

A Comparison of the Immunometabolic Effect of Antibiotics and Plant Extracts in a Chicken Macrophage-like Cell Line during a Salmonella Enteritidis Challenge

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Minimal Inhibitory Concentrations of Antibiotics
2.2. Kinome peptide array Analysis
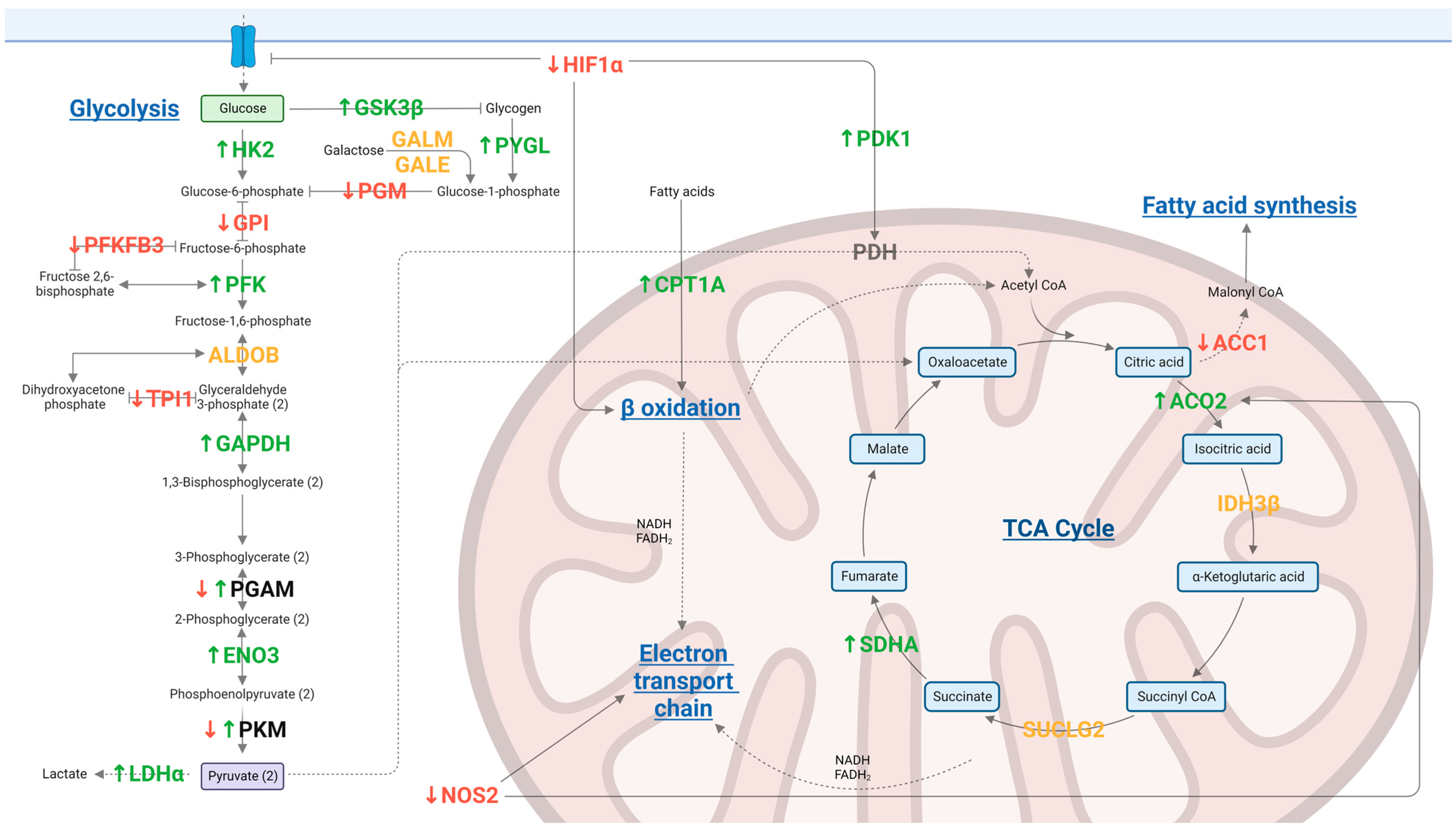
2.2.1. Kinome Peptide Array—Metabolic Modulation
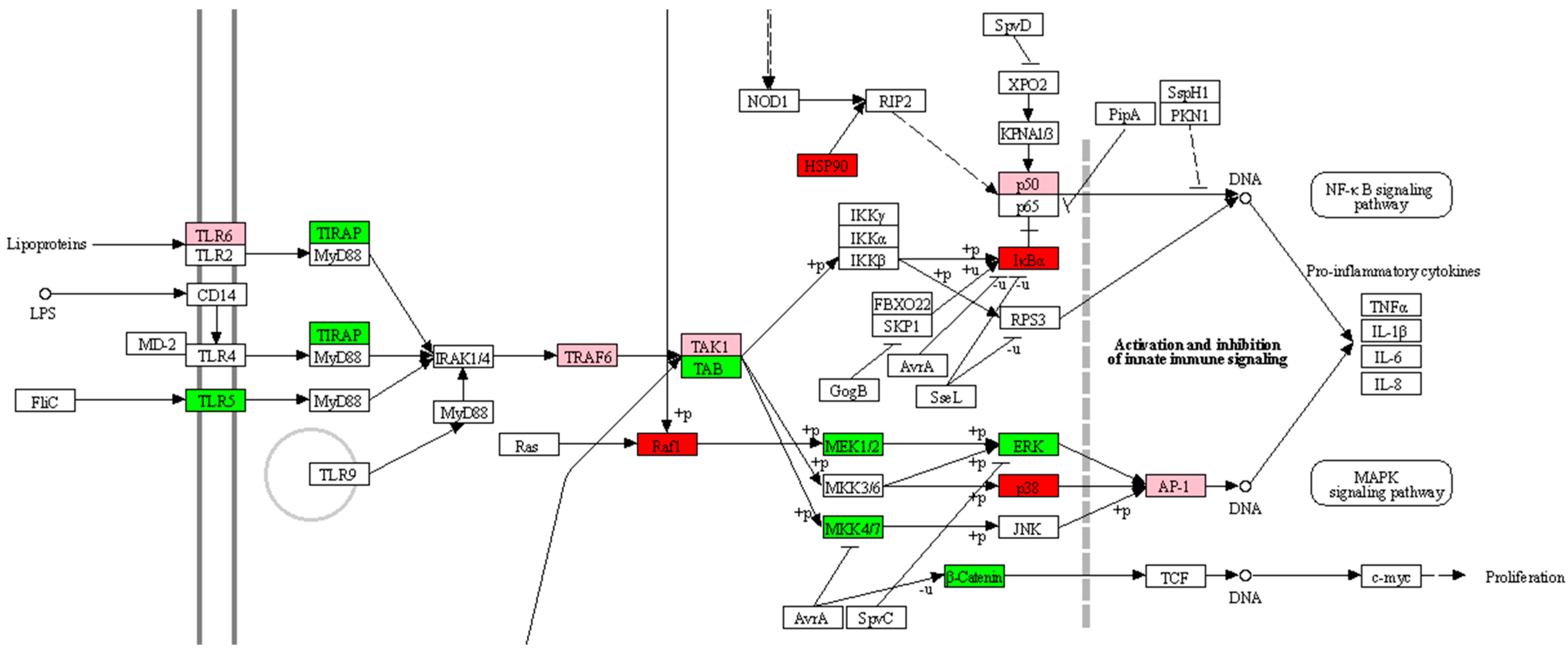
2.2.2. Kinome Peptide Array—Immune Modulation
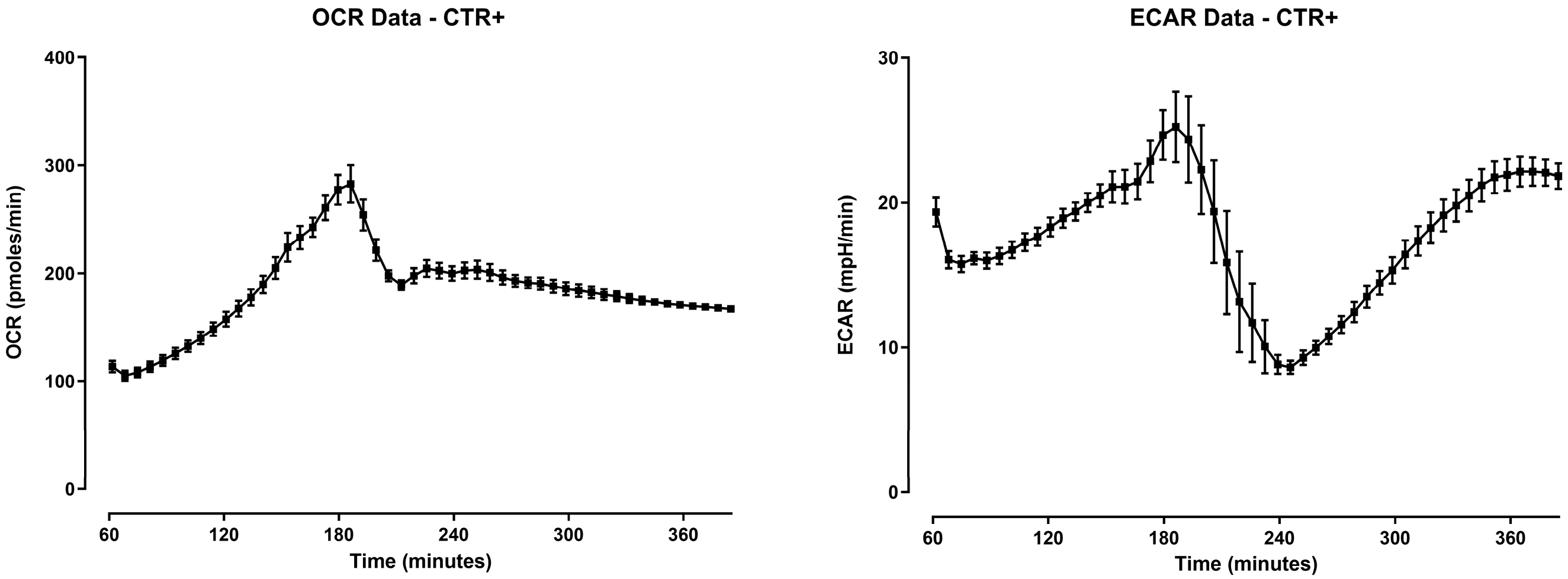
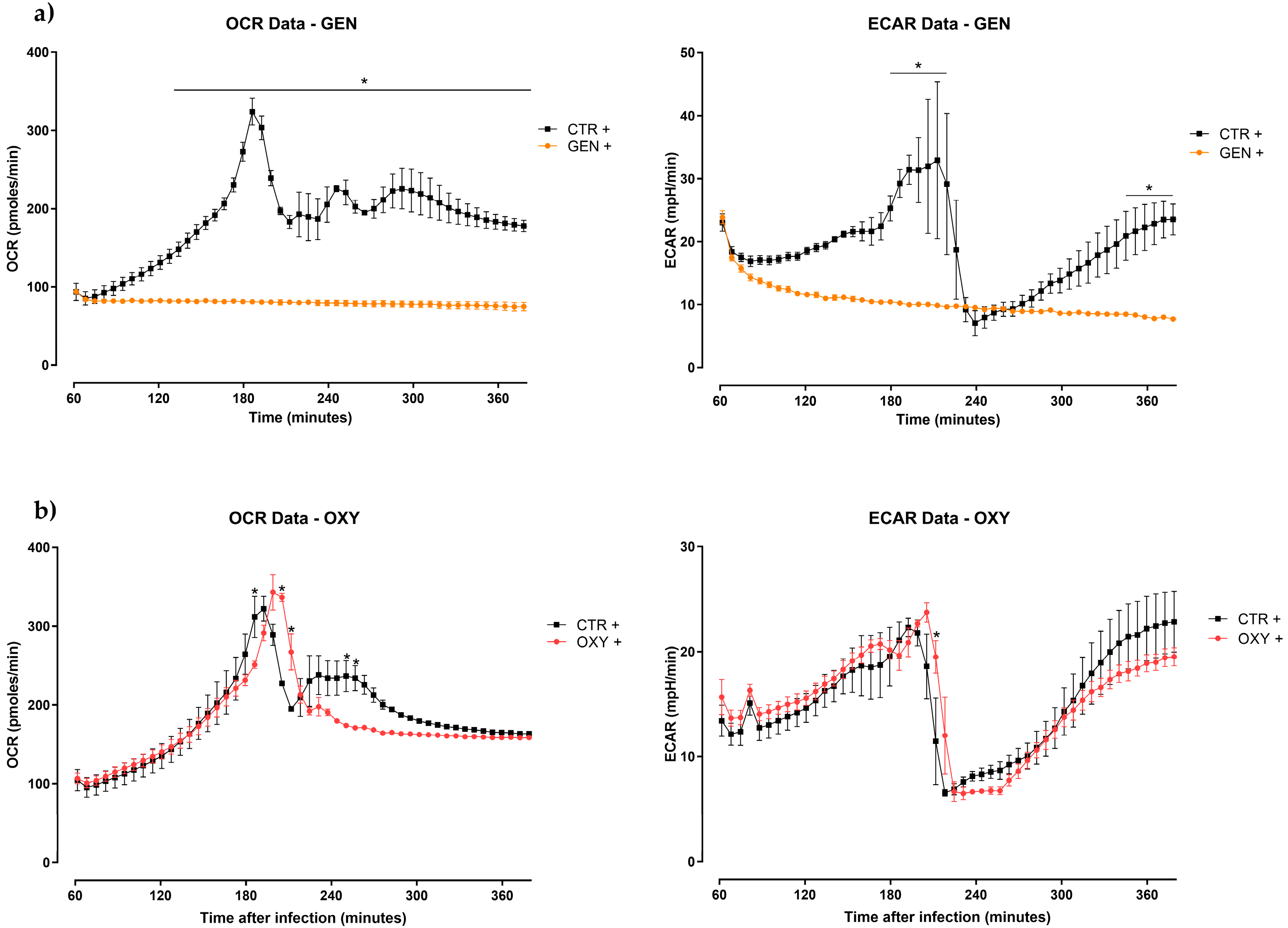
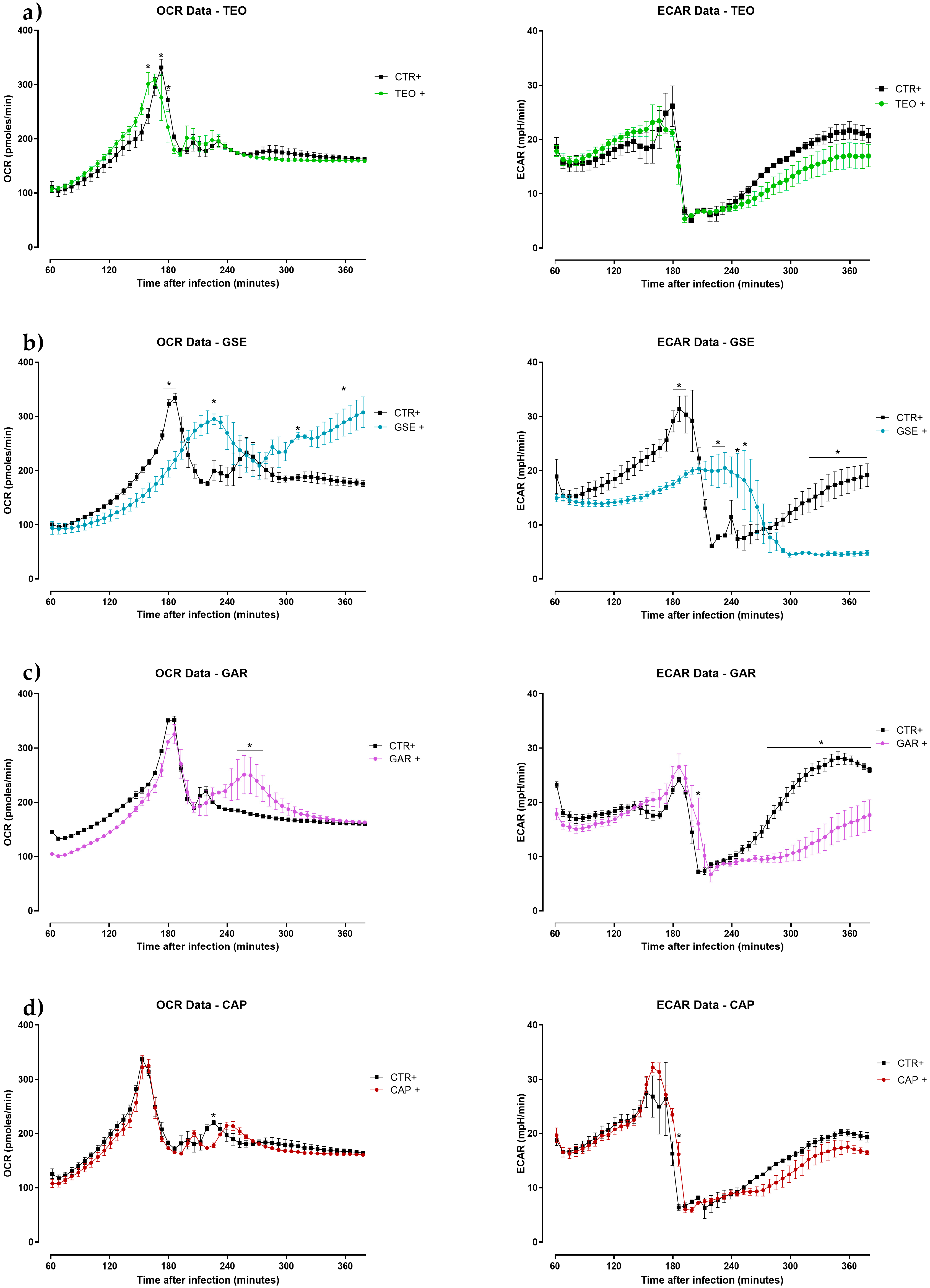
2.3. Seahorse Metabolic Assay Analysis
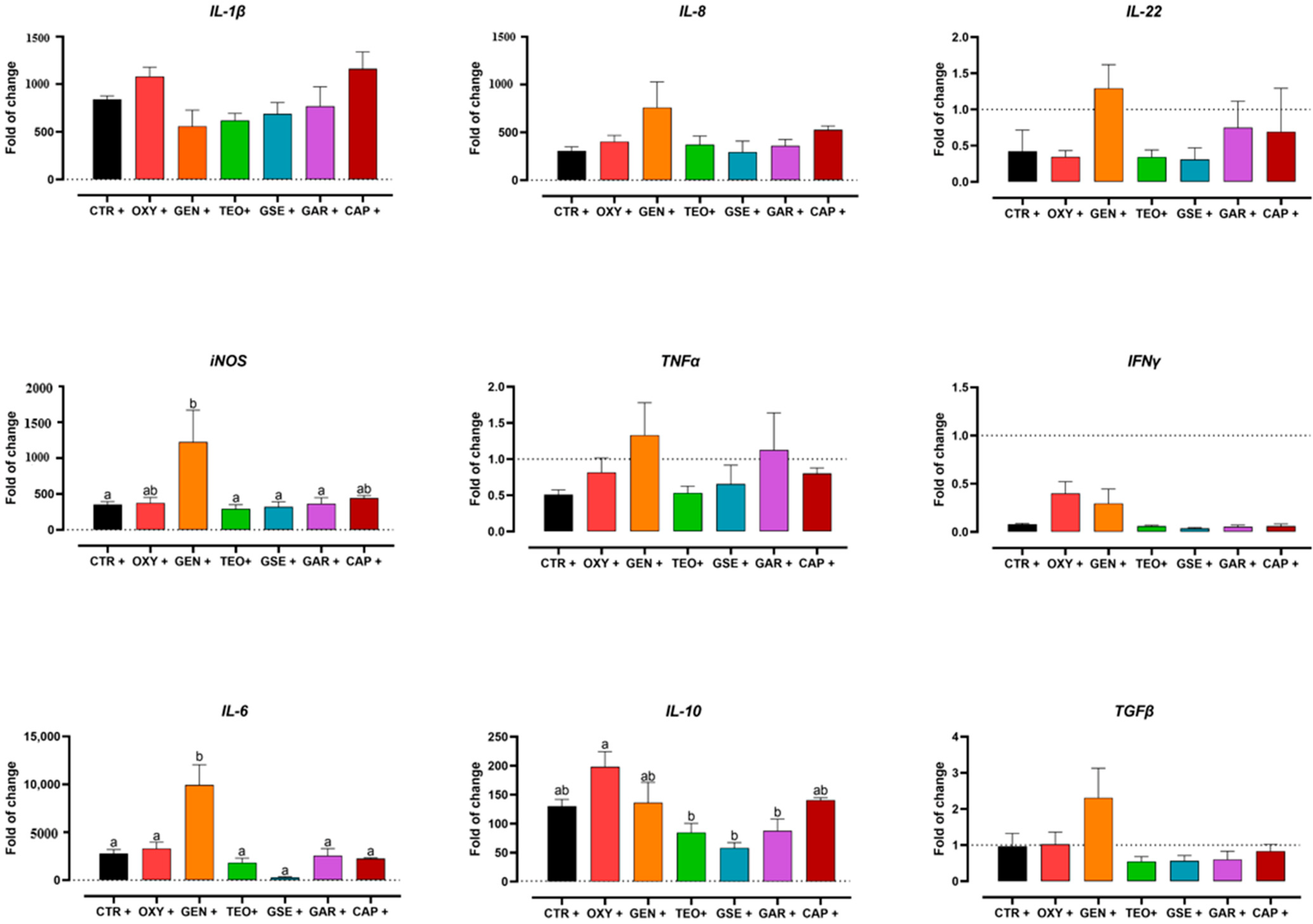
2.4. Gene Expression Analysis
3. Discussion
4. Conclusions
- Gentamicin led to a hybrid phenotype of macrophages, supported by an activation of immune pathways with increase of pro-inflammatory mediators and, at the same time, a less inflammatory metabolic response. Thereby, the effective antibiotic could help the host respond to the bacterial insult, avoiding the overreaction of its metabolic processes.
- The non-effective antibiotic (oxytetracycline) had no influence on macrophages’ metabolic profile and its role in immunomodulation was not totally supported by the kinome data.
- Grape seed extract switched macrophages to a less inflammatory status by promoting TCA cycle and oxidative phosphorylation. Even if several immune pathways were active according to kinome data, no evidence of expression of pro-inflammatory proteins was found.
- Garlic oil affected glycolysis and the metabolic and inflammatory status of macrophages, which was not supported by the kinome analysis.
- Thyme essential oil and capsicum oleoresin did not affect the immunometabolic profile of chicken macrophages. A possible explanation may be that the concentration was too low to elicit a response.
5. Materials and Methods
5.1. Cell Line, Bacterial Strain, and Culture Conditions
5.2. Chemical and Test Solutions
5.3. Salmonella Infection of HD11 Cells
5.3.1. Kinome Peptide Array Analysis
5.3.2. Seahorse Metabolic Assay Analysis
5.3.3. Gene Expression Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′ → 3′) | Accession Number | Reference |
|---|---|---|---|
| 28S | F: GGCGAAGCCAGAGGAAACT’ R: GACGACCGATTGCACGTC | X59733 | [66] |
| IL-1β | F: GCTCTACATGTCGTGTGTGATGAG R: TGTCGATGTCCCGCATGA | AJ245728 | [66] |
| IL-6 | F: GCTCGCCGGCTTCGA R: GGTAGGTCTGAAAGGCGAACAG | AJ250838 | [66] |
| IL-8 | F: GCCCTCCTCCTGGTTTCAG R: TGGCACCGCCAGCTCATT | AJ009800 | [66] |
| IL-10 | F: CATGCTGCTGGGCCTGAA R: CGTCTCCTTGATCTGCTTGATG | AJ621614 | [67] |
| IL-22 | F: CAGGAATCGCACCTACACCT R: CGGTTGTTCTCCCTGATGTT | NM_001199614 | [68] |
| iNOS | F: TTGGAAACCAAAGTGTGTAATATCTTG R: CCCTGGCCATGCGTACAT | NM_204961 | [69] |
| TFGβ | F: AGGATCTGCAGTGGAAGTGGAT R: CCCCGGGTTGTGTGTTGGT | M31160 | [66] |
| TNFα | F: CCCTACCCTGTCCCACAACC R: TGGGCGGTCATAGAACAGCA | XM_046927265.1 | |
| IFNγ | F: GTGAAGAAGGTGAAAGATATATCATGGA R: GCTTTGCGCTGGATTCTCA | Y07922 | [66] |
5.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- CDC. National Enteric Disease Surveillance: Salmonella Annual Report, 2016; CDC: Atlanta, GA, USA, 2016; p. 87.
- CDC. An Atlas of Salmonella in the United States, 1968–2011; CDC: Atlanta, GA, USA, 2013.
- Wessels, K.; Rip, D.; Gouws, P. Salmonella in Chicken Meat: Consumption, Outbreaks, Characteristics, Current Control Methods and the Potential of Bacteriophage Use. Foods 2021, 10, 1742. [Google Scholar] [CrossRef]
- Grilli, E.; Tugnoli, B.; Formigoni, A.; Massi, P.; Fantinati, P.; Tosi, G.; Piva, A. Microencapsulated Sorbic Acid and Nature-Identical Compounds Reduced Salmonella Hadar and Salmonella Enteritidis Colonization in Experimentally Infected Chickens. Poult. Sci. 2011, 90, 1676–1682. [Google Scholar] [CrossRef]
- Jung, Y.; Porto-Fett, A.C.S.; Shoyer, B.A.; Henry, E.; Shane, L.E.; Osoria, M.; Luchansky, J.B. Prevalence, Levels, and Viability of Salmonella in and on Raw Chicken Livers. J. Food Prot. 2019, 82, 834–843. [Google Scholar] [CrossRef]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef]
- Ijaz, A.; Veldhuizen, E.J.A.; Broere, F.; Rutten, V.P.M.G.; Jansen, C.A. The Interplay between Salmonella and Intestinal Innate Immune Cells in Chickens. Pathogens 2021, 10, 1512. [Google Scholar] [CrossRef]
- Gog, J.R.; Murcia, A.; Osterman, N.; Restif, O.; McKinley, T.J.; Sheppard, M.; Achouri, S.; Wei, B.; Mastroeni, P.; Wood, J.L.N.; et al. Dynamics of Salmonella Infection of Macrophages at the Single Cell Level. J. R. Soc. Interface 2012, 9, 2696–2707. [Google Scholar] [CrossRef]
- Kumar, V. Macrophages: The Potent Immunoregulatory Innate Immune Cells; IntechOpen: London, UK, 2019; ISBN 978-1-78984-645-4. [Google Scholar]
- Mills, C.D. M1 and M2 Macrophages: Oracles of Health and Disease. Crit. Rev. Immunol. 2012, 32, 463–488. [Google Scholar] [CrossRef]
- O’Neill, L.A.J.; Kishton, R.J.; Rathmell, J. A Guide to Immunometabolism for Immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Matthijs, M.G.R.; Haagsman, H.P.; Veldhuizen, E.J.A. Avian Pathogenic Escherichia Coli-Induced Activation of Chicken Macrophage HD11 cells. Dev. Comp. Immunol. 2018, 87, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Perry, F.; Johnson, C.; Aylward, B.; Arsenault, R.J. The Differential Phosphorylation-Dependent Signaling and Glucose Immunometabolic Responses Induced during Infection by Salmonella Enteritidis and Salmonella Heidelberg in Chicken Macrophage-like Cells. Microorganisms 2020, 8, 1041. [Google Scholar] [CrossRef]
- Tauber, S.C.; Nau, R. Immunomodulatory Properties of Antibiotics. Curr. Mol. Pharmacol. 2008, 1, 68–79. [Google Scholar]
- Sauer, A.; Peukert, K.; Putensen, C.; Bode, C. Antibiotics as Immunomodulators: A Potential Pharmacologic Approach for ARDS Treatment. Eur. Respir. Rev. 2021, 30, 210093. [Google Scholar] [CrossRef]
- Garrido-Mesa, J.; Algieri, F.; Rodríguez-Nogales, A.; Vezza, T.; Utrilla, M.P.; Garcia, F.; Chueca, N.; Rodríguez-Cabezas, M.E.; Garrido-Mesa, N.; Gálvez, J. Immunomodulatory Tetracyclines Ameliorate DNBS-Colitis: Impact on MicroRNA Expression and Microbiota Composition. Biochem. Pharmacol. 2018, 155, 524–536. [Google Scholar] [CrossRef]
- Garrido-Mesa, J.; Rodríguez-Nogales, A.; Algieri, F.; Vezza, T.; Hidalgo-Garcia, L.; Garrido-Barros, M.; Utrilla, M.P.; Garcia, F.; Chueca, N.; Rodriguez-Cabezas, M.E.; et al. Immunomodulatory Tetracyclines Shape the Intestinal Inflammatory Response Inducing Mucosal Healing and Resolution. Br. J. Pharmacol. 2018, 175, 4353–4370. [Google Scholar] [CrossRef]
- Siplivaia, L.E.; Shetsova, E.M.; Lazarev, A.I.; Prokopenko, L.G. The immunomodulating action of aminoglycoside antibiotics via different administration technics. Antibiot. Khimioterapiia Antibiot. Chemoterapy Sic 1999, 44, 29–32. [Google Scholar]
- Umeki, S. Anti-Inflammatory Action of Gentamycin through Inhibitory Effect on Neutrophil NADPH Oxidase Activity. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1995, 110, 817–821. [Google Scholar] [CrossRef]
- Zeng, Z.; Zhang, S.; Wang, H.; Piao, X. Essential Oil and Aromatic Plants as Feed Additives in Non-Ruminant Nutrition: A Review. J. Anim. Sci. Biotechnol. 2015, 6, 7. [Google Scholar] [CrossRef]
- Frankič, T.; Voljč, M.; Salobir, J.; Rezar, V. Use of Herbs and Spices and Their Extracts in Animal Nutrition. Acta Agric. Slov. 2009, 94, 95–102. [Google Scholar]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single Components of Botanicals and Nature-Identical Compounds as a Non-Antibiotic Strategy to Ameliorate Health Status and Improve Performance in Poultry and Pigs. Nutr. Res. Rev. 2020, 33, 218–234. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and Thyme Essential Oil—New Insights into Selected Therapeutic Applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Dey, S.; Marbaniang, D.; Pal, P.; Ray, S.; Mazumder, B. Grape Seed Extract: Having a Potential Health Benefits. J. Food Sci. Technol. 2020, 57, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, A. Revealing the Therapeutic Uses of Garlic (Allium Sativum) and Its Potential for Drug Discovery. Sci. World J. 2021, 2021, 8817288. [Google Scholar] [CrossRef] [PubMed]
- Sricharoen, P.; Lamaiphan, N.; Patthawaro, P.; Limchoowong, N.; Techawongstien, S.; Chanthai, S. Phytochemicals in Capsicum Oleoresin from Different Varieties of Hot Chilli Peppers with Their Antidiabetic and Antioxidant Activities Due to Some Phenolic Compounds. Ultrason. Sonochem. 2017, 38, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Pilkis, S.J.; El-Maghrabi, M.R.; McGrane, M.; Pilkis, J.; Claus, T.H. Regulation by Glucagon of Hepatic Pyruvate Kinase, 6-Phosphofructo 1-Kinase, and Fructose-1,6-Bisphosphatase. Fed. Proc. 1982, 41, 2623–2628. [Google Scholar] [PubMed]
- Lee, W.-H.; Choi, J.-S.; Byun, M.-R.; Koo, K.-T.; Shin, S.; Lee, S.-K.; Surh, Y.-J. Functional Inactivation of Triosephosphate Isomerase through Phosphorylation during Etoposide-Induced Apoptosis in HeLa Cells: Potential Role of Cdk2. Toxicology 2010, 278, 224–228. [Google Scholar] [CrossRef]
- Litwack, G. Chapter 8—Glycolysis and Gluconeogenesis. In Human Biochemistry; Litwack, G., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 183–198. ISBN 978-0-12-383864-3. [Google Scholar]
- Fan, Y.; Wang, J.; Xu, Y.; Wang, Y.; Song, T.; Liang, X.; Jin, F.; Su, D. Anti-Warburg Effect by Targeting HRD1-PFKP Pathway May Inhibit Breast Cancer Progression. Cell Commun. Signal. CCS 2021, 19, 18. [Google Scholar] [CrossRef]
- Cao, C.; Leng, Y.; Liu, X.; Yi, Y.; Li, P.; Kufe, D. Catalase Is Regulated by Ubiquitination and Proteosomal Degradation. Role of the c-Abl and Arg Tyrosine Kinases. Biochemistry 2003, 42, 10348–10353. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for Taxonomy-Based Analysis of Pathways and Genomes. Nucleic Acids Res. 2022, 51, D587–D592. [Google Scholar] [CrossRef]
- Xu, Q.; Tu, J.; Dou, C.; Zhang, J.; Yang, L.; Liu, X.; Lei, K.; Liu, Z.; Wang, Y.; Li, L.; et al. HSP90 Promotes Cell Glycolysis, Proliferation and Inhibits Apoptosis by Regulating PKM2 Abundance via Thr-328 Phosphorylation in Hepatocellular Carcinoma. Mol. Cancer 2017, 16, 178. [Google Scholar] [CrossRef]
- Nadeau, V.; Guillemette, S.; Bélanger, L.-F.; Jacob, O.; Roy, S.; Charron, J. Map2k1 and Map2k2 Genes Contribute to the Normal Development of Syncytiotrophoblasts during Placentation. Development 2009, 136, 1363–1374. [Google Scholar] [CrossRef]
- Toschi, A.; Piva, A.; Grilli, E. Phenol-Rich Botanicals Modulate Oxidative Stress and Epithelial Integrity in Intestinal Epithelial Cells. Animals 2022, 12, 2188. [Google Scholar] [CrossRef]
- Charles, A.; Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M.J. Principles of Innate and Adaptive Immunity. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Boothe, D.M. Principles of Antimicrobial Therapy. Vet. Clin. N. Am. Small Anim. Pract. 2006, 36, 1003–1047. [Google Scholar] [CrossRef]
- NARMS. CDC Antibiotics Tested by NARMS. Available online: https://www.cdc.gov/narms/antibiotics-tested.html (accessed on 17 October 2022).
- Fuster, J.J.; Walsh, K. The Good, the Bad, and the Ugly of Interleukin-6 Signaling. EMBO J. 2014, 33, 1425–1427. [Google Scholar] [CrossRef]
- Rajpoot, S.; Wary, K.K.; Ibbott, R.; Liu, D.; Saqib, U.; Thurston, T.L.M.; Baig, M.S. TIRAP in the Mechanism of Inflammation. Front. Immunol. 2021, 12, 697588. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Do, H.T.T.; Bui, B.P.; Sim, S.; Jung, J.-K.; Lee, H.; Cho, J. Anti-Inflammatory and Anti-Migratory Activities of Isoquinoline-1-Carboxamide Derivatives in LPS-Treated BV2 Microglial Cells via Inhibition of MAPKs/NF-ΚB Pathway. Int. J. Mol. Sci. 2020, 21, 2319. [Google Scholar] [CrossRef]
- Nair, D.V.T.; Venkitanarayanan, K.; Kollanoor Johny, A. Antibiotic-Resistant Salmonella in the Food Supply and the Potential Role of Antibiotic Alternatives for Control. Foods 2018, 7, 167. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Wada, T.; Suzuki, F.; Takagi, T.; Hasegawa, J.; Handa, H. Casein Kinase II Interacts with the BZIP Domains of Several Transcription Factors. Nucleic Acids Res. 1998, 26, 3854–3861. [Google Scholar] [CrossRef]
- Sanin, D.E.; Prendergast, C.T.; Mountford, A.P. IL-10 Production in Macrophages Is Regulated by a TLR-Driven CREB-Mediated Mechanism That Is Linked to Genes Involved in Cell Metabolism. J. Immunol. 2015, 195, 1218–1232. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Niisato, M.; Kawasaki, Y.; Karaman, S.; Robciuc, M.R.; Shibata, Y.; Ishida, Y.; Nishio, R.; Masuda, T.; Sugai, T.; et al. VEGF-C/VEGFR-3 Signaling in Macrophages Ameliorates Acute Lung Injury. Eur. Respir. J. 2021, 59, 2100880. [Google Scholar] [CrossRef] [PubMed]
- Bharhani, M.S.; Borojevic, R.; Basak, S.; Ho, E.; Zhou, P.; Croitoru, K. IL-10 Protects Mouse Intestinal Epithelial Cells from Fas-Induced Apoptosis via Modulating Fas Expression and Altering Caspase-8 and FLIP Expression. Am. J. Physiol.-Gastrointest. Liver Physiol. 2006, 291, G820–G829. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The Regulation of IL-10 Production by Immune Cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef]
- Kumar, K.G.S.; Krolewski, J.J.; Fuchs, S.Y. Phosphorylation and Specific Ubiquitin Acceptor Sites Are Required for Ubiquitination and Degradation of the IFNAR1 Subunit of Type I Interferon Receptor. J. Biol. Chem. 2004, 279, 46614–46620. [Google Scholar] [CrossRef]
- Xia, C.; Anderson, P.; Hahm, B. Viral Dedication to Vigorous Destruction of Interferon Receptors. Virology 2018, 522, 19–26. [Google Scholar] [CrossRef]
- Srikanth, C.V.; Wall, D.M.; Maldonado-Contreras, A.; Shi, H.; Zhou, D.; Demma, Z.; Mumy, K.L.; McCormick, B.A. Salmonella Pathogenesis and Processing of Secreted Effectors by Caspase-3. Science 2010, 330, 390–393. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Lynch, J.P.; Lesser, C.F. A Host Metabolite Promotes Salmonella Survival. Science 2021, 371, 344–345. [Google Scholar] [CrossRef]
- DeWaal, D.; Nogueira, V.; Terry, A.R.; Patra, K.C.; Jeon, S.-M.; Guzman, G.; Au, J.; Long, C.P.; Antoniewicz, M.R.; Hay, N. Hexokinase-2 Depletion Inhibits Glycolysis and Induces Oxidative Phosphorylation in Hepatocellular Carcinoma and Sensitizes to Metformin. Nat. Commun. 2018, 9, 446. [Google Scholar] [CrossRef]
- Han, Y.; He, M.; Marin, T.; Shen, H.; Wang, W.-T.; Lee, T.-Y.; Hong, H.-C.; Jiang, Z.-L.; Garland, T.; Shyy, J.Y.-J.; et al. Roles of KLF4 and AMPK in the Inhibition of Glycolysis by Pulsatile Shear Stress in Endothelial Cells. Proc. Natl. Acad. Sci. USA 2021, 118, e2103982118. [Google Scholar] [CrossRef]
- Talley, J.T.; Mohiuddin, S.S. Biochemistry, Fatty Acid Oxidation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Lee, M.; Katerelos, M.; Gleich, K.; Galic, S.; Kemp, B.E.; Mount, P.F.; Power, D.A. Phosphorylation of Acetyl-CoA Carboxylase by AMPK Reduces Renal Fibrosis and Is Essential for the Anti-Fibrotic Effect of Metformin. J. Am. Soc. Nephrol. JASN 2018, 29, 2326–2336. [Google Scholar] [CrossRef]
- Arsenault, R.J.; Lee, J.T.; Latham, R.; Carter, B.; Kogut, M.H. Changes in Immune and Metabolic Gut Response in Broilers Fed β-Mannanase in β-Mannan-Containing Diets. Poult. Sci. 2017, 96, 4307–4316. [Google Scholar] [CrossRef]
- Trost, B.; Kindrachuk, J.; Määttänen, P.; Napper, S.; Kusalik, A. PIIKA 2: An Expanded, Web-Based Platform for Analysis of Kinome Microarray Data. PLoS ONE 2013, 8, e80837. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and Recalibrations. Nucleic Acids Res. 2015, 43, D512–D520. [Google Scholar] [CrossRef]
- Venny 2.1. Available online: https://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 17 October 2022).
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kogut, M.H.; Rothwell, L.; Kaiser, P. Differential Regulation of Cytokine Gene Expression by Avian Heterophils During Receptor-Mediated Phagocytosis of Opsonized and Nonopsonized Salmonella Enteritidis. J. Interferon Cytokine Res. 2003, 23, 319–327. [Google Scholar] [CrossRef]
- Ahn, J.; Biswas, D. Influence of Bacteriophage P22 on the Inflammatory Mediator Gene Expression in Chicken Macrophage HD11 Cells Infected with Salmonella Typhimurium. FEMS Microbiol. Lett. 2014, 352, 11–17. [Google Scholar] [CrossRef]
- Boo, S.Y.; Tan, S.W.; Alitheen, N.B.; Ho, C.L.; Omar, A.R.; Yeap, S.K. Identification of Reference Genes in Chicken Intraepithelial Lymphocyte Natural Killer Cells Infected with Very-Virulent Infectious Bursal Disease Virus. Sci. Rep. 2020, 10, 8561. [Google Scholar] [CrossRef]
- Hamal, K.R.; Wideman, R.; Anthony, N.; Erf, G.F. Expression of Inducible Nitric Oxide Synthase in Lungs of Broiler Chickens Following Intravenous Cellulose Microparticle Injection. Poult. Sci. 2008, 87, 636–644. [Google Scholar] [CrossRef] [PubMed]






| Antibiotic | Range of Concentration Tested (μg/mL) | MIC (μg/mL) |
|---|---|---|
| Oxytetracycline | 0.25–32 | >32 |
| Gentamicin | 0.25–32 | 4 |
| Treatment Groups Compared to Infected Control | Number of Changes in Phosphorylation |
|---|---|
| Gentamicin | 540 |
| Grape seed extract | 117 |
| Thyme essential oil | 69 |
| Oxytetracycline | 68 |
| Garlic oil | 66 |
| Capsicum oleoresin | 52 |
| Group | Protein and Phosphorylation Change 1 | Status of the Protein 2 |
|---|---|---|
| Oxytetracycline | PRKAB2 ↑ | Active |
| PGAM1 ↑ | Non active | |
| HK1 ↑ | Active | |
| ACOT12 ↓ | na | |
| PKLR ↓ | Non active (liver) [28] | |
| CAT ↓ | Non active | |
| Thyme essential oil | TPI1 ↓ | Non active [29] |
| ACSL6 ↓ | na | |
| ACSL5 ↓ | na | |
| SUCLA2 ↓ | na | |
| ACAA1 ↑ | na | |
| PGAM1 ↑ | Active | |
| PFKP ↑ | Non active | |
| HK1 ↓ | Active | |
| HK2 ↑ | Active | |
| PFKL ↓ | Non active | |
| Grape seed extract | PFKFB1 ↓ | Non active [30] |
| PYGL ↓ | Active | |
| HK2 ↓ | Active | |
| PGAM1 ↑ | Active | |
| PYGM ↓ | Active | |
| PFKP ↓ | Non active [31] | |
| PKM ↑ | Active | |
| CAT ↓ | Active or non active [32] | |
| PKLR ↓ | Non active (liver) [28] | |
| Garlic oil | PFKP ↓ | Non active [31] |
| PHKA2 ↑ | Active | |
| PGAM1 ↑/↑ | Active/na | |
| PYGL ↑ | Active | |
| Capsicum oleoresin | LDHA ↓ | Active |
| PKLR ↓ | Non active | |
| PRKAA1 ↓ | Non active | |
| CPT1A ↑ | Active | |
| ALDOB ↓ | na |
| Group | Protein and Phosphorylation Change 1 | Status of the Protein 2 |
|---|---|---|
| Oxytetracycline | MAPK14 ↑ | |
| MAPK14 ↑ | MAP2K4 ↑ | Active |
| MAPK14 ↑ | Active | |
| Grape seed extract | CHUK ↑ | Active |
| HSP90AB1 ↑ | Active | |
| HSP90B1 ↑ | Active [34] | |
| MAP2K1 ↓ | Non active [35] | |
| MAP2K4 ↑ | Active | |
| MAP3K7 ↑ | Active | |
| MAPK14 ↑ | Active | |
| RAF1 ↓ | Active | |
| Garlic oil | MAPK1 ↑ | Active |
| RAF1 ↑ | Active | |
| Capsicum oleoresin | MAP2K4 ↑ | Non active |
| MAPK1 ↑ | Active |
| Group (n = 3) | Concentration of Tested Compound | Infection 1 | Experiments 2 |
|---|---|---|---|
| Negative control (CTR –) | None | – | G |
| Positive control (CTR +) | None | + | K/S/G |
| Oxytetracycline (OXY) | 32 μg/mL | + | K/S/G |
| Gentamicin (GEN) | 2 μg/mL | + | K/S/G |
| Thyme essential oil (TEO) | 10 μg/mL | + | K/S/G |
| Grape seed extract (GSE) | 100 μg/mL | + | K/S/G |
| Garlic oil (GAR) | 10 μg/mL | + | K/S/G |
| Capsicum oleoresin (CAP) | 10 μg/mL | + | K/S/G |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovagnoni, G.; Perry, F.; Tugnoli, B.; Piva, A.; Grilli, E.; Arsenault, R.J. A Comparison of the Immunometabolic Effect of Antibiotics and Plant Extracts in a Chicken Macrophage-like Cell Line during a Salmonella Enteritidis Challenge. Antibiotics 2023, 12, 357. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12020357
Giovagnoni G, Perry F, Tugnoli B, Piva A, Grilli E, Arsenault RJ. A Comparison of the Immunometabolic Effect of Antibiotics and Plant Extracts in a Chicken Macrophage-like Cell Line during a Salmonella Enteritidis Challenge. Antibiotics. 2023; 12(2):357. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12020357
Chicago/Turabian StyleGiovagnoni, Giulia, Famatta Perry, Benedetta Tugnoli, Andrea Piva, Ester Grilli, and Ryan J. Arsenault. 2023. "A Comparison of the Immunometabolic Effect of Antibiotics and Plant Extracts in a Chicken Macrophage-like Cell Line during a Salmonella Enteritidis Challenge" Antibiotics 12, no. 2: 357. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12020357