
Antimicrobial Activities of Secondary Metabolites from Model Mosses

 , ,
, ,
Abstract
:1. Introduction
2. Results
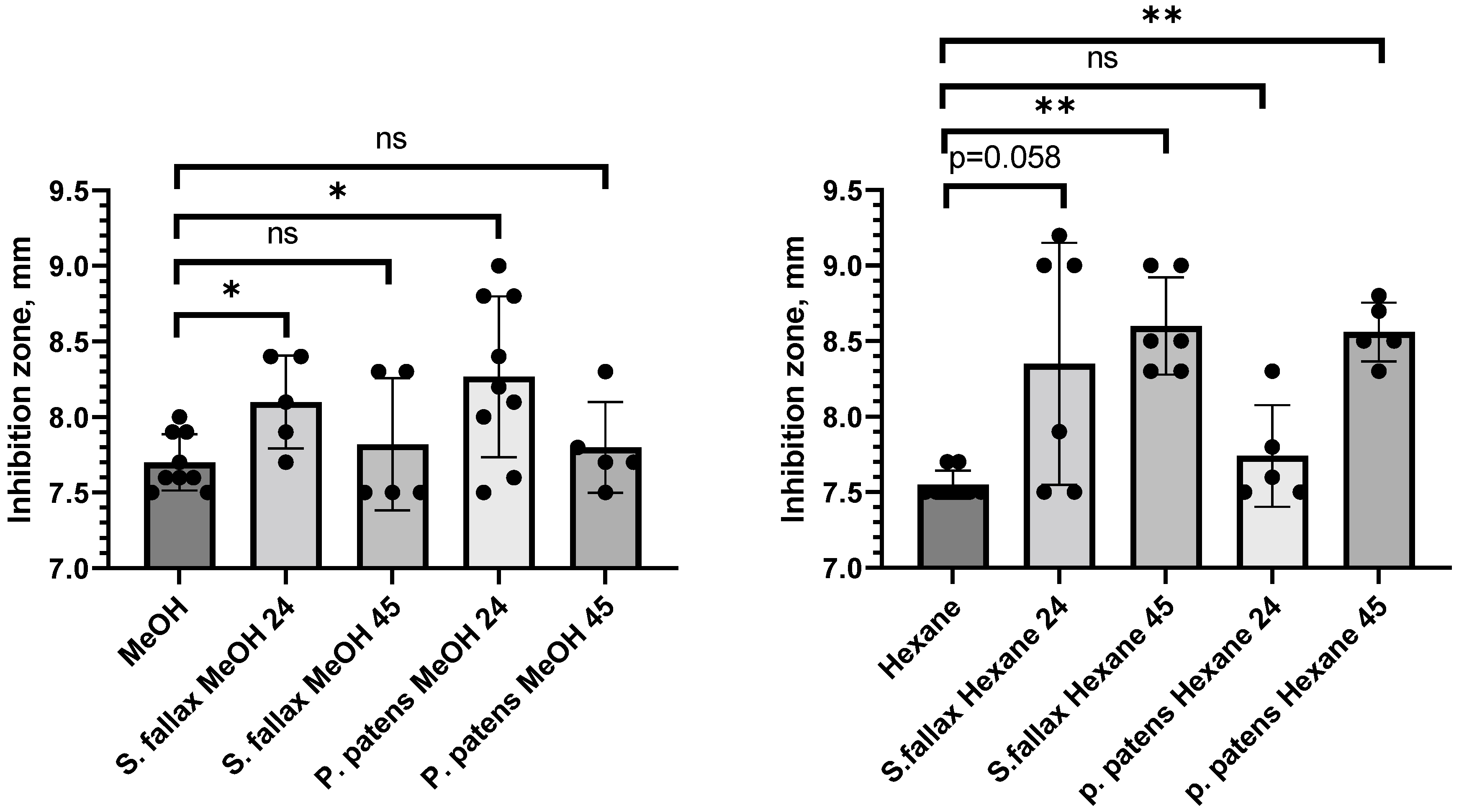
2.1. Determination of Antibacterial Activity of Intracellular Moss Metabolites
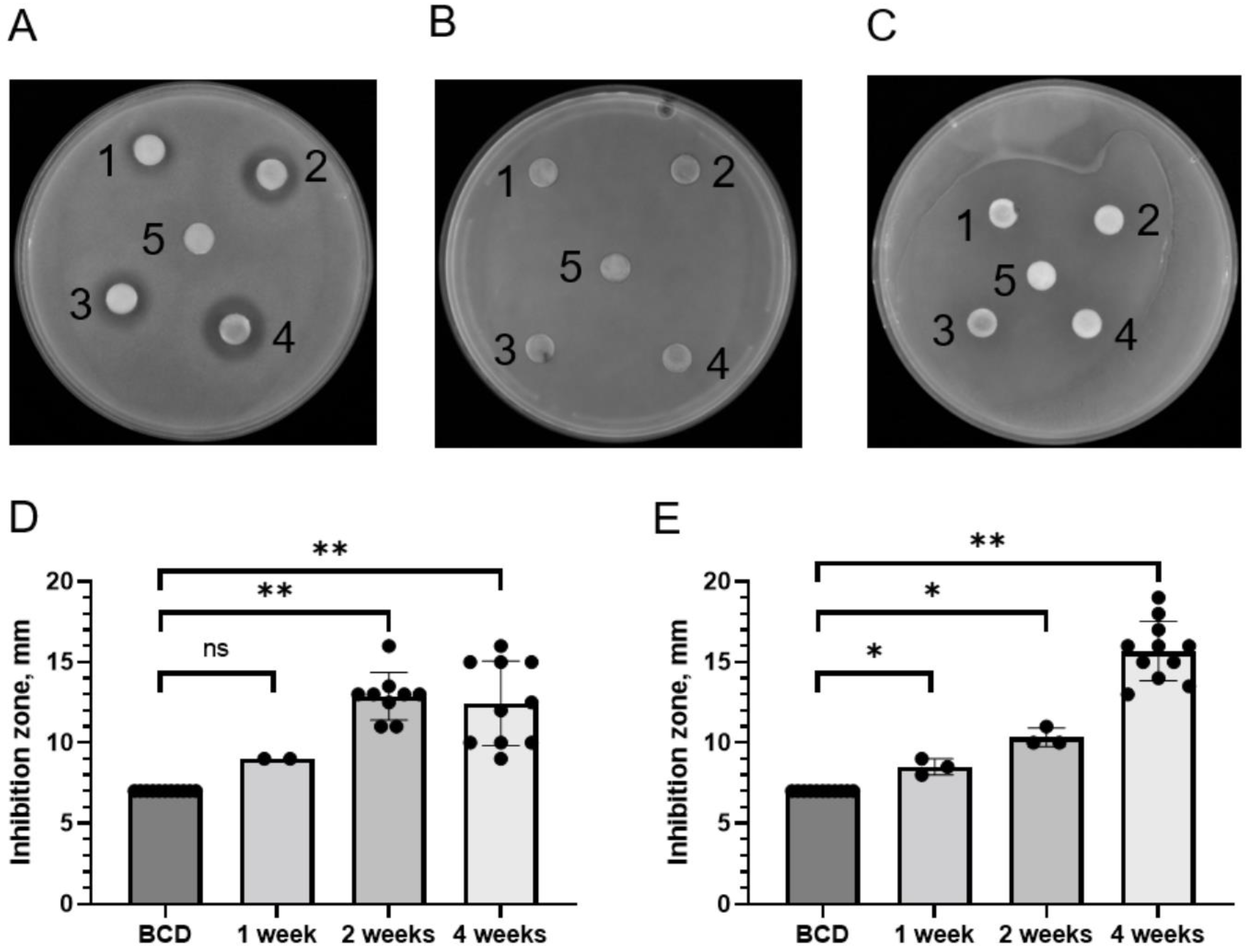
2.2. Identification of Antibacterial Activity in Moss Exudates
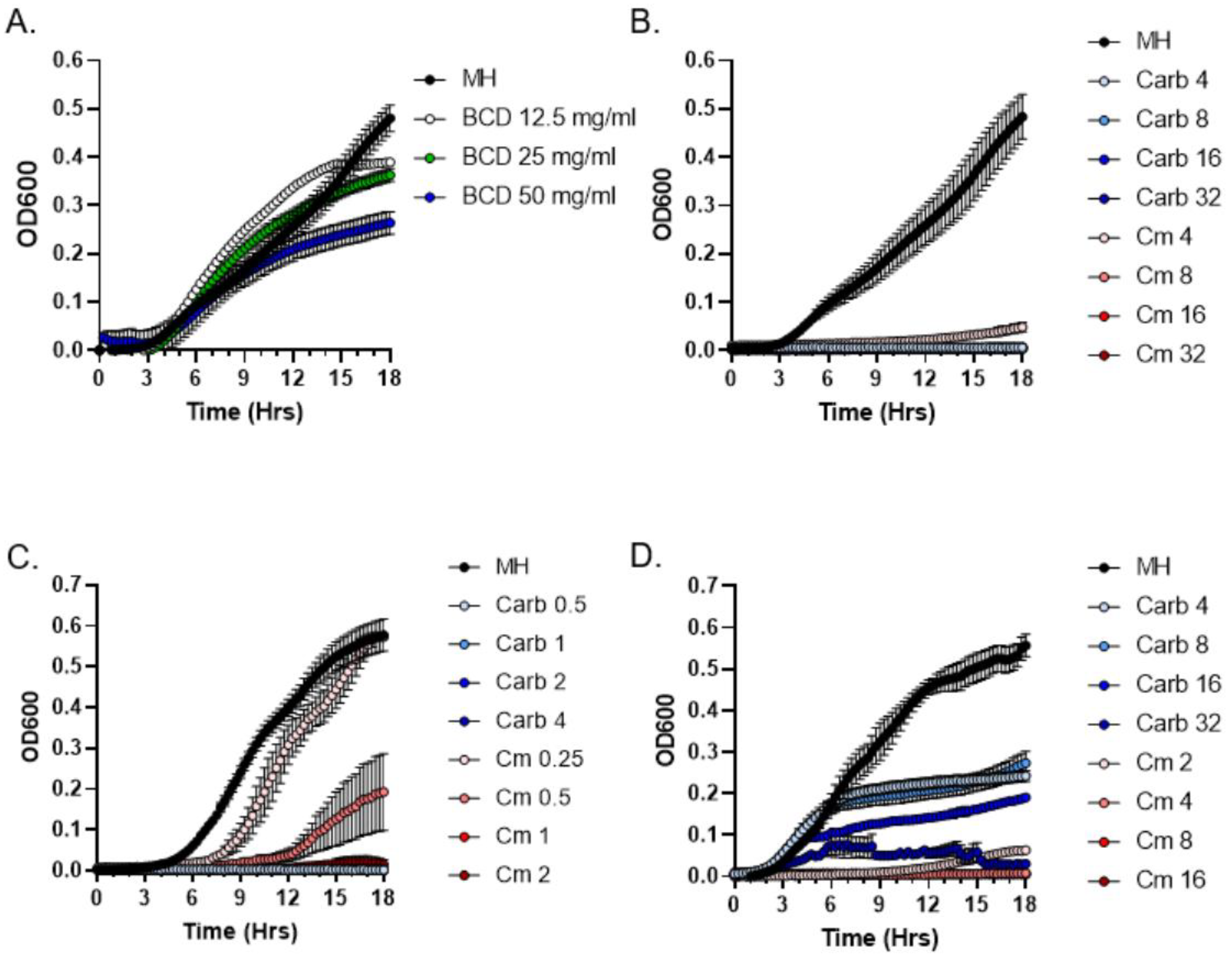
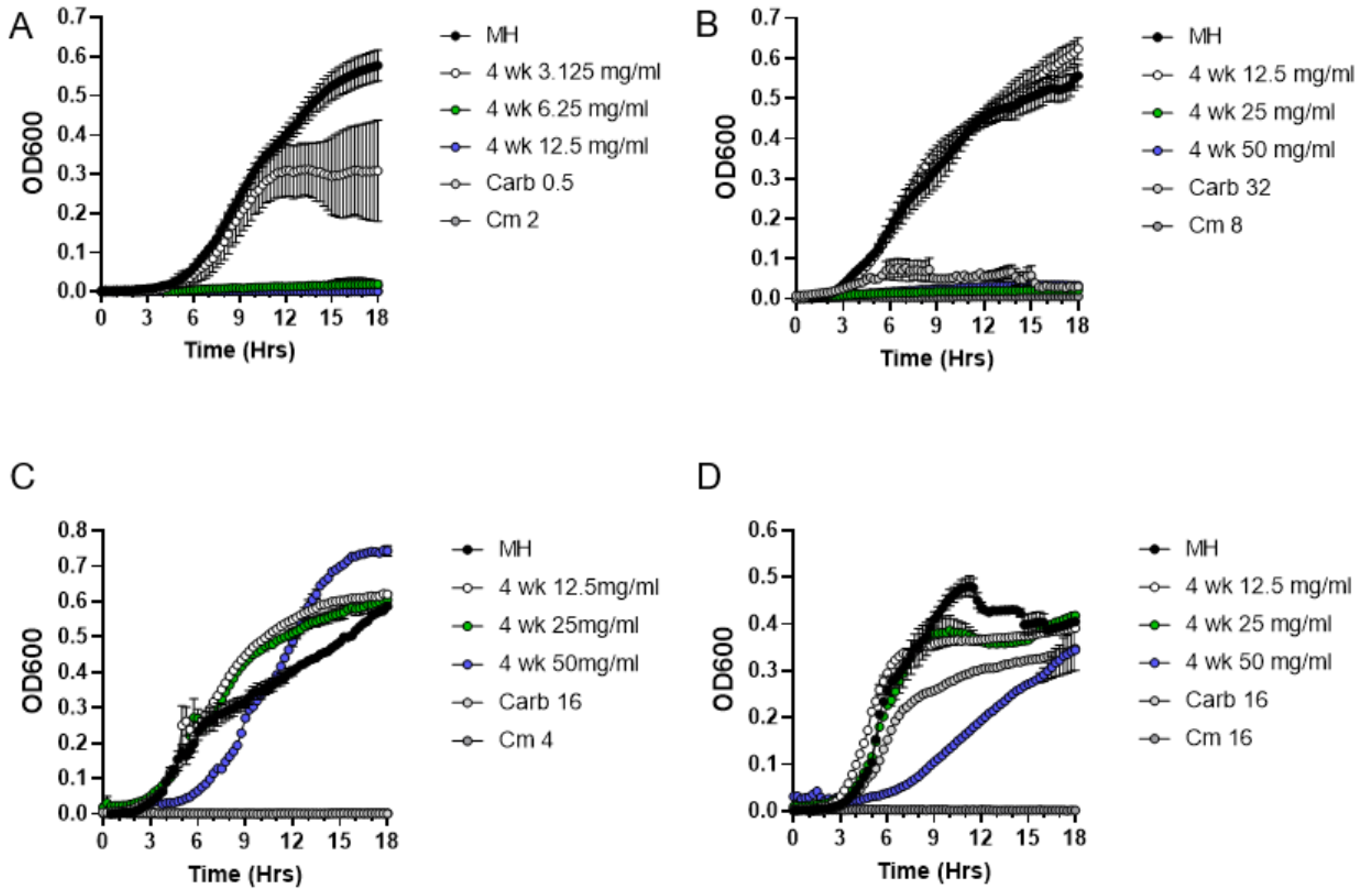
2.3. Quantitative Analysis of Antibacterial Activities of P. patens Exudate
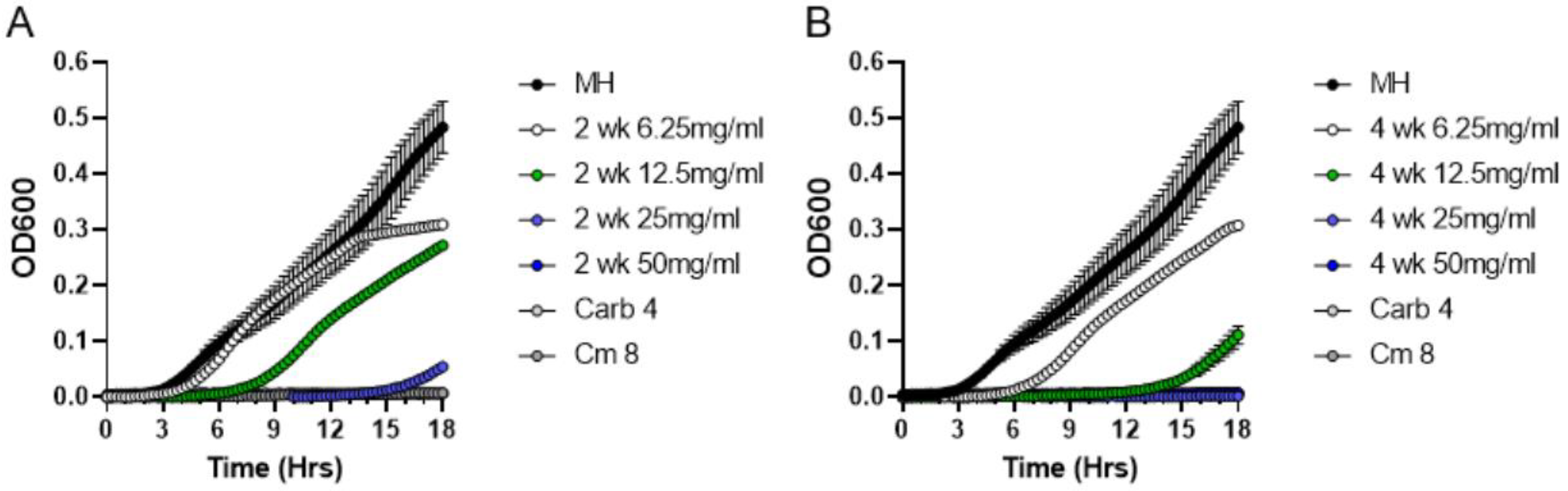
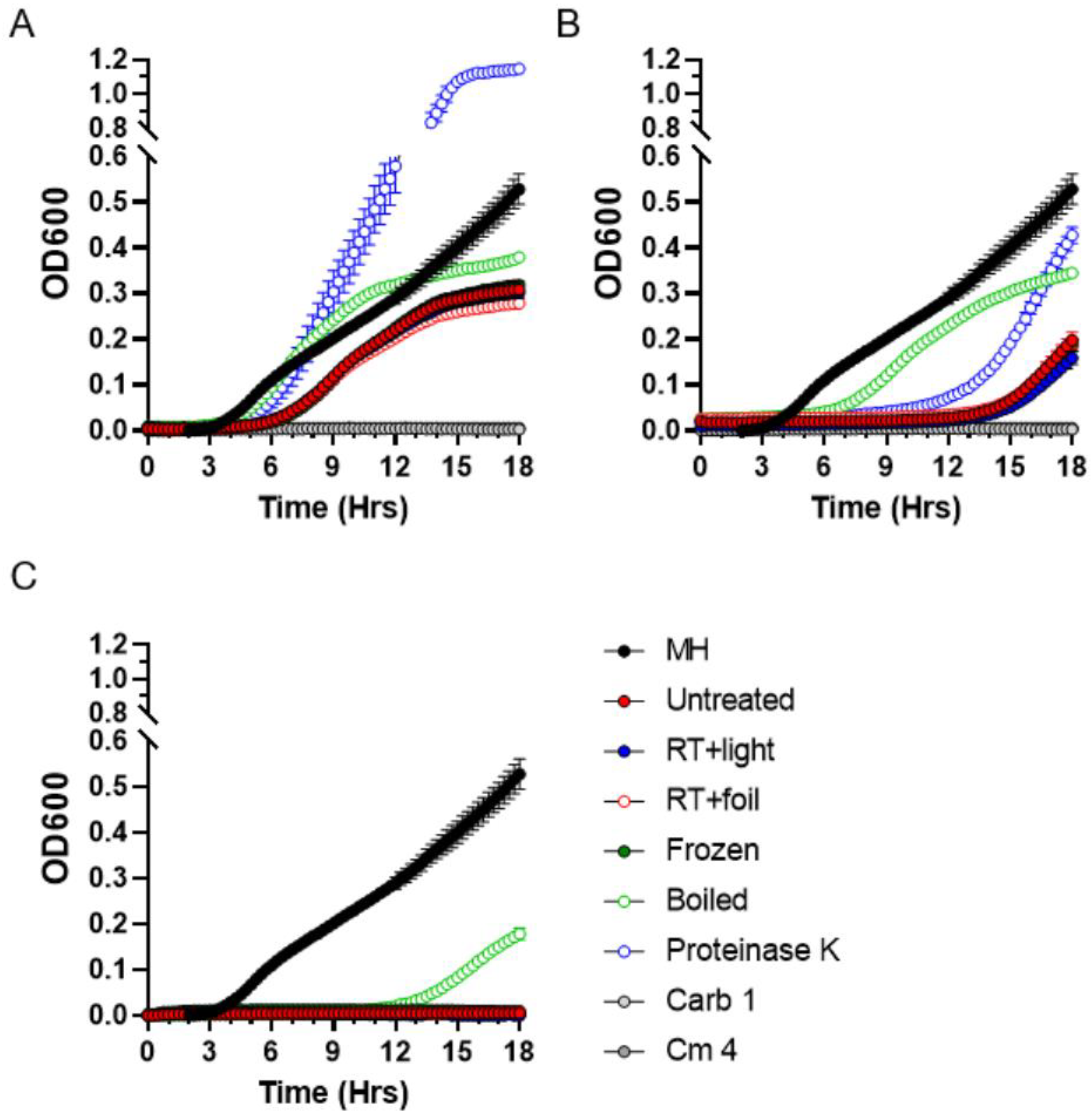
2.4. Stability of Antibacterial Compounds in Moss Exudates
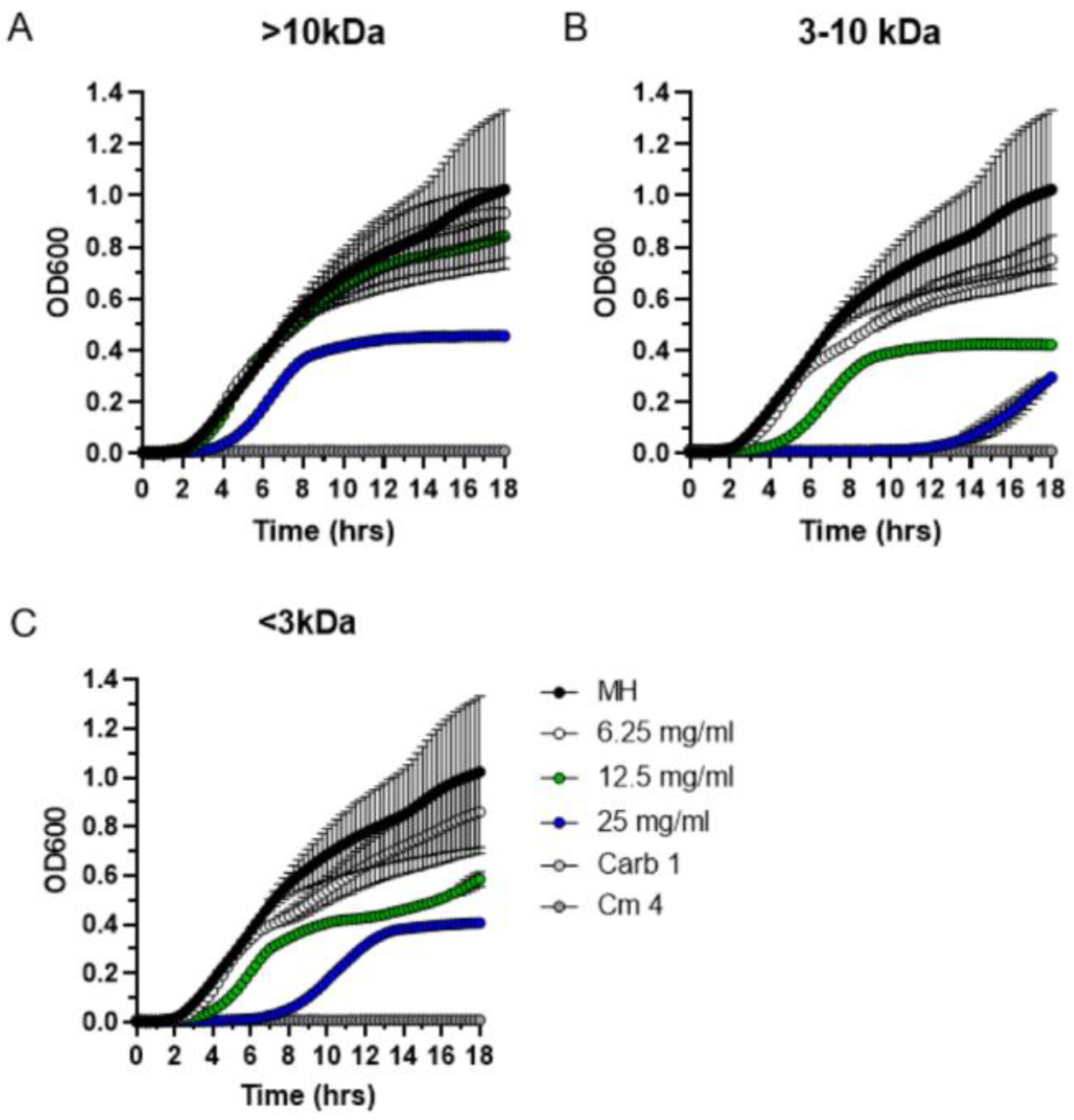
2.5. Size Fractionation of Bioactive Exudate Metabolites
3. Discussion
4. Materials and Methods
4.1. Moss Strains and Growth Conditions
4.2. Intracellular Metabolite Extraction
4.3. Preparation f Extracellular Metabolites from Moss Exudates
4.4. Tests for Antibacterial Activity
4.5. Metabolite Stability Test
4.6. Size Fractionation of Extracellular Moss Metabolites
4.7. Data analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- The SDG Knowledge Hub. WHO Identifies Top Health Challenges, Begins Five-Year Health Plan. 2019. Available online: http://sdg.iisd.org/news/who-identifies-top-health-challenges-begins-five-year-health-plan/ (accessed on 11 July 2022).
- Centers of Disease Control and Prevention. 2019 AR Threats Report. Available online: https://www.cdc.gov/drugresistance/biggest-threats.html (accessed on 11 July 2022).
- Cragg, G.M.; Kingston, D.G.I.; Newman, D.J. Anticancer Agents from Natural Products; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Erb, M.; Kliebensteinb, D.J. Plant Secondary Metabolites as Defenses, Regulators, Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Lelario, F.; Scrano, L.; De Franchi, S.; Bonomo, M.G.; Salzano, G.; Milan, S.; Milella, L.; Bufo, S.A. Identification and antimicrobial activity of most representative secondary metabolites from different plant species. Chem. Biol. Technol. Agric. 2018, 5, 13. [Google Scholar] [CrossRef]
- Alexandersson, E.; Ali, A.; Resjö, S.; Andreasson, E. Plant secretome proteomics. Front. Plant Sci. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fesenko, I.; Azarkina, R.; Kirov, I.; Kniazev, A.; Filippova, A.; Grafskaia, E.; Lazarev, V.; Zgoda, V.; Butenko, I.; Bukato, O.; et al. Phytohormone treatment induces generation of cryptic peptides with antimicrobial activity in the Moss Physcomitrella patens. BMC Plant Biol. 2019, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Lyapina, I.; Filippova, A.; Kovalchuk, S.; Ziganshin, R.; Mamaeva, A.; Lazarev, V.; Latsis, I.; Mikhalchik, E.; Panasenko, O.; Ivanov, O.; et al. Possible role of small secreted peptides (SSPs) in immune signaling in bryophytes. Plant Mol. Biol. 2021, 106, 123–143. [Google Scholar] [CrossRef]
- Frahm, J.P. Recent developments of commercial products from Bryophytes. Bryologist 2004, 107, 277–283. [Google Scholar] [CrossRef]
- Xie, C.-F.; Lou, H.-X. Secondary Metabolites in Bryophytes: An Ecological Aspect. Chem. Biodivers. 2009, 6, 303–312. [Google Scholar] [CrossRef]
- Veljić, M.; Tarbuk, M.; Marin, P.D.; Ćirić, A.; Soković, M.; Marin, M. Antimicrobial Activity of Methanol Extracts of Mosses from Serbia. Pharm. Biol. 2008, 46, 871–875. [Google Scholar] [CrossRef]
- Olofin, T.A.; Akande, A.O.; Oyetayo, V.O. Assessment of the antimicrobial properties of fractions obtained from bryophytes. J. Microbiol. Antimicrob. 2013, 5, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Asakawa, Y. Biologically active compounds from bryophytes. Pure Appl. Chem. 2007, 79, 557–580. [Google Scholar] [CrossRef]
- Mishra, R.; Pandey, V.K.; Chandra, R. Potential of Bryophytes as therapeutics. Int. J. Pharm. Sci. Res. 2014, 5, 3584–3593. [Google Scholar]
- Medina, R.; Johnson, M.G.; Liu, Y.; Wickett, N.J.; Shaw, A.J.; Goffinet, B. Phylogenomic delineation of Physcomitrium (Bryophyta: Funariaceae) based on targeted sequencing of nuclear exons and their flanking regions rejects the retention of Physcomitrella, Physcomitridium and Aphanorrhegma. J. Syst. Evol. 2019, 57, 404–417. [Google Scholar] [CrossRef] [Green Version]
- Erxleben, A.; Gessler, A.; Vervliet-Scheebaum, M.; Reski, R. Metabolite profiling of the moss Physcomitrella patens reveals evolutionary conservation of osmoprotective substances. Plant Cell Rep. 2012, 31, 427–436. [Google Scholar] [CrossRef]
- Asakawa, Y.; Ludwiczuk, A. Chemical Constituents of Bryophytes: Structures and Biological Activity. J. Nat. Prod. 2018, 81, 641–660. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.A.; Kamisugi, Y.; et al. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weston, D.J.; Turetsky, M.R.; Johnson, M.G.; Granath, G.; Lindo, Z.; Belyea, L.R.; Rice, S.K.; Hanson, D.T.; Engelhardt, K.A.M.; Schmutz, J.; et al. The Sphagnome Project: Enabling ecological and evolutionary insights through a genus-level se-quencing project. New Phytol. 2018, 217, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Decker, E.L.; Reski, R. Mosses in biotechnology. Curr. Opin. Biotechnol. 2020, 61, 21–27. [Google Scholar] [CrossRef]
- Rensing, S.A.; Goffinet, B.; Meyberg, R.; Wu, S.-Z.; Bezanilla, M. The Moss Physcomitrium (Physcomitrella) patens: A Model Organism for Non-Seed Plants. Plant Cell 2020, 32, 1361–1376. [Google Scholar] [CrossRef] [Green Version]
- Frank, W.; Ratnadewi, D.; Reski, R. Physcomitrella patens is highly tolerant against drought, salt and osmotic stress. Planta 2005, 220, 384–394. [Google Scholar] [CrossRef]
- Lehtonen, M.; Takikawa, Y.; Rönnholm, G.; Akita, M.; Kalkkinen, N.; Ahola-Iivarinen, E.; Somervuo, P.; Varjosalo, M.; Valkonen, J.P.T. Protein Secretome of Moss Plants (Physcomitrella patens) with Emphasis on Changes Induced by a Fungal Elicitor. J. Proteome Res. 2014, 13, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Klavina, L.; Spriņge, G.; Nikolajeva, V.; Martsinkevich, I.; Nakurte, I.; Dzabijeva, D.; Steinberga, I. Chemical Composition Analysis, Antimicrobial Activity and Cytotoxicity Screening of Moss Extracts (Moss Phytochemistry). Molecules 2015, 20, 17221–17243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cove, D.; Bezanilla, M.; Harries, P.; Quatrano, R. Mosses as Model Systems for the Study of Metabolism and Development. Annu. Rev. Plant Biol. 2006, 57, 497–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkins, J.; Shu, S.; Ishizaki, K.; Yamaoka, S.; Nishihama, R.; Nakamura, Y.; Berger, F.; et al. Insights into Land Plant Evolution Garnered from the Marchantia polymorpha Genome. Cell 2017, 171, 287–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heck, M.A.; Lüth, V.M.; Gessel, N.; Krebs, M.; Kohl, M.; Prager, A.; Joosten, H.; Decker, E.L.; Reski, R. Axenic in vitro cultivation of 19 peat moss (Sphagnum L.) species as a resource for basic biology, biotechnology, and paludiculture. New Phytol. 2021, 229, 861–876. [Google Scholar] [CrossRef]
- Vollár, M.; Gyovai, A.; Szűcs, P.; Zupkó, I.; Marschall, M.; Csupor-Löffler, B.; Bérdi, P.; Vecsernyés, A.; Csorba, A.; Liktor-Busa, E.; et al. Antiproliferative and Antimicrobial Activities of Selected Bryophytes. Molecules 2018, 23, 1520. [Google Scholar] [CrossRef] [Green Version]
- Choi, U.; Lee, C.-R. Distinct Roles of Outer Membrane Porins in Antibiotic Resistance and Membrane Integrity in Escherichia coli. Front. Microbiol. 2019, 10, 953. [Google Scholar] [CrossRef]
- Gruenheid, S.; Le Moual, H. Resistance to antimicrobial peptides in Gram-negative bacteria. FEMS Microbiol. Lett. 2012, 330, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Saul-McBeth, J.; Matson, J.S. A Periplasmic Antimicrobial Peptide-Binding Protein Is Required for Stress Survival in Vibrio cholerae. Front. Microbiol. 2019, 10, 161. [Google Scholar] [CrossRef]
- Xie, F.; Wang, Y.; Li, G.; Liu, S.; Cui, N.; Liu, S.; Langford, P.R.; Wang, C. The SapA Protein Is Involved in Resistance to Antimicrobial Peptide PR-39 and Virulence of Actinobacillus pleuropneumoniae. Front. Microbiol. 2017, 8, 811. [Google Scholar] [CrossRef]
- Bogomolnaya, L.M.; Tilvawala, R.; Elfenbein, J.R.; Cirillo, J.D.; Andrews-Polymenis, H.L. Linearized Siderophore Products Secreted via MacAB Efflux Pump Protect Salmonella enterica Serovar Typhimurium from Oxidative Stress. mBio 2020, 11, e00528-20. [Google Scholar] [CrossRef] [PubMed]
- Ashton, N.V.; Cove, D.J. The Isolation and Preliminary Characterisation of Auxotrophic and Analogue Resistant Mutants of the Moss, Physcomitreila patens. Molec. Gen. Genet. 1977, 154, 87–95. [Google Scholar] [CrossRef]
- Shirshikova, T.V.; Sierra-Bakhshi, C.G.; Kamaletdinova, L.K.; Matrosova, L.E.; Khabipova, N.N.; Evtugyn, V.G.; Khilyas, I.V.; Danilova, I.V.; Mardanova, A.M.; Sharipova, M.R.; et al. The ABC-Type Efflux Pump MacAB Is Involved in Protection of Serratia marcescens against Aminoglycoside Antibiotics, Polymyxins, and Oxidative Stress. mSphere 2021, 6, e00033-21. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moss Species | Growth Time, Days | Extraction Solvent | Metabolite Extraction Time, h |
|---|---|---|---|
| P. patens Gd | 10 | 80% methanol hexane | 24, 45 24, 45 |
| S. fallax MW | 30 | 80% methanol hexane | 24, 45 24, 45 |
| Moss Line | Extraction Solvent | Extraction Time, h | Growth Inhibition Zone, mm | ||||
|---|---|---|---|---|---|---|---|
| Negative Control a | P. syringae DC3000 | S. marcescens SM6 | E. coli TOP10 | S. aureus ATCC25923 | |||
| P. patens Gd | 80% methanol | 24 | 7.00 b | 8.30 ± 0.53 * | 7.00 | 8.03 ± 0.59 | 7.00 |
| 45 | 7.00 | 7.80 ± 0.30 | 7.00 | 8.13 ± 0.47 | 7.00 | ||
| Hexane | 24 | 7.00 | 7.74 ± 0.34 | 7.00 | 8.37 ± 0.40 | NA | |
| 45 | 7.00 | 8.56 ± 0.19 ** | 7.00 | 8.50 ± 0.50 | NA | ||
| S. fallax MW | 80% methanol | 24 | 7.00 | 8.10 ± 0.31 * | 7.00 | NA | NA |
| 45 | 7.00 | 7.82 ± 0.44 | 7.00 | NA | NA | ||
| Hexane | 24 | 7.00 | 8.35 ± 0.80 | 7.00 | NA | NA | |
| 45 | 7.00 | 8.60 ± 0.32 ** | 7.00 | NA | NA | ||
| P. patens Ecotype | Bacterial Growth Inhibition Zone, in mm | |||
|---|---|---|---|---|
| No Exudate Control | 1-Week-Old Moss Exudate | 2-Week-Old Moss Exudate | 4-Week-Old Moss Exudate | |
| Gd | 7 a | 9 ± 0.01 | 13.17 ± 1.27 ** | 12.97 ± 2.36 ** |
| Vx | 7 | 8.5 ± 0.71 * | 10.33 ± 0.58 * | 15.88 ± 1.65 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valeeva, L.R.; Dague, A.L.; Hall, M.H.; Tikhonova, A.E.; Sharipova, M.R.; Valentovic, M.A.; Bogomolnaya, L.M.; Shakirov, E.V. Antimicrobial Activities of Secondary Metabolites from Model Mosses. Antibiotics 2022, 11, 1004. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11081004
Valeeva LR, Dague AL, Hall MH, Tikhonova AE, Sharipova MR, Valentovic MA, Bogomolnaya LM, Shakirov EV. Antimicrobial Activities of Secondary Metabolites from Model Mosses. Antibiotics. 2022; 11(8):1004. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11081004
Chicago/Turabian StyleValeeva, Lia R., Ashley L. Dague, Mitchell H. Hall, Anastasia E. Tikhonova, Margarita R. Sharipova, Monica A. Valentovic, Lydia M. Bogomolnaya, and Eugene V. Shakirov. 2022. "Antimicrobial Activities of Secondary Metabolites from Model Mosses" Antibiotics 11, no. 8: 1004. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11081004