new antifungal non-specific lipid transfer protein (nsLTP) from sugar beet leaves. Plant Sci. 2000, 155, 31–40. [Google Scholar] [CrossRef]
- Lin, P.; Xia, L.; Wong, J.H.; Ng, T.B.; Ye, X.; Wang, S.; Xiangzhu, S. Lipid Transfer Proteins from Brassica campestris and Mung Bean Surpass Mung Bean Chitinase in Exploitability. J. Pept. Sci. 2007, 13, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.J.; Sathoff, A.E.; Holl, C.; Bauer, B.; Samac, D.A.; Carter, C.J. The Major Nectar Protein of Brassica rapa Is a Non-Specific Lipid Transfer Protein, BrLTP2.1, with Strong Antifungal Activity. J. Exp. Bot. 2018, 69, 5587–5597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawrot, R.; Józefiak, D.; Sip, A.; Kuźma, D.; Musidlak, O.; Goździcka-Józefiak, A. Isolation and Characterization of a Non-Specific Lipid Transfer Protein from Chelidonium majus L. Latex. Int. J. Biol. Macromol. 2017, 104, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Zottich, U.; Da Cunha, M.; Carvalho, A.O.; Dias, G.B.; Silva, N.C.M.; Santos, I.S.; do Nacimento, V.V.; Miguel, E.C.; Machado, O.L.T.; Gomes, V.M. Purification, Biochemical Characterization and Antifungal Activity of a New Lipid Transfer Protein (LTP) from Coffea canephora Seeds with α-Amylase Inhibitor Properties. Biochim. Biophys. Acta BBA-Gen. Subj. 2011, 1810, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Bard, G.C.V.; Nascimento, V.V.; Ribeiro, S.F.F.; Rodrigues, R.; Perales, J.; Teixeira-Ferreira, A.; Carvalho, A.O.; Fernandes, K.V.S.; Gomes, V.M. Characterization of Peptides from Capsicum annuum Hybrid Seeds with Inhibitory Activity Against α-Amylase, Serine Proteinases and Fungi. Protein J. 2015, 34, 122–129. [Google Scholar] [CrossRef]
- Zaman, U.; Abbasi, A. Isolation, Purification and Characterization of a Nonspecific Lipid Transfer Protein from Cuminum cyminum. Phytochemistry 2009, 70, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Regente, M.C.; Giudici, A.M.; Villalaín, J.; De la Canal, L. The cytotoxic properties of a plant lipid transfer protein involve membrane permeabilization of target cells. Lett. Appl. Microbiol. 2005, 40, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.C.O.; Costa, A.S.; Lima, A.D.R.; Silva, F.D.A.; Lobo, M.D.P.; Monteiro-Moreira, A.C.O.; Moreira, R.A.; Leal, L.K.A.M.; Miron, D.; Vasconcelos, I.M.; et al. First Isolation and Antinociceptive Activity of a Lipid Transfer Protein from Noni (Morinda citrifolia) Seeds. Int. J. Biol. Macromol. 2016, 86, 71–79. [Google Scholar] [CrossRef]
- Campos, D.C.O.; Costa, A.S.; Luz, P.B.; Soares, P.M.G.; Alencar, N.M.N.; Oliveira, H.D. Morinda citrifolia Lipid Transfer Protein 1 Exhibits Anti-Inflammatory Activity by Modulation of pro- and Anti-Inflammatory Cytokines. Int. J. Biol. Macromol. 2017, 103, 1121–1129. [Google Scholar] [CrossRef]
- Wang, S.Y.; Wu, J.H.; Ng, T.B.; Ye, X.Y.; Rao, P.F. A Non-Specific Lipid Transfer Protein with Antifungal and Antibacterial Activities from the Mung Bean. Peptides 2004, 25, 1235–1242. [Google Scholar] [CrossRef]
- Bogdanov, I.V.; Shenkarev, Z.O.; Finkina, E.I.; Melnikova, D.N.; Rumynskiy, E.I.; Arseniev, A.S.; Ovchinnikova, T.V. A Novel Lipid Transfer Protein from the Pea Pisum sativum: Isolation, Recombinant Expression, Solution Structure, Antifungal Activity, Lipid Binding, and Allergenic Properties. BMC Plant Biol. 2016, 16, 107. [Google Scholar] [CrossRef] [Green Version]
- Terras, F.R.G.; Goderis, I.J.; Van Leuven, F.; Vanderleyden, J.; Cammue, B.P.A.; Broekaert, W.F. In Vitro Antifungal Activity of a Radish (Raphanus sativus L.) Seed Protein Homologous to Nonspecific Lipid Transfer Proteins 1. Plant Physiol. 1992, 100, 1055–1058. [Google Scholar] [CrossRef] [Green Version]
- Nazeer, M.; Waheed, H.; Saeed, M.; Ali, S.Y.; Choudhary, M.I.; Ul-Haq, Z.; Ahmed, A. Purification and Characterization of a Nonspecific Lipid Transfer Protein 1 (NsLTP1) from Ajwain (Trachyspermum ammi) Seeds. Sci. Rep. 2019, 9, 4148. [Google Scholar] [CrossRef]
- Gomar, J.; Petit, M.-C.; Sodano, P.; Sy, D.; Marion, D.; Kader, J.-C.; Vovelle, F.; Ptak, M. Solution structure and lipid binding of a nonspecific lipid transfer protein extracted from maize seeds. Protein Sci. 1996, 5, 565–577. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.; Betzel, C.; Pusey, M. Latest Methods of Fluorescence-Based Protein Crystal Identification. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2015, 71, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos-Silva, C.A.; Zupin, L.; Oliveira-Lima, M.; Vilela, L.M.B.; Bezerra-Neto, J.P.; Ferreira-Neto, J.R.; Ferreira, J.D.C.; de Oliveira-Silva, R.L.; de Pires, C.J.; Aburjaile, F.F.; et al. Plant Antimicrobial Peptides: State of the Art, In Silico Prediction and Perspectives in the Omics Era. Bioinform. Biol. Insights 2020, 14. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Krogh, A.; Brown, M.; Mian, I.S.; Sjölander, K.; Haussler, D. Hidden Markov Models in Computational Biology: Applications to Protein Modeling. J. Mol. Biol. 1994, 235, 1501–1531. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The Conserved Domain Database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [Green Version]
- Almagro, A.; Juan, J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Ferrè, F.; Clote, P. DiANNA 1.1: An Extension of the DiANNA Web Server for Ternary Cysteine Classification. Nucleic Acids Res. 2006, 34 (Suppl. 2), W182–W185. [Google Scholar] [CrossRef] [Green Version]
- Kozlowski, L.P. PC 2.0: Prediction of isoelectric point and pKa dissociation constants.Vnucleic. Acids Res. 2021, 49, W285–W292. [Google Scholar] [CrossRef] [PubMed]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMPR3: A Database on Sequences, Structures and Signatures of Antimicrobial Peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-T.; Lee, C.-C.; Yang, J.-R.; Lai, J.Z.C.; Chang, K.Y. A Large-Scale Structural Classification of Antimicrobial Peptides. BioMed Res. Int. 2015, 2015, e475062. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, H.; Lu, H.; Li, G.; Huang, Q. LAMP: A Database Linking Antimicrobial Peptides. PLoS ONE 2013, 8, e66557. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Klein, Z.; Cuevas-Zuviria, B.; Wangorsch, A.; Hernandez-Ramirez, G.; Pazos-Castro, D.; Oeo-Santos, C.; Romero-Sahagun, A.; Pacios, L.F.; Tome-Amat, J.; Diaz-Perales, A.; et al. The key to the allergenicity of lipid transfer protein (LTP) ligands: A structural characterization. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2021, 18866, 158928. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Studer, G.; Tauriello, G.; Bienert, S.; Biasini, M.; Johner, N.; Schwede, T. ProMod3—A versatile homology modelling toolbox. PLoS Comput. Biol. 2021, 17, e1008667. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative protein structure modeling using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robustelli, P.; Piana, S.; Shaw, D.E. Developing a molecular dynamics force field for both folded and disordered protein states. Proc. Natl. Acad. Sci. USA 2018, 115, E4758–E4766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, L.H.; Čopič, A.; Levine, T.P. Advances on the Transfer of Lipids by Lipid Transfer Proteins. Trends Biochem. Sci. 2017, 42, 516–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeo-Santos, C.; Mas, S.; Benedé, S.; López-Lucendo, M.; Quiralte, J.; Blanca, M.; Mayorga, C.; Villalba, M.; Barderas, R. A Recombinant Isoform of the Ole e 7 Olive Pollen Allergen Assembled by de Novo Mass Spectrometry Retains the Allergenic Ability of the Natural Allergen. J. Proteom. 2018, 187, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Buhler, S.; Akkerdaas, J.H.; Pertinhez, T.H.; Van Ree, R.; Dossena, A.; Sforza, S.; Tedeschi, T. Development of a Strategy for the Total Chemical Synthesis of an Allergenic Protein: The Peach LTP Pru p 3: CHEMICAL SYNTHESIS OF THE ALLERGENIC PEACH LIPID TRANSFER PROTEIN. J. Pept. Sci. 2017, 23, 282–293. [Google Scholar] [CrossRef]
- Deng, T.; Ge, H.; He, H.; Liu, Y.; Zhai, C.; Feng, L.; Yi, L. The Heterologous Expression Strategies of Antimicrobial Peptides in Microbial Systems. Protein Expr. Purif. 2017, 140, 52–59. [Google Scholar] [CrossRef]
- Parachin, N.S.; Mulder, K.C.; Viana, A.A.B.; Dias, S.C.; Franco, O.L. Expression Systems for Heterologous Production of Antimicrobial Peptides. Peptides 2012, 38, 446–456. [Google Scholar] [CrossRef]
- Safi, H.; Belgaroui, N.; Masmoudi, K.; Brini, F.; Safi, H.; Belgaroui, N.; Masmoudi, K.; Brini, F. Promoter of the Wheat Lipid Transfer Protein, TdLTP4, Drives Leaf-Preferential Expression in Transgenic Arabidopsis Plants. Funct. Plant Biol. 2019, 46, 275–285. [Google Scholar] [CrossRef]
- Canevascini, S.; Caderas, D.; Mandel, T.; Fleming, A.J.; Dupuis, I.; Kuhlemeier, C. Tissue-Specific Expression and Promoter Analysis of the Tobacco Ltp1 Gene. Plant Physiol. 1996, 112, 513–524. [Google Scholar] [CrossRef] [Green Version]
- Samuel, D.; Liu, Y.J.; Cheng, C.S.; Lyu, P.C. Solution Structure of Plant Nonspecific Lipid Transfer Protein-2 from Rice (Oryza Sativa). J. Biol. Chem. 2002, 277, 35267–35273. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, J.E.; Bin-Umer, M.A.; Widiez, T.; Finn, D.; McCormick, S.; Tumer, N.E. A Lipid Transfer Protein Increases the Glutathione Content and Enhances Arabidopsis Resistance to a Trichothecene Mycotoxin. PLoS ONE 2015, 10, e0130204. [Google Scholar] [CrossRef]
- Eguia, F.A.P.; Ramos, H.R.; Kraschowetz, S.; Omote, D.; Ramos, C.R.R.; Ho, P.L.; Carvalho, E.; Gonçalves, V.M. A new vector for heterologous gene expression in Escherichia coli with increased stability in the absence of antibiotic. Plasmid 2018, 98, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Wang, J.; Wang, M.; Zhang, H.; Wang, S.; Zhao, Y. Molecular Cloning, Recombinant Expression, and Antifungal Functional Characterization of the Lipid Transfer Protein from Panax ginseng. Biotechnol. Lett. 2016, 38, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Harper, S.; Speicher, D.W. Purification of Proteins Fused to Glutathione S-Transferase. In Protein Chromatography: Methods and Protocols; Walls, D., Loughran, S.T., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; pp. 259–280. ISBN 978-1-60761-913-0. [Google Scholar]
- Duan, G.; Ding, L.; Wei, D.; Zhou, H.; Chu, J.; Zhang, S.; Qian, J. Screening Endogenous Signal Peptides and Protein Folding Factors to Promote the Secretory Expression of Heterologous Proteins in Pichia pastoris. J. Biotechnol. 2019, 306, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Pokoj, S.; Lauer, I.; Fötisch, K.; Himly, M.; Mari, A.; Enrique, E.; Miguel-Moncin, M.D.M.; Lidholm, J.; Vieths, S.; Scheurer, S. Pichia pastoris Is Superior to E. Coli for the Production of Recombinant Allergenic Non-Specific Lipid-Transfer Proteins. Protein Expr. Purif. 2010, 69, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, T.Q.; Santos, S.B.; Martins, I.M.; Domingues, L.; Oliveira, C. Chapter 9-Production and Bioengineering of Recombinant Pharmaceuticals. In Proteins: Sustainable Source, Processing and Applications; Galanakis, C.M., Ed.; Academic Press: Cambridge, MA, USA, 2019; pp. 259–293. ISBN 978-0-12-816695-6. [Google Scholar]
- Rihs, H.-P.; Ruëff, F.; Lundberg, M.; Rozynek, P.; Barber, D.; Scheurer, S.; Cisteró-Bahima, A.; Brüning, T.; Raulf-Heimsoth, M. Relevance of the Recombinant Lipid Transfer Protein of Hevea brasiliensis: IgE-Binding Reactivity in Fruit-Allergic Adults. Ann. Allergy. Asthma. Immunol. 2006, 97, 643–649. [Google Scholar] [CrossRef]
- Bogdanov, I.V.; Finkina, E.I.; Baladin, S.V.; Melnikova, D.N.; Stukacheva, E.A.; Ovchinnikova, T.V. Structural and Functional Characterization of Recombinant Isoforms of the Lentil Lipid Transfer Protein. Acta Nat. 2015, 7, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Mhaske, V.A.; Datla, R.; Qiu, S.; Harsulkar, A.M. Isolation and Characterization of Genes Encoding Lipid Transfer Proteins in Linum usitatissimum. Biol. Plant. 2016, 60, 285–291. [Google Scholar] [CrossRef]
- Gräslund, S.; Nordlund, P.; Weigelt, J.; Hallberg, B.M.; Bray, J.; Gileadi, O.; Knapp, S.; Oppermann, U.; Arrowsmith, C.; Hui, R.; et al. Protein Production and Purification. Nat. Methods 2008, 5, 135–146. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant Protein Expression in Escherichia coli: Advances and Challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [Green Version]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial Resistance and the Alternative Resources with Special Emphasis on Plant-Based Antimicrobials—A Review. Plants 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Prabhakaran, V.-S.; Kim, K. The Multi-Faceted Potential of Plant-Derived Metabolites as Antimicrobial Agents against Multidrug-Resistant Pathogens. Microb. Pathog. 2018, 116, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Ferdes, M. Antimicrobial Compounds from Plants; IAPC-OBP: Zagreb, Croatia, 2018; pp. 243–271. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K.C. The Biochemistry and Biology of Extracellular Plant Lipid-Transfer Proteins (LTPs). Protein Sci. 2008, 17, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Stotz, H.U.; Waller, F.; Wang, K. Innate Immunity in Plants: The Role of Antimicrobial Peptides. In Antimicrobial Peptides and Innate Immunity; Hiemstra, P.S., Zaat, S.A.J., Eds.; Progress in Inflammation Research; Springer: Basel, Switzerland, 2013; pp. 29–51. ISBN 978-3-0348-0541-4. [Google Scholar]
- Finkina, E.I.; Melnikova, D.N.; Bogdanov, I.V.; Ovchinnikova, T.V. Plant Pathogenesis-Related Proteins PR-10 and PR-14 as Components of Innate Immunity System and Ubiquitous Allergens. Curr. Med. Chem. 2017, 24, 1772–1787. [Google Scholar] [CrossRef] [PubMed]
- Segura, A.; Moreno, M.; García-Olmedo, F. Purification and Antipathogenic Activity of Lipid Transfer Proteins (LTPs) from the Leaves of Arabidopsis and Spinach. FEBS Lett. 1993, 332, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Regente, M.C.; Canal, L.D.L. Purification, Characterization and Antifungal Properties of a Lipid-Transfer Protein from Sunflower (Helianthus annuus) Seeds. Physiol. Plant. 2000, 110, 158–163. [Google Scholar] [CrossRef]
- Jia, Z.; Gou, J.; Sun, Y.; Yuan, L.; Tang, Q.; Yang, X.; Pei, Y.; Luo, K. Enhanced Resistance to Fungal Pathogens in Transgenic Populus Tomentosa Carr. by Overexpression of an NsLTP-like Antimicrobial Protein Gene from Motherwort (Leonurus japonicus). Tree Physiol. 2010, 30, 1599–1605. [Google Scholar] [CrossRef] [Green Version]
- Bechinger, B. Structure and Functions of Channel-Forming Peptides: Magainins, Cecropins, Melittin and Alamethicin. J. Membr. Biol. 1997, 156, 197–211. [Google Scholar] [CrossRef]
- Jayaraj, J.; Punja, Z.K. Combined Expression of Chitinase and Lipid Transfer Protein Genes in Transgenic Carrot Plants Enhances Resistance to Foliar Fungal Pathogens. Plant Cell Rep. 2007, 26, 1539–1546. [Google Scholar] [CrossRef]
- Zou, H.-W.; Tian, X.-H.; Ma, G.-H.; Li, Z.-X. Isolation and Functional Analysis of ZmLTP3, a Homologue to Arabidopsis LTP3. Int. J. Mol. Sci. 2013, 14, 5025–5035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luepke, K.H.; Suda, K.J.; Boucher, H.; Russo, R.L.; Bonney, M.W.; Hunt, T.D.; Mohr, J.F. Past, Present, and Future of Antibacterial Economics: Increasing Bacterial Resistance, Limited Antibiotic Pipeline, and Societal Implications. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2017, 37, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and Consequences of Bacterial Resistance to Antimicrobial Peptides. Drug Resist. Updat. 2016, 26, 43–57. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Carvalho, A.; Gomes, V.M. Role of Plant Lipid Transfer Proteins in Plant Cell Physiology—A Concise Review. Peptides 2007, 28, 1144–1153. [Google Scholar] [CrossRef]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant Antimicrobial Peptides. Folia Microbiol. 2014, 59, 181–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaci, Z.; Shalaby, L.; Kamal, S.; Boudjerra, N.; Gao, X.; Xue, M.; Belkacem, S.A.; Ibrahim, A.G.A.; Kotb, R.; Charbonneau, C. Treatment of Immunocompromised Patients with Suspected Invasive Fungal Infections: Economic Analysis of Diagnostic-Driven versus Empirical Strategies in Algeria and Egypt. Expert Rev. Pharmacoecon. Outcomes Res. 2019, 19, 693–700. [Google Scholar] [CrossRef]
- Cheng, C.-S.; Chen, M.-N.; Liu, Y.-J.; Huang, L.-Y.; Lin, K.-F.; Lyu, P.-C. Evaluation of Plant Non-Specific Lipid-Transfer Proteins for Potential Application in Drug Delivery. Enzym. Microb. Technol. 2004, 35, 532–539. [Google Scholar] [CrossRef]
- Diz, M.S.; Carvalho, A.O.; Ribeiro, S.F.F.; Cunha, M.D.; Beltramini, L.; Rodrigues, R.; Nascimento, V.V.; Machado, O.L.T.; Gomes, V.M. Characterisation, Immunolocalisation and Antifungal Activity of a Lipid Transfer Protein from Chili Pepper (Capsicum annuum) Seeds with Novel α-Amylase Inhibitory Properties. Physiol. Plant. 2011, 142, 233–246. [Google Scholar] [CrossRef]
- Cruz, L.P.; Ribeiro, S.F.F.; Carvalho, A.O.; Vasconcelos, I.M.; Rodrigues, R.; Da Cunha, M.; Gomes, V.M. Isolation and Partial Characterization of a Novel Lipid Transfer Protein (LTP) and Antifungal Activity of Peptides from Chilli Pepper Seeds. Protein Pept. Lett. 2010, 17, 311–318. [Google Scholar] [CrossRef]
- Ooi, L.S.M.; Tian, L.; Su, M.; Ho, W.-S.; Sun, S.S.M.; Chung, H.-Y.; Wong, H.N.C.; Ooi, V.E.C. Isolation, Characterization, Molecular Cloning and Modeling of a New Lipid Transfer Protein with Antiviral and Antiproliferative Activities from Narcissus tazetta. Peptides 2008, 29, 2101–2109. [Google Scholar] [CrossRef]
- Peretti, D.; Kim, S.; Tufi, R.; Lev, S. Lipid Transfer Proteins and Membrane Contact Sites in Human Cancer. Front. Cell Dev. Biol. 2020, 7, 371. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, L.D.A.; Taveira, G.B.; Ribeiro, S.D.F.F.; da Silva Pereira, L.; de Oliveira Carvalho, A.; Rodrigues, R.; Oliveira, A.E.A.; Machado, O.L.T.; da Silva Araújo, J.; Vasconcelos, I.M.; et al. Purification and Characterization of Peptides from Capsicum annuum Fruits Which Are α-Amylase Inhibitors and Exhibit High Antimicrobial Activity against Fungi of Agronomic Importance. Protein Expr. Purif. 2017, 132, 97–107. [Google Scholar] [CrossRef]
- da Silva, F.C.V.; Pessoa Costa, E.; Moreira Gomes, V.; de Oliveira Carvalho, A. Inhibition Mechanism of Human Salivary α-Amylase by Lipid Transfer Protein from Vigna unguiculata. Comput. Biol. Chem. 2020, 85, 107193. [Google Scholar] [CrossRef]
- da Silva, F.C.V.; do Nascimento, V.V.; Machado, O.L.T.; da Silva Pereira, L.; Gomes, V.M.; de Oliveira Carvalho, A. Insight into the α-Amylase Inhibitory Activity of Plant Lipid Transfer Proteins. J. Chem. Inf. Model. 2018, 58, 2294–2304. [Google Scholar] [CrossRef] [PubMed]
- Pato, C.; Le Borgne, M.; Le Baut, G.; Le Pape, P.; Marion, D.; Douliez, J.-P. Potential Application of Plant Lipid Transfer Proteins for Drug Delivery. Biochem. Pharmacol. 2001, 62, 555–560. [Google Scholar] [CrossRef]
- Li, N.; Xu, C.; Li-Beisson, Y.; Philippar, K. Fatty Acid and Lipid Transport in Plant Cells. Trends Plant Sci. 2016, 21, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oeo-Santos, C.; López-Rodríguez, J.C.; García-Mouton, C.; Segundo-Acosta, P.S.; Jurado, A.; Moreno-Aguilar, C.; García-Álvarez, B.; Pérez-Gil, J.; Villalba, M.; Barderas, R.; et al. Biophysical and Biological Impact on the Structure and IgE-Binding of the Interaction of the Olive Pollen Allergen Ole e 7 with Lipids. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183258. [Google Scholar] [CrossRef] [PubMed]
- Maghsoudi, S.; Ashrafi-Kooshk, M.R.; Shahlaei, M.; Ghadami, S.A.; Ghobadi, S.; Mostafaie, A.; Khodarahmi, R. Comparative Evaluation of Amphotericin B Binding to the Native and Modified Forms of Rice Lipid-Transfer Protein: A Possible Perspective on Improving the Drug-Binding Affinity and Specificity. J. Iran. Chem. Soc. 2013, 10, 937–950. [Google Scholar] [CrossRef]
- Aruanno, A.; Urbani, S.; Frati, F.; Nucera, E. LTP Allergy/Sensitization in a Pediatric Population. Allergol. Immunopathol. 2020, 48, 763–770. [Google Scholar] [CrossRef]
- Bartra, J.; Sastre, J.; del Cuvillo, A.; Montoro, J.; Jáuregui, I.; Dávila, I.; Ferrer, M.; Mullol, J.; Valero, A. From Pollinosis to Digestive Allergy. J. Investig. Allergol. Clin. Immunol. 2009, 19 (Suppl. 1), 3–10. [Google Scholar] [PubMed]
- Scheurer, S.; Schülke, S. Interaction of Non-Specific Lipid-Transfer Proteins With Plant-Derived Lipids and Its Impact on Allergic Sensitization. Front. Immunol. 2018, 9, 1389. [Google Scholar] [CrossRef] [Green Version]
- Kronfel, C.M.; Cheng, H.; Hurlburt, B.K.; Simon, R.J.; Maleki, S.J. Epitope Mapping for the Non-Specific Lipid Transfer Proteins (nsLTP) Among Peanut Allergic Patients. J. Allergy Clin. Immunol. 2018, 141, AB239. [Google Scholar] [CrossRef]
- Matsuo, H.; Yokooji, T.; Taogoshi, T. Common Food Allergens and Their IgE-Binding Epitopes. Allergol. Int. 2015, 64, 332–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucera, E.; Rienzo, A.D.; Ricci, A.G.; Buonomo, A.; Mezzacappa, S.; Schiavino, D. Adverse Reactions to Nonsteroidal Anti-Inflammatory Drugs and Hypersensitivity to Lipid Transfer Proteins. Adv. Dermatol. Allergol. Dermatol. Alergol. 2016, 33, 309–310. [Google Scholar] [CrossRef] [Green Version]
- Pastorello, E.A.; Pravettoni, V.; Trambaioli, C.; Pompei, C.; Brenna, O.; Farioli, L.; Conti, A. Lipid Transfer Proteins and 2S Albumins as Allergens. Allergy 2001, 56, 45–47. [Google Scholar] [CrossRef]
- Martín-Pedraza, L.; Wangorsch, A.; Bueno-Diaz, C.; de las Heras, M.; Scheurer, S.; Cuesta-Herranz, J.; Villalba, M. 2S Albumins and NsLTP Are Involved in Anaphylaxis to Pizza Sauce: IgE Recognition before and after Allergen Processing. Food Chem. 2020, 321, 126679. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Pompei, C.; Pravettoni, V.; Farioli, L.; Calamari, A.M.; Scibilia, J.; Robino, A.M.; Conti, A.; Iametti, S.; Fortunato, D.; et al. Lipid-Transfer Protein Is the Major Maize Allergen Maintaining IgE-Binding Activity after Cooking at 100 °C, as Demonstrated in Anaphylactic Patients and Patients with Positive Double-Blind, Placebo-Controlled Food Challenge Results. J. Allergy Clin. Immunol. 2003, 112, 775–783. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Pravettoni, V.; Farioli, L.; Ispano, M.; Fortunato, D.; Monza, M.; Giuffrida, M.G.; Rivolta, F.; Scibola, E.; Ansaloni, R.; et al. Clinical Role of a Lipid Transfer Protein That Acts as a New Apple-Specific Allergen. J. Allergy Clin. Immunol. 1999, 104, 1099–1106. [Google Scholar] [CrossRef]
- Gülsen, A.; Jappe, U. Lipid Transfer Protein Sensitization in an Apple-Allergic Patient: A Case Report from Northern Europe. Eur. Ann. Allergy Clin. Immunol. 2018, 51, 80. [Google Scholar] [CrossRef] [Green Version]
- Tabar, A.I.; Alvarez-Puebla, M.J.; Gomez, B.; Sanchez-Monge, R.; García, B.E.; Echechipia, S.; Olaguibel, J.M.; Salcedo, G. Diversity of Asparagus Allergy: Clinical and Immunological Features. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2004, 34, 131–136. [Google Scholar] [CrossRef]
- Palacín, A.; Cumplido, J.; Figueroa, J.; Ahrazem, O.; Sánchez-Monge, R.; Carrillo, T.; Salcedo, G.; Blanco, C. Cabbage Lipid Transfer Protein Bra o 3 Is a Major Allergen Responsible for Cross-Reactivity between Plant Foods and Pollens. J. Allergy Clin. Immunol. 2006, 117, 1423–1429. [Google Scholar] [CrossRef]
- Sánchez-Monge, R.; Blanco, C.; López-Torrejón, G.; Cumplido, J.; Recas, M.; Figueroa, J.; Carrillo, T.; Salcedo, G. Differential Allergen Sensitization Patterns in Chestnut Allergy with or without Associated Latex-Fruit Syndrome. J. Allergy Clin. Immunol. 2006, 118, 705–710. [Google Scholar] [CrossRef]
- Zoccatelli, G.; Pokoj, S.; Foetisch, K.; Bartra, J.; Valero, A.; del Mar San Miguel-Moncin, M.; Vieths, S.; Scheurer, S. Identification and Characterization of the Major Allergen of Green Bean (Phaseolus vulgaris) as a Non-Specific Lipid Transfer Protein (Pha v 3). Mol. Immunol. 2010, 47, 1561–1568. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Pravettoni, V.; Farioli, L.; Rivolta, F.; Conti, A.; Ispano, M.; Fortunato, D.; Bengtsson, A.; Bianchi, M. Hypersensitivity to Mugwort (Artemisia vulgaris) in Patients with Peach Allergy Is Due to a Common Lipid Transfer Protein Allergen and Is Often without Clinical Expression. J. Allergy Clin. Immunol. 2002, 110, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Le, T.-M.; Bublin, M.; Breiteneder, H.; Fernández-Rivas, M.; Asero, R.; Ballmer-Weber, B.; Barreales, L.; Bures, P.; Belohlavkova, S.; de Blay, F.; et al. Kiwifruit Allergy across Europe: Clinical Manifestation and IgE Recognition Patterns to Kiwifruit Allergens. J. Allergy Clin. Immunol. 2013, 131, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Akkerdaas, J.; Finkina, E.I.; Balandin, S.V.; Magadán, S.S.; Knulst, A.; Fernandez-Rivas, M.; Asero, R.; van Ree, R.; Ovchinnikova, T.V. Lentil (Lens culinaris) Lipid Transfer Protein Len c 3: A Novel Legume Allergen. Int. Arch. Allergy Immunol. 2012, 157, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Moncín, M.S.; Krail, M.; Scheurer, S.; Enrique, E.; Alonso, R.; Conti, A.; Cisteró-Bahíma, A.; Vieths, S. Lettuce Anaphylaxis: Identification of a Lipid Transfer Protein as the Major Allergen. Allergy 2003, 58, 511–517. [Google Scholar] [CrossRef]
- Ebo, D.G.; Ahrazem, O.; Lopez-Torrejon, G.; Bridts, C.H.; Salcedo, G.; Stevens, W.J. Anaphylaxis from Mandarin (Citrus reticulata): Identification of Potential Responsible Allergens. Int. Arch. Allergy Immunol. 2007, 144, 39–43. [Google Scholar] [CrossRef]
- Vereda, A.; Sirvent, S.; Villalba, M.; Rodríguez, R.; Cuesta-Herranz, J.; Palomares, O. Improvement of Mustard (Sinapis alba) Allergy Diagnosis and Management by Linking Clinical Features and Component-Resolved Approaches. J. Allergy Clin. Immunol. 2011, 127, 1304–1307. [Google Scholar] [CrossRef]
- Ahrazem, O.; Ibáñez, M.D.; López-Torrejón, G.; Sánchez-Monge, R.; Sastre, J.; Lombardero, M.; Barber, D.; Salcedo, G. Lipid Transfer Proteins and Allergy to Oranges. Int. Arch. Allergy Immunol. 2005, 137, 201–210. [Google Scholar] [CrossRef]
- Krause, S.; Reese, G.; Randow, S.; Zennaro, D.; Quaratino, D.; Palazzo, P.; Ciardiello, M.A.; Petersen, A.; Becker, W.-M.; Mari, A. Lipid Transfer Protein (Ara h 9) as a New Peanut Allergen Relevant for a Mediterranean Allergic Population. J. Allergy Clin. Immunol. 2009, 124, 771–778.e5. [Google Scholar] [CrossRef] [PubMed]
- Vereda, A.; van Hage, M.; Ahlstedt, S.; Ibañez, M.D.; Cuesta-Herranz, J.; van Odijk, J.; Wickman, M.; Sampson, H.A. Peanut Allergy: Clinical and Immunologic Differences among Patients from 3 Different Geographic Regions. J. Allergy Clin. Immunol. 2011, 127, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Enrique, E.; Utz, M.; de Mateo, J.A.; Castelló, J.V.; Malek, T.; Pineda, F. Allergy to Lipid Transfer Proteins: Cross-reactivity Among Pomegranate, Hazelnut, And Peanut. Ann. Allergy Asthma Immunol. 2006, 96, 122–123. [Google Scholar] [CrossRef]
- Le, L.Q.; Lorenz, Y.; Scheurer, S.; Fötisch, K.; Enrique, E.; Bartra, J.; Biemelt, S.; Vieths, S.; Sonnewald, U. Design of Tomato Fruits with Reduced Allergenicity by DsRNAi-Mediated Inhibition of Ns-LTP (Lyc e 3) Expression. Plant Biotechnol. J. 2006, 4, 231–242. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Farioli, L.; Pravettoni, V.; Robino, A.M.; Scibilia, J.; Fortunato, D.; Conti, A.; Borgonovo, L.; Bengtsson, A.; Ortolani, C. Lipid Transfer Protein and Vicilin Are Important Walnut Allergens in Patients Not Allergic to Pollen. J. Allergy Clin. Immunol. 2004, 114, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Palacin, A.; Quirce, S.; Armentia, A.; Fernández-Nieto, M.; Pacios, L.F.; Asensio, T.; Sastre, J.; Diaz-Perales, A.; Salcedo, G. Wheat Lipid Transfer Protein Is a Major Allergen Associated with Baker’s Asthma. J. Allergy Clin. Immunol. 2007, 120, 1132–1138. [Google Scholar] [CrossRef]
- Zuidmeer, L.; van Ree, R. Lipid Transfer Protein Allergy: Primary Food Allergy or Pollen/Food Syndrome in Some Cases. Curr. Opin. Allergy Clin. Immunol. 2007, 7, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Rougé, P.; Borges, J.-P.; Culerrier, R.; Brulé, C.; Didier, A.; Barre, A. Les allergies alimentaires aux fruits. Rev. Fr. Allergol. 2009, 49, S23–S27. [Google Scholar] [CrossRef]
- Salcedo, G.; Sánchez-Monge, R.; Barber, D.; Díaz-Perales, A. Plant Non-Specific Lipid Transfer Proteins: An Interface between Plant Defence and Human Allergy. Biochim. Biophys. Acta 2007, 1771, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Borges, J.-P.; Barre, A.; Culerrier, R.; Granier, C.; Didier, A.; Rougé, P. Lipid Transfer Proteins from Rosaceae Fruits Share Consensus Epitopes Responsible for Their IgE-Binding Cross-Reactivity. Biochem. Biophys. Res. Commun. 2008, 365, 685–690. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Taxa | Expressed under/Putative Function | Experimental Background | References |
|---|---|---|---|
| Arabidopsis thaliana | Plant development, biotic stress, abiotic stress | Microarray, RT-qPCR | [62,72] |
| Brassica spp. | Abiotic stress, biotic stress | EST, vector cloning, RNA-Seq | [55,69] |
| Capsicum annuum | Abiotic stress, biotic stress, molecular signaling | RNA gel blot, RT-qPCR | [73,74] |
| Glycine max | Abiotic stress, biotic stress | In silico SuperSAGE tags screening | [52] |
| Oryza sativa | Abiotic stress, plant development | RT-qPCR | [75] |
| Saccharum spp. | Abiotic stress, as signaling molecule | RT-qPCR | [60] |
| Solanum lycopersicum | Plant development | RNA-Seq/RT-qPCR | [54] |
| Triticum spp. | Biotic stress, abiotic stress | RT-qPCR | [25,29] |
| Vigna unguiculata | Abiotic stress, biotic stress, plant development | Northern blot, vector cloning | [66,76] |
| Zea mays | Biotic stress | RT-qPCR | [71] |
| Species | nsLTP Name | Source | Extraction | Precipitation | Chromatography (or Other) | Biological Activity | References |
|---|---|---|---|---|---|---|---|
| Allium cepa | Ace-AMP1 | Seed | Buffer solution | ammonium sulfate (85%) a | ion exchange and RP-HPLC b | antifungal and antibacterial | [77] |
| Anethum graveolens | Ag-LTP | Aerial parts | Buffer solution | - | ion exchange and RP-HPLC | antifungal * | [78] |
| Beta vulgaris | IWF5 | Leaf | Acid | - | ion exchange and RP-HPLC | antifungal | [79] |
| Brassica campestris | Brassica campestris nsLTP | Seed | Not mentioned | - | ion exchange, affinity and gel filtration | antifungal | [80] |
| Brassica rapa | BrLTP2.1 | Nectar | - | - | 1-D SDS-PAGE | antifungal * | [81] |
| Chelidonium majus | CmLTP 9.5 | Root latex | Buffer solution | isoelectric point | 1-D SDS-PAGE | antibacterial ** | [82] |
| Coffea canefora | Cc-LTP1 | Seed | Buffer solution | ammonium sulfate (90%) | ion exchange and RP-HPLC | α-amylase inhibitor and antifungal | [83] |
| Cc-LTP2 | Seed | Acid | - | ion exchange and RP-HPLC | antifungal and antibacterial | [84] | |
| Cuminum cyminum | Cumin nsLTP1 | Seed | Buffer solution | ammonium sulfate (72%) | gel filtration and RP-HPLC | - | [85] |
| Helianthus annus | HA-AP10 | Seed | Buffer solution | ammonium sulfate (80%) | gel filtration, ion exchange and RP-HPLC | antifungal | [86] |
| Morinda citrifolia | McLTP1 | Seed | Buffer solution | - | gel filtration and RP-HPLC | antibacterial, anti-inflammatory c, lethal sepsis prevention c and antinociceptive c | [24,87,88] |
| Phaseolus mungo | mung bean nsLTP | Seed | Buffer solution | ammonium sulfate (80%) | ion exchange | antifungal and antibacterial | [89] |
| Pisum sativum | Ps-LTP1 | Seed | Buffer solution | - | ion exchange and RP-HPLC | antifungal d, antibacterial d and allergen d | [90] |
| Raphanus sativus | radish ns-LTP-like | Seed | Not mentioned | ammonium sulfate (30–70%) | ion exchange and RP-HPLC | antifungal | [91] |
| Trachyspermum ammi | ajwain nsLTP1 | Seed | buffer solution | ammonium sulfate (80%) | gel filtration and RP-HPLC | - | [92] |
| Origin Species | Host System | Express nsLTP | Biological Activity | Yield (mg.L−1 of Host Culture) | Reference |
|---|---|---|---|---|---|
| Anethum graveolens | Escherichia coli | His8-TrxL-Ag-LTP | Antifungal | 1.5 | [78] |
| Brassica rapa | Escherichia coli | T7-BrLTP2.1-His6 | Antifungal | Not informed | [81] |
| Corylus avellana | Escherichia coli | His6-Cor a 8 | Allergen | 0.3 | [125] |
| Pichia pastoris | Cor a. 8 | Allergen | 82 | [125] | |
| Hevea brasiliensis | Escherichia coli | MBP-rHev b 12 | Allergen | 4 | [126] |
| Lens culinaris | Escherichia coli | His8-TrxL-Lc-LTP1 | Antifungal, antibacterial, allergen | 3 | [127] |
| Escherichia coli | His8-TrxL-Lc-LTP3 | Antifungal, antibacterial, allergen | 5 | [127] | |
| Prunus persica | Escherichia coli | His8-TrxL-Pru p 3 | Antifungal, antibacterial, allergen | 4 | [127] |
| Linum usitatissimum | Saccharomyces cerevisiae | (His tag) LuLTP_Ls1 e LuLTP_Ls4 | Antibacterial | Not informed | [128] |
| Panax ginseng | Escherichia coli | GST-pgLTP-His6 | Antifungal | Not informed | [121] |
| Pisum sativum | Escherichia coli | His8-TrxL-Ps-LTP1 | Antifungal, antibacterial, allergen | 5 | [90] |
| Solanum lycopersicum | Escherichia coli | (His tag) Sola l 3 | - | 0.5 | [53] |
| Food Source | Taxon | nsLTP Allergen | Reported Geographical Areas | Sources |
|---|---|---|---|---|
| Apple | Malus domestica (Rosaceae) | Mald 3 | Mainly southern Europe, also northern Europe and Australia | [169,170] |
| Asparagus | Aparagus officinalis (Asparagaceae) | Aspao 1 | Mainly southern Europe | [171] |
| Cabbage | Brassica oleracea (Brassicaceae) | Brao 3 | Mainly southern Europe | [172] |
| Chestnut | Castanea sativa (Fagaceae) | Cass 8 | Mainly southern Europe, also USA | [173] |
| Grape | Vitis vinifera (Vitaceae) | Vitv 1 | Mainly southern Europe, also Australia and Germany | [168] |
| Green bean | Phaseolus vulgaris (Leguminosae) | Phav 3 | Mainly southern Europe | [174] |
| Hazelnut | Corylus avellana (Betulaceae) | Cora 8 | Mainly southern Europe, also Switzerland and Denmark | [175] |
| Kiwi | Actinidia deliciosa (Actinidiaceae) | Actd10 | Mainly southern Europe | [176] |
| Lentil | Lens culinaris (Leguminosae) | Lenc 3 | Mainly southern Europe | [177] |
| Lettuce | Lactuca sativa (Asteraceae) | Lacs 1 | Mainly southern Europe | [178] |
| Mandarin | Citrus reticulata (Rutaceae) | Citr 3 | Mainly southern Europe | [179] |
| White Mustard | Sinapis alba (Brassicaceae) | Sina 3 | Mainly southern Europe | [180] |
| Oranges | Citrus spp. (Rutaceae) | Cits 3 | Mainly southern Europe | [181] |
| Peanut | Arachis hypogaea (Leguminosae) | Arah 9 | Mainly southern Europe, also USA | [182,183] |
| Pomegranate | Punica granatum (Punicaceae) | Pung 1 | Mainly southern Europe | [184] |
| Tomato | Solanum lycopersicon (Solanaceae) | Lyce 3 | Mainly southern Europe | [185] |
| Walnut | Juglans regia (Juglandaceae) | Jugr 3 | Mainly southern Europe, also USA | [186] |
| Wheat | Triticum aestivum (Poaceae) | Tria 14 | Mainly southern Europe | [187] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amador, V.C.; Santos-Silva, C.A.d.; Vilela, L.M.B.; Oliveira-Lima, M.; de Santana Rêgo, M.; Roldan-Filho, R.S.; Oliveira-Silva, R.L.d.; Lemos, A.B.; de Oliveira, W.D.; Ferreira-Neto, J.R.C.; et al. Lipid Transfer Proteins (LTPs)—Structure, Diversity and Roles beyond Antimicrobial Activity. Antibiotics 2021, 10, 1281. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111281
Amador VC, Santos-Silva CAd, Vilela LMB, Oliveira-Lima M, de Santana Rêgo M, Roldan-Filho RS, Oliveira-Silva RLd, Lemos AB, de Oliveira WD, Ferreira-Neto JRC, et al. Lipid Transfer Proteins (LTPs)—Structure, Diversity and Roles beyond Antimicrobial Activity. Antibiotics. 2021; 10(11):1281. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111281
Chicago/Turabian StyleAmador, Vinícius Costa, Carlos André dos Santos-Silva, Lívia Maria Batista Vilela, Marx Oliveira-Lima, Mireli de Santana Rêgo, Ricardo Salas Roldan-Filho, Roberta Lane de Oliveira-Silva, Ayug Bezerra Lemos, Wilson Dias de Oliveira, José Ribamar Costa Ferreira-Neto, and et al. 2021. "Lipid Transfer Proteins (LTPs)—Structure, Diversity and Roles beyond Antimicrobial Activity" Antibiotics 10, no. 11: 1281. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111281