
Techniques for Screening Translation Inhibitors

Abstract
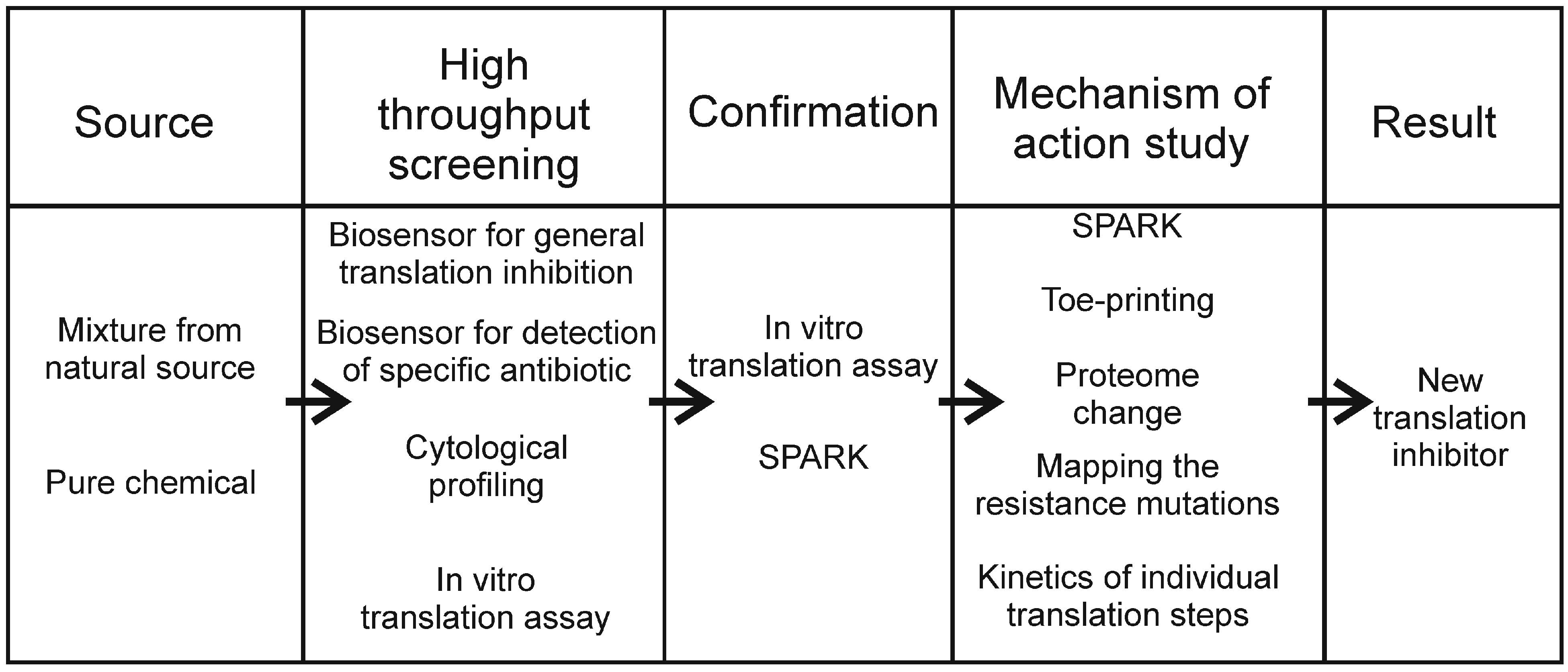
:1. Introduction
2. Biosensor Assays
2.1. Real-Time Measurement of Protein Inhibition Using a Luciferase Assay
2.2. Stress Response Assay
2.3. Tetracycline Detection Assay
2.4. Macrolide Detection: mphA Inducible Assay
2.5. Macrolide Detection: ermC Inducible Assay
2.6. Attenuation-Based Dual Fluorescent Reporter Assay
2.7. Panel of Reporter Strains that Lack Antibiotic Resistance
2.8. Biosensor for Translational Antibiotics Based on Bacillus subtilis yheI Gene
2.9. Transcriptional Sensors Based on Promoters Library
3. In Vitro Methods
3.1. In vitro Protein Synthesis Inhibition Assay
3.2. Toe-Printing of Antibiotic-Stalled Ribosomes
3.3. SPARK-Sensitive Method for Monitoring Peptidyl Transferase Activity
3.4. Antibiotic Binding to a Fluorescently Labeled Ribosome
3.5. The Influence of an Antibiotic on Ribosomal Assembly Measured by Means of Fluorescenly Labeled Ribosomal Subunits
3.6. The Kinetics of Translocation as Sensor of Antibiotic Mechanism of Action
3.7. Biotinylated Biosensors Based on ELISA Technology
4. In Vivo Methods
4.1. Fluorescent Microscopy and Bacterial Cytological Profiling
4.2. Proteomic Signature
4.3. The Mechanism of Resistance as a Sensor of the Mechanism of Action
4.4. Phenotypic Profiling by Raman Spectroscopy
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lewis, K. Platforms for antibiotic discovery. Nat. Rev. Drug Discov. 2013, 12, 371–387. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Lampinen, J.; Virta, M.; Karp, M. Use of controlled luciferase expression to monitor chemicals affecting protein synthesis. Appl. Environ. Microbiol. 1995, 61, 2981–2989. [Google Scholar] [PubMed]
- Galluzzi, L.; Karp, M. Amplified detection of transcriptional and translational inhibitors in bioluminescent Escherichia coli K-12. J. Biomol. Screen. 2003, 8, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, A.A.; Baneyx, F. Stress responses as a tool to detect and characterize the mode of action of antibacterial agents. Appl. Environ. Microbiol. 1999, 65, 5023–5027. [Google Scholar] [PubMed]
- Shapiro, E.; Baneyx, F. Stress-based identification and classification of antibacterial agents: Second-generation Escherichia coli reporter strains and optimization of detection. Antimicrob. Agents Chemother. 2002, 46, 2490–2497. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, E.; Baneyx, F. Stress-activated bioluminescent Escherichia coli sensors for antimicrobial agents detection. J. Biotechnol. 2007, 132, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Kurittu, J.; Karp, M.; Korpela, M. Detection of tetracyclines with luminescent bacterial strains. Lumin J. Biol. Chem. Lumin. 2000, 15, 291–297. [Google Scholar] [CrossRef]
- Mohrle, V.; Stadler, M.; Eberz, G. Biosensor-guided screening for macrolides. Anal. Bioanal. Chem. 2007, 388, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.; Chettiath, T.; Mankin, A.S. Induction of erm(c) expression by noninducing antibiotics. Antimicrob. Agents Chemother. 2008, 52, 866–874. [Google Scholar] [CrossRef] [PubMed]
- Osterman, I.A.; Prokhorova, I.V.; Sysoev, V.O.; Boykova, Y.V.; Efremenkova, O.V.; Svetlov, M.S.; Kolb, V.A.; Bogdanov, A.A.; Sergiev, P.V.; Dontsova, O.A. Attenuation-based dual-fluorescent-protein reporter for screening translation inhibitors. Antimicrob. Agents Chemother. 2012, 56, 1774–1783. [Google Scholar] [CrossRef] [PubMed]
- Melamed, S.; Lalush, C.; Elad, T.; Yagur-Kroll, S.; Belkin, S.; Pedahzur, R. A bacterial reporter panel for the detection and classification of antibiotic substances. Microb. Biotechnol. 2012, 5, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Urban, A.; Eckermann, S.; Fast, B.; Metzger, S.; Gehling, M.; Ziegelbauer, K.; Rubsamen-Waigmann, H.; Freiberg, C. Novel whole-cell antibiotic biosensors for compound discovery. Appl. Environ. Microbiol. 2007, 73, 6436–6443. [Google Scholar] [CrossRef] [PubMed]
- VanBogelen, R.A.; Neidhardt, F.C. Ribosomes as sensors of heat and cold shock in Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 5589–5593. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.; Pollitt, N.S.; Inouye, M. Major cold shock protein of Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, B. Insights into erythromycin action from studies of its activity as inducer of resistance. Antimicrob. Agents Chemother. 1995, 39, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Polikanov, Y.S.; Osterman, I.A.; Szal, T.; Tashlitsky, V.N.; Serebryakova, M.V.; Kusochek, P.; Bulkley, D.; Malanicheva, I.A.; Efimenko, T.A.; Efremenkova, O.V.; et al. Amicoumacin a inhibits translation by stabilizing mRNA interaction with the ribosome. Mol. Cell 2014, 56, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Goh, E.B.; Yim, G.; Tsui, W.; McClure, J.; Surette, M.G.; Davies, J. Transcriptional modulation of bacterial gene expression by subinhibitory concentrations of antibiotics. Proc. Natl. Acad. Sci. USA 2002, 99, 17025–17030. [Google Scholar] [CrossRef] [PubMed]
- Svetlov, M.S.; Kommer, A.; Kolb, V.A.; Spirin, A.S. Effective cotranslational folding of firefly luciferase without chaperones of the Hsp70 family. Protein Sci. Publ. Protein Soc. 2006, 15, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Inoue, A.; Tomari, Y.; Suzuki, T.; Yokogawa, T.; Nishikawa, K.; Ueda, T. Cell-free translation reconstituted with purified components. Nat. Biotechnol. 2001, 19, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Lowell, A.N.; Santoro, N.; Swaney, S.M.; McQuade, T.J.; Schultz, P.J.; Larsen, M.J.; Sherman, D.H. Microscale adaptation of in vitro transcription/translation for high-throughput screening of natural product extract libraries. Chem. Biol. Drug Des. 2015, 86, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Orelle, C.; Carlson, S.; Kaushal, B.; Almutairi, M.M.; Liu, H.; Ochabowicz, A.; Quan, S.; Pham, V.C.; Squires, C.L.; Murphy, B.T.; et al. Tools for characterizing bacterial protein synthesis inhibitors. Antimicrob. Agents Chemother. 2013, 57, 5994–6004. [Google Scholar] [CrossRef] [PubMed]
- Polacek, N.; Swaney, S.; Shinabarger, D.; Mankin, A.S. Spark—A novel method to monitor ribosomal peptidyl transferase activity. Biochemistry 2002, 41, 11602–11610. [Google Scholar] [CrossRef] [PubMed]
- Llano-Sotelo, B.; Hickerson, R.P.; Lancaster, L.; Noller, H.F.; Mankin, A.S. Fluorescently labeled ribosomes as a tool for analyzing antibiotic binding. RNA 2009, 15, 1597–1604. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, R.; Schloemer, R.; Schmidt, S.; Mueller, S.; Heubach, A.; Deuerling, E. Validation of a fluorescence-based screening concept to identify ribosome assembly defects in Escherichia coli. Nucleic Acids Res. 2014, 42, e100. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, W.; Cunha, C.E.; Peske, F.; Konevega, A.L.; Wintermeyer, W.; Rodnina, M.V. GTP hydrolysis by EF-G synchronizes tRNA movement on small and large ribosomal subunits. EMBO J. 2014, 33, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.C.; Link, N.; Fux, C.; Zisch, A.H.; Weber, W.; Fussenegger, M. Broad-spectrum protein biosensors for class-specific detection of antibiotics. Biotechnol. Bioeng. 2005, 89, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Nonejuie, P.; Burkart, M.; Pogliano, K.; Pogliano, J. Bacterial cytological profiling rapidly identifies the cellular pathways targeted by antibacterial molecules. Proc. Natl. Acad. Sci. USA 2013, 110, 16169–16174. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, M.; Bandow, J.E. Proteomic signatures in antibiotic research. Proteomics 2011, 11, 3256–3268. [Google Scholar] [CrossRef] [PubMed]
- Bandow, J.E.; Brotz, H.; Leichert, L.I.; Labischinski, H.; Hecker, M. Proteomic approach to understanding antibiotic action. Antimicrob. Agents Chemother. 2003, 47, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Vazquez-Laslop, N.; Mankin, A.S. Selective protein synthesis by ribosomes with a drug-obstructed exit tunnel. Cell 2012, 151, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Osterman, I.A.; Ustinov, A.V.; Evdokimov, D.V.; Korshun, V.A.; Sergiev, P.V.; Serebryakova, M.V.; Demina, I.A.; Galyamina, M.A.; Govorun, V.M.; Dontsova, O.A. A nascent proteome study combining click chemistry with 2DE. Proteomics 2013, 13, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Athamneh, A.I.; Alajlouni, R.A.; Wallace, R.S.; Seleem, M.N.; Senger, R.S. Phenotypic profiling of antibiotic response signatures in Escherichia coli using Raman spectroscopy. Antimicrob. Agents Chemother. 2014, 58, 1302–1314. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type | Year, Reference | Signal | Assay Time and Conditions | Description | Tested Molecules | Notes |
|---|---|---|---|---|---|---|
| Biosensor for translation inhibitors | 1995, [3] | Luciferase | 30–45 min (liquid assay) | Plasmid reporter, E. coli pL (bacteriophage lambda leftward promoter)–luxAB (Photorhabdus luminescens) pL–luxAB (Vibrio harveyi) pL–lucGR (Pyrophorus plagiophthalamus) | Tetracycline P, Sodium fluoride FP, Chloramphenicol P, Nicotine FP, Cycloheximide N, Mercuric chloride FP, Polymixin B FP, Phenol FP | Direct measuring of translation |
| Biosensor for translation inhibitors | 2003, [4] | Luciferase | Up to 3 h (liquid assay) | Genome reporter, E. coli ptetA (tetracycline inducible)–luxCDABE (P. luminescens) | Rifampicin FP, Imipenem FP, Chloramphenicol P, Sulfadiazine FP, Ciprofloxacin FP, Cefotaxime FP, Erythromycin P, Spiramycin P, Trimethoprim FP | Direct measuring of translation |
| Biosensor for translation inhibitors | 1999, [5] | Beta-galactosidase | Up to 3 h (liquid assay) Overnight (solid assay) | Genome reporter, E. coli or E. coli:ΔtolC [6] , pcspA (promoter-utr-start of codon region of cspA gene)–lacZ or luc (Photinus pyralis) [7] pibp (promoter-utr-start of codon region of ibp gene)–lacZ or luc (Photinus pyralis) [7] | Tetracycline P, Chloramphenicol P, Streptomycin P, Neomicin P, Polymyxin B FP, Nalidixic acid N, Carbenicillin N | Stress response promoters |
| Biosensor for tetracyclines | 2000, [8] | Luciferase | 90 min (liquid assay) | Genome reporters, E. coli, ptetA-luc (Photinus pyralis) ptetA-luxCDABE (Photorhabdus luminescens) | Tetracyclines P | Tetracycline inducible promoter of tetA |
| Biosensor for macrolides | 2007, [9] | Luciferase | 2 h (liquid assay) 1–2 h (solid assay) | Genome reporter, E. coli pmphR (A) (mphA promoter)–luxCDABE (Vibrio fischeri) | Erythromycin P, Methymycin P, Azithromycin P, Clarithromycin P, Oleandomycin P, Tetracycline N, Chloramphenicol N, Gentamycin N, Cefaclor N, Novobiocin N, Fosfomycin N, Vancomycin N | Macrolide inducible promoter of mphA |
| Biosensor for macrolides | 2008, [10] | Beta-galactosidase | Overnight (solid assay) | Plasmid reporter, E. coli, Ptac-ermC (L)–lacZ(α). | Erythromycin P, Clarithromycin P, Azithromycin FN, Clarithromycin P, Josamycin N, Telithromycin P, Spiramycin N, Tylosin N, Roxithromycin P, Chloramphenicol N | Macrolide inducible expression of ermC |
| Biosensor for ribosome stalling inhibitors | 2012, [11] | Cerulean fluorescent protein | Overnight (solid assay) Overnight (liquid assay) | Plasmid reporter, E. coli or E. coli:ΔtolC, pT5–trpL–2Ala–CER. | Erythromycin P, Clarithromycin P, Roxithromycin P, Oleandomycin P, Tylosin P, Azithromycin P, Chloramphenicol P, Linezolid P, Lincomycin P, Sulfanilamide N, Polymyxin N, Rifampicin N, Kanamycin N, Tetracycline FN, Streptomycin N, Vancomycin N, Clindamycin FN, Nalidixic acid N, Phosphomycin N, Levofloxacin N, Norfloxacin N, Tobramycin N, Neomycin N | Ribosome stalling induces expression of the signal gene |
| Biosensor for translation inhibitors | 2012, [12] | Luciferase | 10 h (liquid assay) | Plasmid reporter, E. coli psoxS (promoter of soxS gene)–luxCDABE (Photorhabdus luminescens) | Tetracycline P, Oxytetracycline P, Chloramphenicol P, Sulfamethoxazole N, Sulfadimethoxine N, Ampicillin N, Amoxicillin N, Nalidixic Acid N, Rifampicin N, Puromycin FN, Colistin N | |
| Biosensor for translation inhibitors | 2007, [13] | Luciferase | 4 h (liquid assay) | Plasmid reporter, Bacillus subtilis pyheI (promoter of yheI gene)–luc (Photinus pyralis) | Moxifloxacin N, Ciprofloxacin N, Trovafloxacin N, Nalidixic acid N, Novobiocin N, Trimethoprim N, Azaserine N, Mitomycin C N, Streptovaricin N, Rifampicin N, Linezolid P, Doxycycline P, Fusidic acid P, Chloramphenicol P, Gentamicin FN, Kanamycin FN, Puromycin FN, Actinonin FN, Cefoxitin, Oxacillin N, Mersacidin N, Ramoplanin N, Vancomycin N, Tunicamycin N, Polymyxin B N, Moiramide B N, Cerulenin N, Triclosan N, Netropsin N, Ethidium bromide N, Actinomycin D N, Monensin N, N-ethyl maleimide N |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osterman, I.A.; Bogdanov, A.A.; Dontsova, O.A.; Sergiev, P.V. Techniques for Screening Translation Inhibitors. Antibiotics 2016, 5, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics5030022
Osterman IA, Bogdanov AA, Dontsova OA, Sergiev PV. Techniques for Screening Translation Inhibitors. Antibiotics. 2016; 5(3):22. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics5030022
Chicago/Turabian StyleOsterman, Ilya A., Alexey A. Bogdanov, Olga A. Dontsova, and Petr V. Sergiev. 2016. "Techniques for Screening Translation Inhibitors" Antibiotics 5, no. 3: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics5030022