1. Introduction
The predisposition of
S. aureus to cause infections requires the deployment of numerous virulence factors. Using these factors,
S. aureus colonizes, disseminates, and adapts to various environments in the host and causes infections. Once they establish the infection they subvert host functions, invade tissues, and overcome host defenses [
1]. Antibiotics are currently used to control the spread of
S. aureus infections. They subside bacterial infections either by killing, decelerating, or suspending their growth. During this process, antibiotics raise pressure for the selection of resistant or tolerant strains [
2]. One of the best examples is the emergence of resistance to methicillin in
S. aureus. The resistant strains were first reported within two years after being marketed [
3]. This robust emergence of resistance in pathogens like
S. aureus has made antibiotics ineffective [
4]. In addition to discovering novel antibiotics, finding antibiotic adjuvants is a promising approach to treating infections caused by resistant bacteria. Using these novel adjuvants, we can tackle the problems arising from safe and old antibiotics.
In addition to antibiotic resistance, sub-inhibitory concentrations of antibiotics may exacerbate infection by inducing staphylococcal pathogenicity [
5]. For example, in previous research, we found that sub-inhibitory doses of ampicillin increased the adhesion and invasion of
S. aureus on epithelial cells [
6]. Thus, the quest for an alternate and long-lasting antivirulence agent is essential and very important. Antivirulence agents operate differently from antibiotics and have distinct benefits. For instance, the development of resistant or tolerant strains may be challenging for this class of drugs, and their administration may have no effect on the typical flora of the host [
7]. In our previous study, we identified an antivirulence agent, M21, which specifically inhibited the activity of a key virulence determinant Clp protease. The inactivation of Clp protease by M21 further repressed multiple virulence factors simultaneously. M21 reduced the ratio of adherence and invasion in the host and attenuated the establishment of infection in mice by
S. aureus [
8]. Thus, different from traditional antibiotic adjuvants, antivirulence agents may compensate for the shortcomings of antibiotics and suppress
S. aureus pathogenicity effectively.
In this work, we examined the impact of antivirulence agent M21 on staphylococcal virulence factors induced by sub-inhibitory concentrations of antibiotics. Using peritonitis and bacteremia infection models, we found that the antibiotic adjuvant M21 substantially curtailed the β-lactam-induced exacerbation of S. aureus infections. Thus, our findings proved that the novel antibiotic adjuvant M21 can reduce virulence induced by sub-inhibitory concentrations of antibiotics in vivo and illustrated the possibility of using a combination therapy comprising antivirulence agents and antibiotics to treat staphylococcal infections.
3. Discussion
The objective of conventional drug development strategies is to discover the next chemical or molecular entity with a novel mode of action. For each promising novel substance, the journey from early discovery to market introduction is slow, expensive, and rife with obstacles. Moving a novel medicine from pre-clinical stages to the market typically involves a minimum of 10 to 12 years and costs over USD 2 billion [
10]. Antibiotics, which are considered one of the greatest discoveries of humankind, received immense scientific and pharmaceutical attention in the 20th century. However, the emergence of antibiotic-resistant strains created irreparable damage not only to the pharmaceutical industry but also to the people suffering from life-threatening bacterial infections. To combat these drawbacks, the zeal for the discovery of novel antibiotics or adjuvants, or alternatives to antibiotics has not stopped. Especially, there has been tremendous progress in the discovery of novel adjuvants with the hope that they can “Make Antibiotics Great Again” [
11]. However, studies have shown that the sub-inhibitory concentration of antibiotics such as ampicillin can induce virulence expression in
S. aureus. Despite this drawback, there has been limited progress in discovering novel molecules that can control antibiotic-induced virulence. Thus, to address this lacuna, in the current study we focused on one such novel molecule called M21, which not only displayed antivirulence properties against MRSA but also functioned as an adjuvant for β-lactam antibiotics and suppressed the antibiotic-induced virulence.
Combination regimens for the prevention or reversal of antibiotic resistance are already a widespread method. They usually consist of two drugs, an antibiotic plus an antibiotic adjuvant [
12]. A combination of an antibiotic and an antibiotic adjuvant varies from a combination of two antibiotics. In the former, the adjuvant may have little to no in vitro action against bacteria, particularly at clinically relevant levels [
13]. The adjuvant’s principal function is to improve antibiotic activity. Additionally, these adjuvants may act as anti-resistant or antivirulence molecules by targeting specific pathways in the bacterial system. For example, adjuvants are reported to inhibit quorum sensing pathways in bacteria and prevented the formation of biofilm under in vitro and in vivo conditions [
7,
14]. Owing to these findings, the antivirulence properties of M21 were reassessed to address the bacterial virulence induced by antibiotics.
Virulence in
S. aureus is mediated by several factors such as surface proteins, toxins, and superantigens. Among these factors, surface proteins contribute to the initial adhesion and colonization of bacteria to the host surface and promote bacterial virulence. Once they colonize, toxins attack the host immune cells and favor the survival of
S. aureus in the host, leading to life-threatening infections [
15]. Thus, combating
S. aureus infection through virulence suppression by the non-antibiotic method is a promising alternative approach [
16,
17]. Thus, in the present study, the combination of antibiotics and an antivirulence agent was tested to control antibiotic-induced
S. aureus virulence.
β-lactams comprise a class of therapeutically significant antibiotics that are commonly employed as the first line of treatment for infectious diseases. However, more than 75% of patients with MRSA infections were shown to receive inappropriate agents for initial antimicrobial treatment [
3]. Due to the differences in tissue distribution, clearance, and metabolism rates, even when suitable antibiotics and dosages are utilized, the concentrations in certain organs of the human body might be lower than the MIC for some time periods [
18]. Antibiotics that are used for an extended period of time and at a low dose can develop resistance and increase pathogenicity. At non-lethal concentrations, β-lactam antibiotics induce the production of α-toxin and PVL toxin, toxic shock syndrome toxin (TSST), enterotoxins, and LukED, which should contribute to the alteration of therapy and, therefore, contribute to worse outcomes [
19,
20]. Induced bacterial adhesion by antibiotics commonly used in the management of
S. aureus infections may encourage invasive intracellular strains, which may have an important effect on the persistence and recurrence of infections [
3]. Numerous investigations established a relationship between methicillin resistance and poor clinical outcomes. MRSA strains were not shown to be considerably more virulent than MSSA strains [
21]. If antibiotic exposure is included, a resistant strain is more likely to be exposed to an unsuitable substance, resulting in greater pathogenicity [
22]. By introducing an antivirulence agent, we can control the antibiotics-induced virulence, to manage MRSA infection. A combination of β-lactam antibiotics and an antivirulence drug is preferable than using β-lactam antibiotics alone because of the risk of a worse outcome [
9].
The combination of antibiotics and antivirulence agents may harness the full potential of both agents [
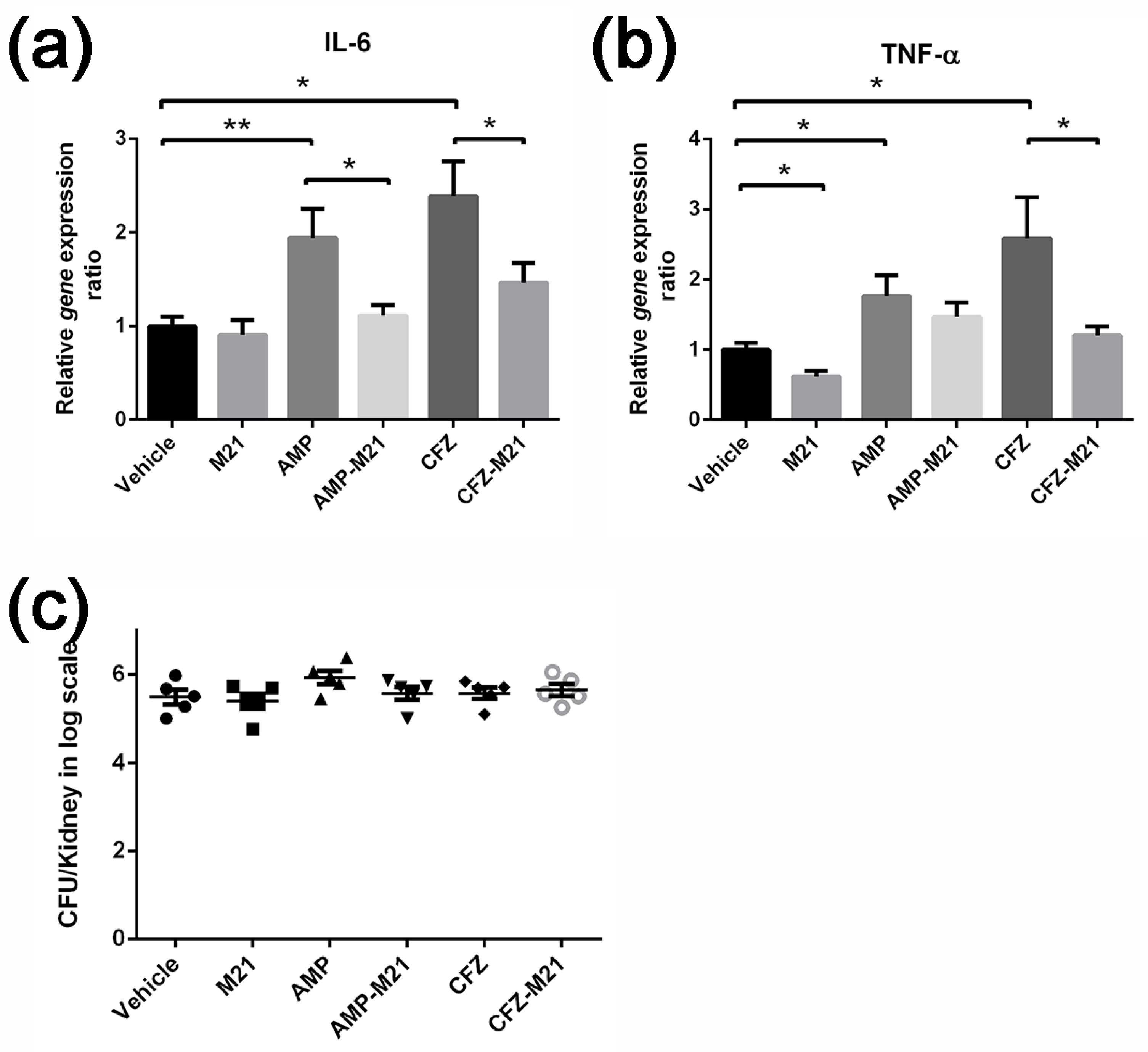
23]. A combination of the antivirulence agent with antibiotic was evaluated with various in vitro molecular, biochemical, and cellular assays and in vivo mouse models. Using the bacteremia model, we tested the induction of virulence by sub-inhibitory concentrations of ceftazidime (β-lactam antibiotics) and confirmed the worsening of infection by MRSA in mice. Infected mice treated with these antibiotics alone not only failed to reduce the bacterial load but also heightened the host immune response by enhancing bacterial virulence. Some toxins, such as PSMs, enhance the release of cytokines CXCL8, CCL20 IL-6, and TNF-α [
24]; α-toxin stimulate cytokines, such as nuclear factor-kB (NF-kB), IL-6, but not TNF-α [
25]; protein A will induce IL-8, TNF-α, and MIP-1α. In the present study, we demonstrated that the antivirulence molecule M21 could profoundly reduce α-toxin and protein A expression induced by subinhibitory concentrations of antibiotics [
26]. Additionally, the antivirulence agent M21, in combination with antibiotics, significantly improved the survival of mice in the bacteremia model and reduced the
S. aureus virulence expression in vivo. Additionally, by repressing the antibiotic-induced bacterial virulence, this novel combinatorial therapy reduced the host immune response during the infection. Thus, findings from the present study not only epitomize the dark side of antibiotics but also provide us an opportunity to reform our understanding of concealed virulence observed in
S. aureus strains. One of the intriguing approaches to address this challenge is the inclusion of antivirulence agents in the treatment regimen.
Currently, most of the research focused on the discovery of novel antibiotic adjuvants to sensitize resistant bacteria. After restoring the susceptibility to antibiotics, such as β-lactam antibiotics and aminoglycosides, the old drugs can be used again. Antibiotics adjuvants will show a synergistic effect with certain antibiotics by targeting the resistance genes or increasing the intake or inhibiting the efflux pumps. It is noteworthy to mention that the compound M21 did not show any synergistic effects with the tested antibiotics and reduced the antibiotic-induced virulence independently. This suggests that M21 does not interfere with antibiotics but works independently by suppressing multiple virulence factors. Hence, an antivirulence molecule such as M21 differs completely from traditional antibiotic adjuvants which are known to potentiate the activity of antibiotics. Thus, our findings using this small molecule, justify a novel perspective of developing antibiotics adjuvant and the need for antivirulence agents in suppressing antibiotic-induced virulence.
In concordance with these findings, our studies provide explicable evidence that the exacerbated infections caused by sub-inhibitory concentrations of antibiotics can be controlled by an antivirulence agent. We found that the compound M21 could overcome the virulence induction caused by ampicillin and ceftazidime (and presumably some β-lactam antibiotics as well). Thus, our studies illustrate the necessity for non-antibiotic-based antivirulence agents in suppressing S. aureus virulence. The application of these compounds in combination with antibiotics will possess high therapeutic values, and this could be a better choice to control S. aureus infection globally.
4. Materials and Methods
4.1. Bacterial Strains and Plasmids
The bacterial strains used in this study are listed in
Table 2. Brain heart infusion (BHI) broth and BHI agar plates were used to grow
S. aureus. Chloramphenicol was used at 10 µg/mL. Unless otherwise stated, all cultures were grown aerobically at 37 °C with shaking at 250 rpm and growth was monitored at 600 nm with a HITACHI U-2800 (Hitachi, Japan) spectrophotometer.
4.2. Minimum Inhibitory Concentration (MIC) Tests
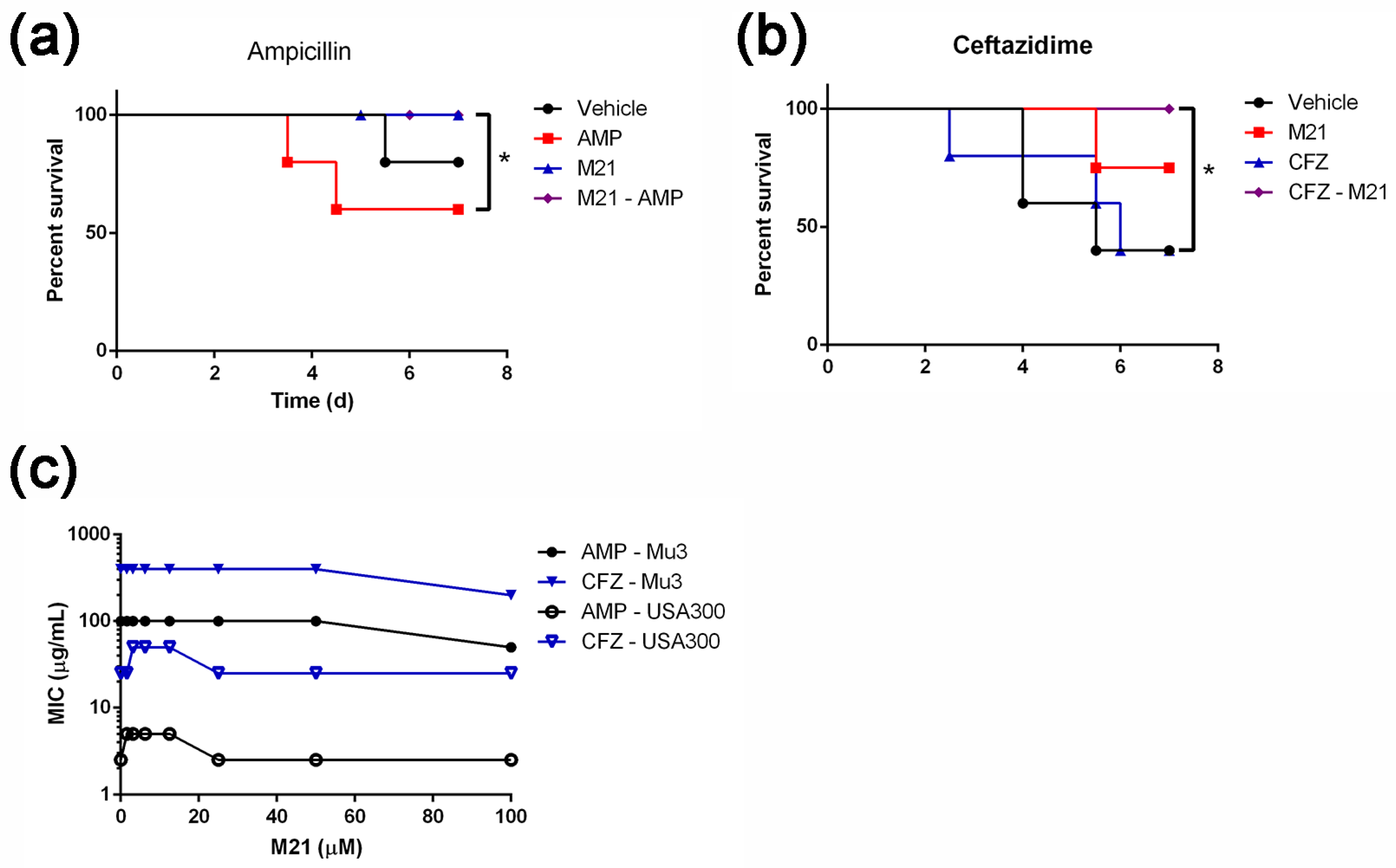
In 96-well plates, the MIC was measured by inoculating 5 × 104 S. aureus cells in 100 μL BHI medium with a serial dilution of antibiotics. After 18 h at 37 °C, the MIC was determined as the lowest concentration that resulted in a cell density of less than 0.01 OD at 620 nm, indicating no observable growth.
4.3. Disk Diffusion and Lux Assays
A single colony of bioluminescent
S. aureus strains containing plasmid pGL
hla (
Table 2) from BHI agar was resuspended in 200 μL sterile water, diluted to 75 mL 0.7% (
w/
v) soft agar, and plated on BHI agar. On the overlay, antibiotic discs (diameter 6 mm; Advantec Co., Tokyo, Japan) were placed, and the plates were incubated at 37 °C. After 20 h, inhibitory zones were determined, and luminescence was detected using a PE IVIS Spectrum in vivo imaging system (PerkinElmer, Hong Kong, China) [
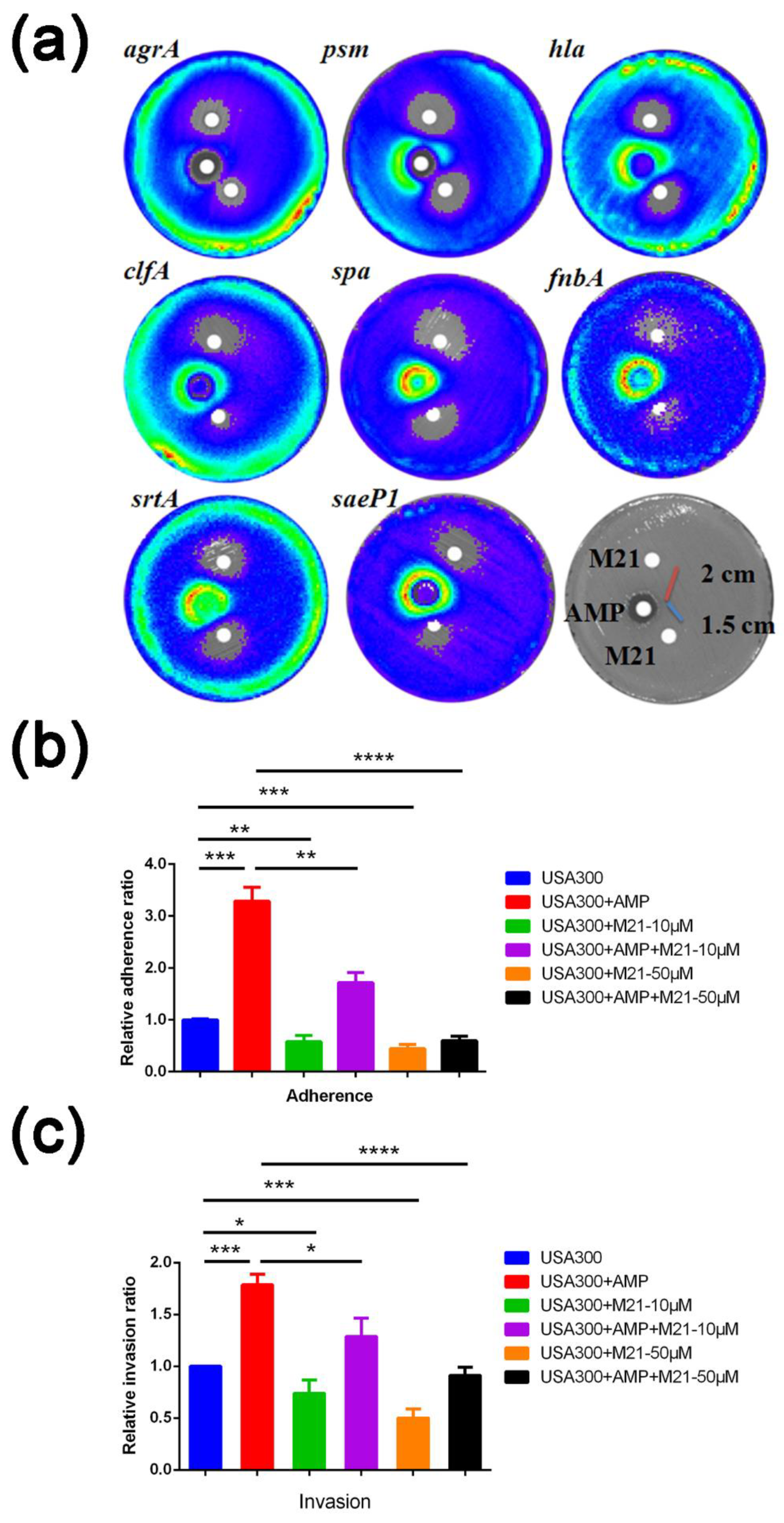
6]. When combining antibiotics and compounds, antibiotics (10 mM, 5 μL) and M21 (50 mM, 5 μL) were placed on the plates at a distance of 1.5 cm and 2 cm, respectively. When analyzing the interaction between cefoxitin and M21, we used a concentration of 4 mM of antibiotic and M21 and the distance between the discs was 1.5 cm.
4.4. Real-Time PCR to Verify Expression Levels
The preparation of total RNA from
S. aureus was performed using an RNA protection reagent according to the manufacturer’s instructions (Qiagen, Hilden, Germany) [
8].
To study the expression of mouse cytokines by q-PCR, we acquired cDNA as described above. The relative quantification of IL-6 and TNF-α transcripts was determined by the ratio of expression of target transcripts relative to Hprt gene (housekeeping gene). The sequences of primers for real-time PCR experiments are provided in
Table 3.
4.5. Adherence Assay and Invasion Assay
As previously described [
6], before inoculation, overnight bacterial cultures treated with ampicillin (0.2 g/mL) and/or compound M21 (10 μM or 50 μM) were washed three times with PBS (pH 7.4) and diluted to 10
7 CFU/mL with MEM medium (defined as the original bacterial CFU). A549 cells were seeded at a concentration of 2 × 10
5/mL in MEM onto a 24-well tissue culture plate (Greiner) for measuring adherence and invasion ratio. Briefly, confluent monolayers of A549 cells were cultivated overnight at 37 °C in 5% CO
2 to form. The next morning, the medium was removed and A549 cells were washed twice with 1 mL of PBS before being infected with 1 mL of the prepared bacterial inoculum. For the invasion experiment, after infecting A549 cells at 37 °C for two hours, the well supernatants were collected for bacterial count (defined as the total bacterial CFU). All wells were then washed three times with 1 mL PBS after which A549 cells were incubated for one hour at 37 °C in MEM containing gentamicin (100 μg/mL; Sigma-Aldrich, St. Louis, MO. USA) and lysostaphin (10 μg/mL; Sigma-Aldrich, St. Louis, MO. USA). Afterward, the wells were trypsinized with 150 μL of 0.25% trypsin-EDTA for 5 min, the cells in each well were carefully collected into tubes, and 400 μL of ice-cold 0.025% Triton X-100 was added to the tubes, which were then placed on ice.
The number of bacterial CFU released from lysed epithelial cells was evaluated by plating lysates on BHI agar plates (the invaded bacterial CFU). After infecting A549 cells at 37 °C for one hour for the adherence test, the medium was withdrawn, and the cells were washed three times with 1 mL PBS. The adherent bacterial CFU was then defined as the total number of attached and invasive bacteria liberated from the lysed epithelial cells.
The bacterial adhesion in each well is given as a percentage of the CFU in the inoculum that adhered to and penetrated the cells. The control wells had just medium as a pre-treatment (MEM). Using the formulae, adhesion and invasion were then standardized versus controls.
Each experiment was conducted three times, and all relative adhesion and invasion data were calculated and statistically evaluated using SigmaPlot software 11.0 (Sigma Plot Software, Jandel, Chicago, IL, USA) and the Student’s t-test. Based on the p values, statistical significance was assessed; p < 0.05 was deemed significant.
4.6. Western Blot
S. aureus strains were cultured in BHI broth. This allowed us to collect the supernatants at different time points. Supernatants of ampicillin or compound M21 treated S. aureus was isolated after 24 h, adjusted to an OD600 of 6.0, with BHI, and centrifuged. After boiling in Laemmli sample buffer, 5 μL of culture supernatant was loaded onto a 12% sodium dodecyl sulfate-polyacrylamide gel. Alpha-hemolysin was detected with rabbit anti-staphylococcal α-hemolysin antibody (1:20,000) (Sigma-Aldrich, St. Louis, MO. USA) and goat Horseradish Peroxidase (HRP)-conjugated anti-rabbit IgG (1:5000) (Sigma-Aldrich). Protein A was visualized with HRP-conjugated Rabbit anti-staphylococcal Spa antibody (1:20,000) (Abcam, Cambridge, MA, USA). The Western blot protocol was performed as described in the product guide for Amersham ECL Western blotting detection reagents (GE Healthcare, Buckinghamshire, UK).
4.7. Mouse Peritonitis Model
As previously described [
27], we kept the 6- to 8-week old BALB/c female mice in biosafety level 2 animal facility. Mice were housed in microisolator cages and received food and water ad libitum. All experimental protocols (CULATR 3055-13 and 3678-15) followed the standard operating procedures of the approved biosafety level 2 animal facilities and were approved by the Animal Ethics Committee.
Mid-exponential phase of S. aureus culture was washed twice with sterile PBS and resuspended again in PBS to obtain 1 × 108 cfu/100 μL. Mice were i.p. injected with 4 × 108 suspended early stationary phase S. aureus. After six hours post-infection, mice were randomized into two groups (n = 12). Every day, they were treated with 100 μL PBS or 8 mg/mL ampicillin in PBS subcutaneously (s.c.) twice (12-h interval). The third control group (n = 6), without bacterial infection, was treated only with ampicillin. Animals were daily monitored for symptoms of disease (body weight drop, inactivity, ruffled fur, and labored breath) and death.
To assess the antivirulence property of the compound M21, at the beginning of the experiment, mice were randomized into two groups (n = 10) consisting of vehicle control group (receiving PBS with 5% DMSO and 2% tween 80) and treatment group (receiving M21). Each group was treated with designated concentrations of compound M21 (3.85 mg/kg/dose) or vehicle control. Prior to infection, mice were pretreated with two doses of M21 one day before infection. On day 0, infection in mice was established by i.p. injection of 4 × 108 early stationary phase S. aureus. Two doses of compound M21 with an interval of 12 h were administrated. On day 3, 10 animals from each group were euthanized, kidneys, livers, and spleens were harvested, homogenized in PBS, and plated on BHI agar for counting the viable bacteria.
For studying the efficacy of combinatorial treatment, before the beginning of the experiment, mice were randomized into two groups (n = 12 for ampicillin group and n = 14 for combination treatment group). They were either administered with the designated concentrations of compound M21 or vehicle control. On day 0, mice were i.p. injected with 4 × 108 early stationary phase S. aureus mu3. On day 1, two doses of M21 pretreatment were given by i.p. injection. Six hours later, mice were treated with 100 μL of 8 mg/mL ampicillin in PBS subcutaneously. The ampicillin group was treated with the ampicillin by s.c. and vehicle group received the vehicle control by i.p., whereas, the combinatorial treatment group received ampicillin by s.c. and M21 by i.p. All these treatments comprised two dosages per day (12 h interval). The animals were daily monitored for symptoms of disease (body weight drop, inactivity, ruffled fur, and labored breath) and death.
On day 6, 16 animals were euthanized, kidneys, livers, and spleens were harvested, homogenized in PBS, and plated on BHIA for determining the bacterial viable count.
4.8. Mouse Bacteraemia Model
As previously described [
8],
S. aureus strain mu3 was cultured to the early exponential phase, washed twice with sterile PBS, and resuspended in PBS to attain a cell density of 1 × 10
8 CFU/100 μL. This bacterial suspension was used to establish the lethal mice infection model. The female BALB/c mice, 6–8 weeks, were infected through tail vein (i.v.) with
S. aureus and randomized into 4 groups consisting of 5–12 mice per group. One hour post-infection, mice were treated with designated concentrations of antibiotics (s.c.) or compound M21 (3.85 mg/kg/dose) (i.p.) or combination of antibiotics and M21 or vehicle, serving as control. Antibiotics such as ampicillin (40 mg/kg/dose), and ceftazidime (16.5 mg/kg/dose) were used for the treatment twice per day. To study the combinatorial effect of M21 and antibiotics, we used four groups of mice for vehicle, M21, antibiotic, and combined treatments, respectively. M21 or injection buffer treatments were performed twice per day at 12 h intervals. The survival was monitored according to the body condition scoring system.
For q-PCR studies, samples were obtained from mice that had undergone survival experiments. On day 2, animals from each group were euthanized and kidneys were collected. The kidney from each mouse was divided into two halves; one half of the kidney was stored in liquid nitrogen for RNA extraction, and the other half was homogenized in PBS and plated on BHIA for determining the bacterial viable count.
4.9. Statistics
Statistical analysis was performed using Graph Pad Prism version 7.0. All error bars depict the standard error of mean (SEM). Horizontal lines depict the mean. All replicates are biological (from different samples). Non-parametric tests (Mann–Whitney test) were used for significant analysis. For bacteraemia model, data were analyzed by survival analysis, and Log-rank (Mantel–Cox) test was used for significance analysis.


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}