Specificity in the Susceptibilities of Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus Clinical Isolates to Six Metal Antimicrobials



Abstract
:1. Introduction
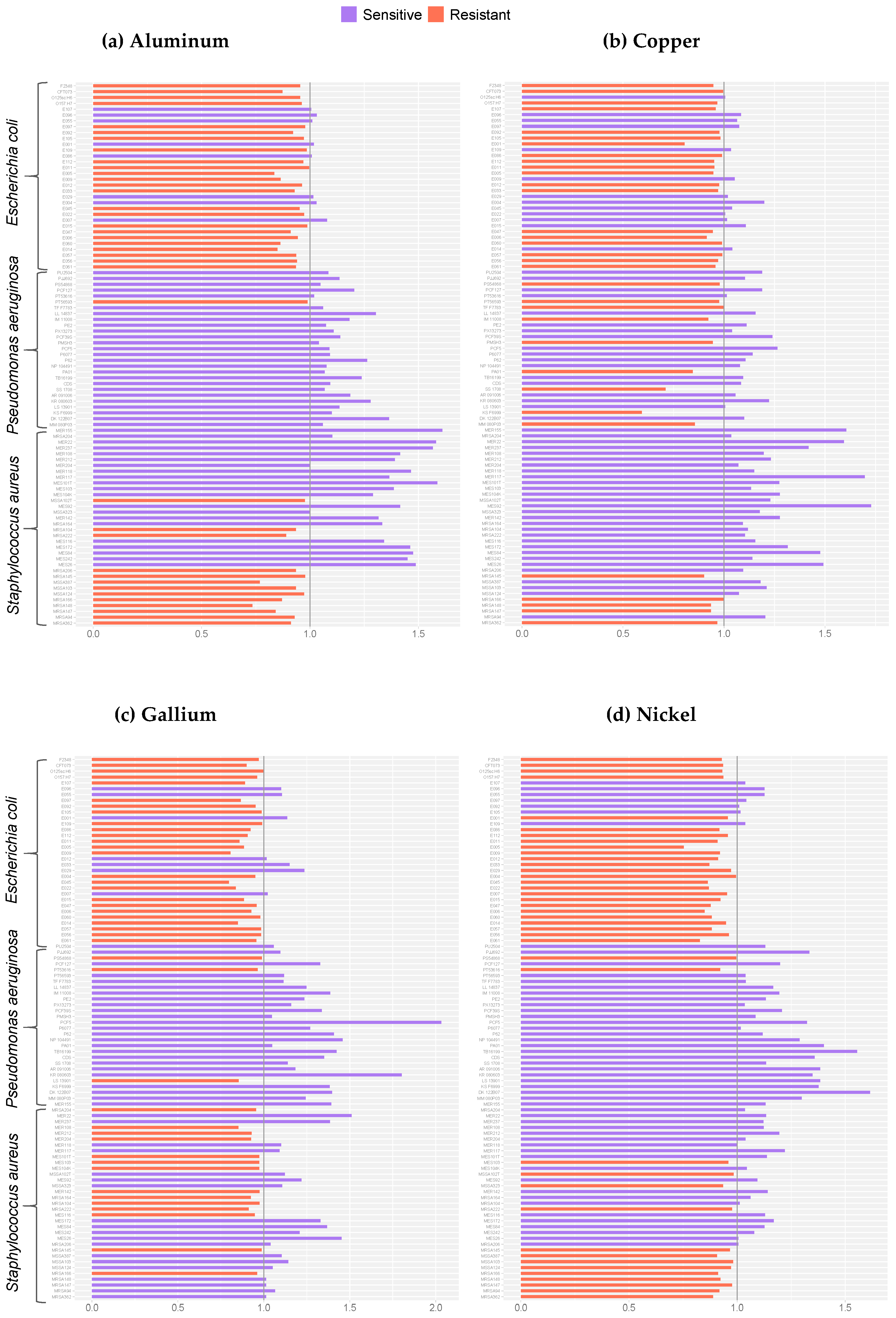
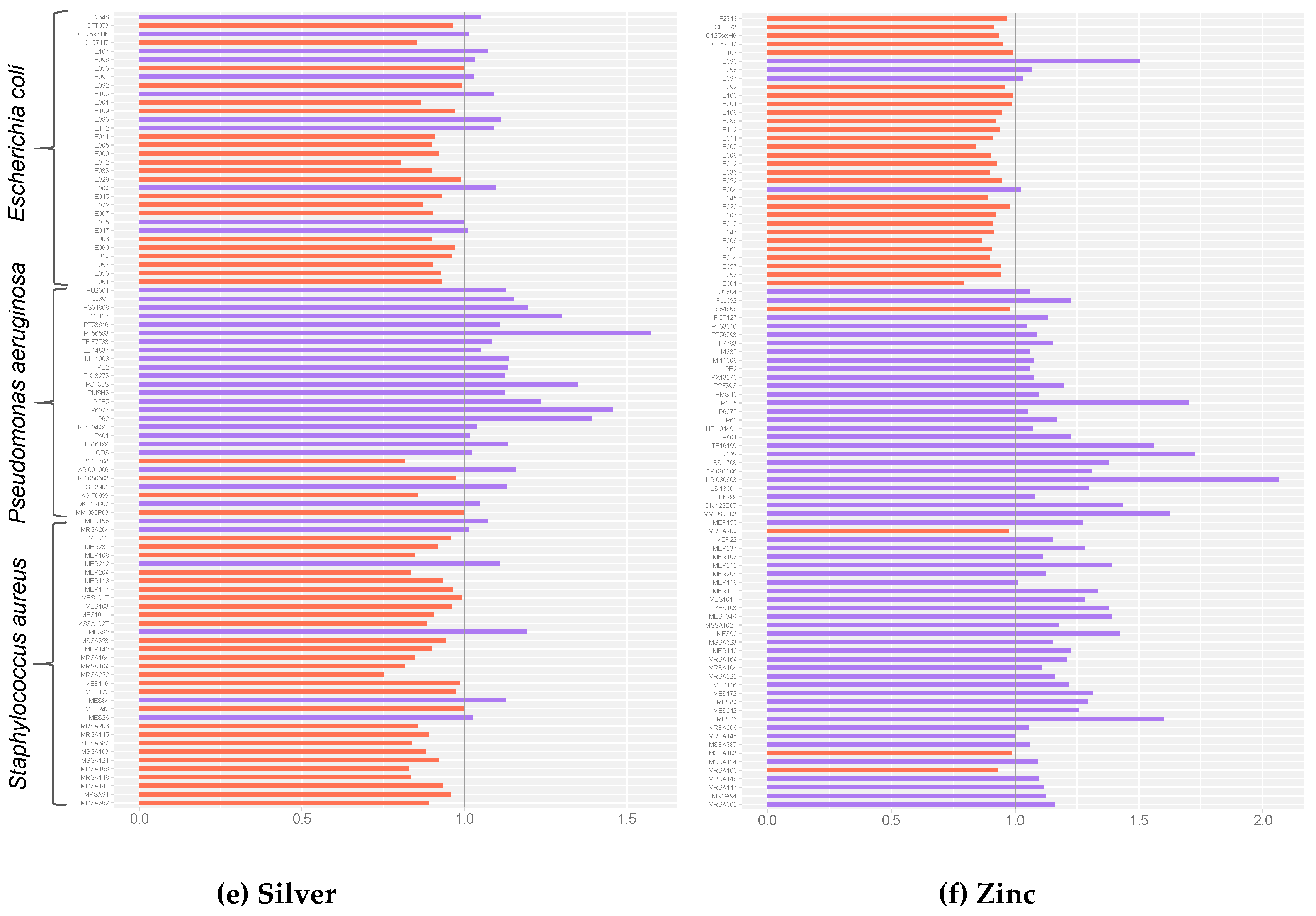
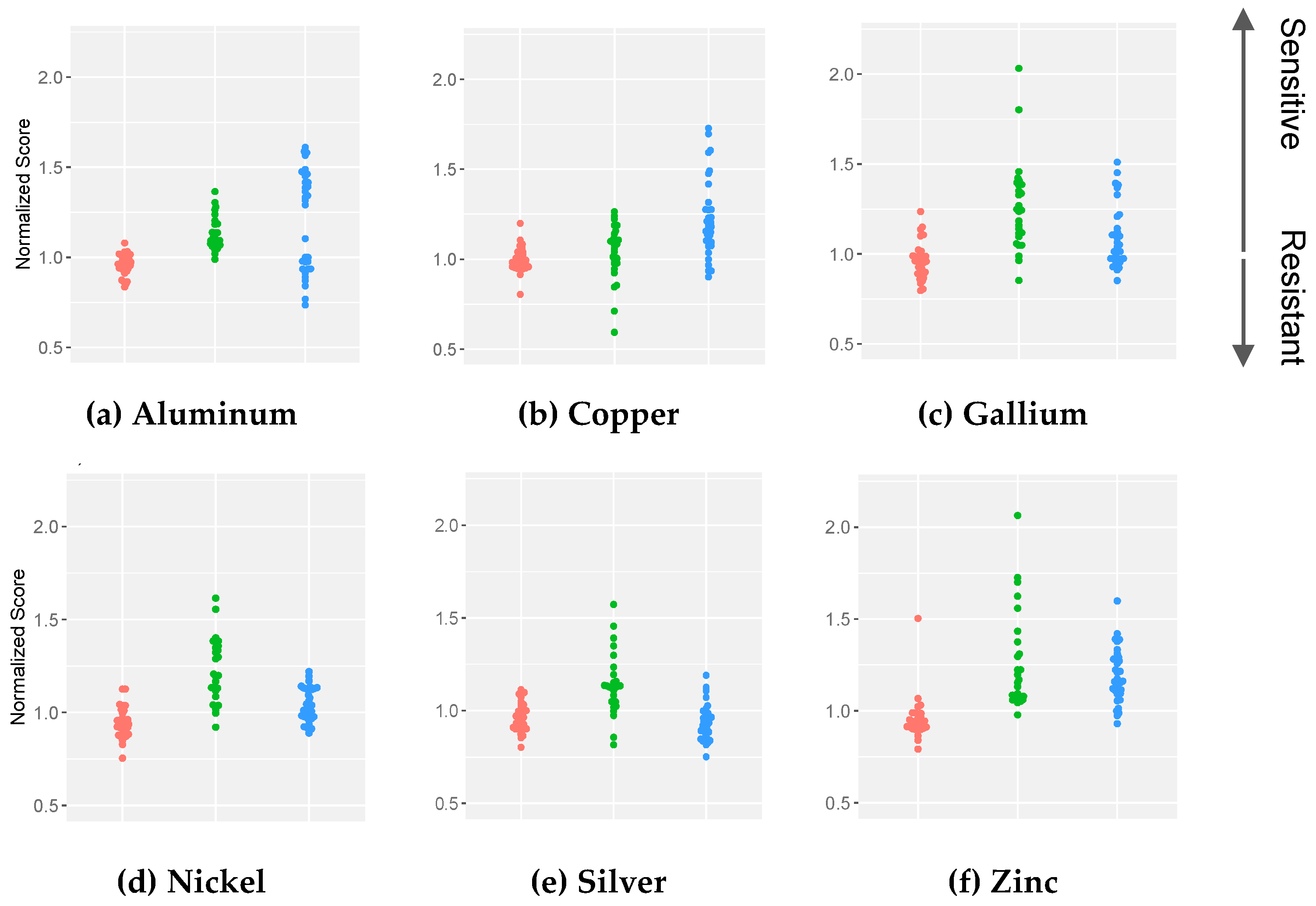
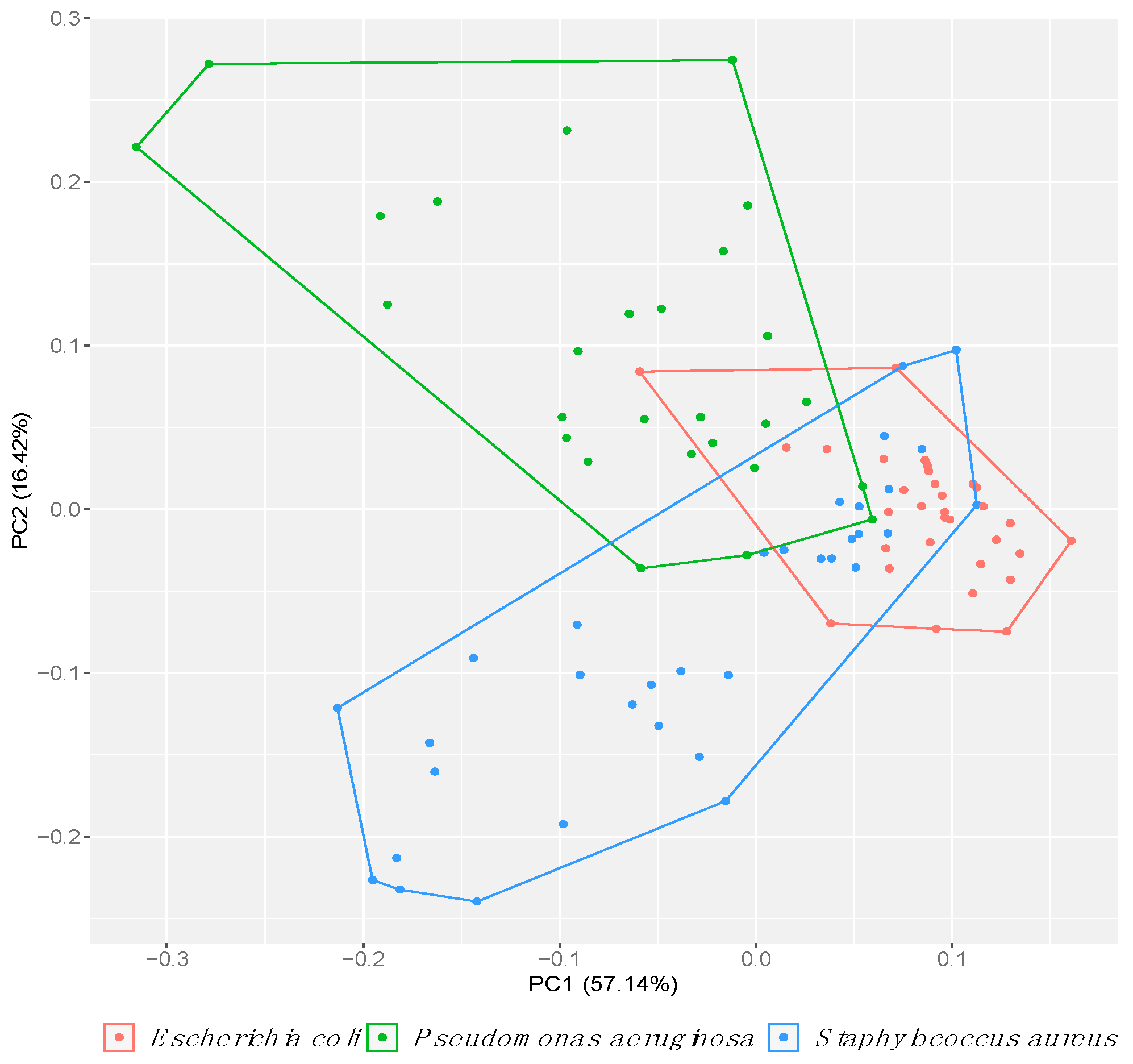
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Storage
4.2. Determination of the Effective Metal Concentrations and Metal Storage
4.3. Bacterial Growth and the Agar Disk-Diffusion Method
4.4. Normalization and Statistical Analyses
- Biological trial 1 diameters for zinc (mm): 26, 27 mean: 26.5
- Biological trial 2 diameters for zinc (mm): 26, 27 mean: 26.5
- Biological trial 3 diameters for zinc (mm): 25, 26 mean: 25.5
- Biological trial 1 diameters for zinc (mm): 25, 25 mean: 25
- Biological trial 2 diameters for zinc (mm): 25, 25 mean: 25
- Biological trial 3 diameters for zinc (mm): 22. 23 mean: 22.5
- Biological trial 1, zinc: 25/26.5 = 0.943
- Biological trial 2, zinc: 25/26.5 = 0.943
- Biological trial 3, zinc: 22.5/25.5 = 0.882
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal | Escherichia coli ATCC 25922 | P. aeruginosa ATCC 27853 | S. aureus ATCC 25923 |
|---|---|---|---|
| Aluminum [Al2(SO4)3·H2O] | 250 mM | 1.95 mM | > 250 mM |
| Copper (CuSO4) | 12.5 mM | 6.25 mM | 12.5 mM |
| Gallium [Ga(NO3)3·H2O] | 31.25 mM | 15.63 mM | 15.63 mM |
| Nickel (NiSO4) | >625 mM | >625 mM | >625 mM |
| Silver (AgNO3) | >0.50 mM 1 | >0.50 mM 1 | >0.50 mM 1 |
| Zinc (ZnSO4·7H2O) | >650 mM | > 375 mM 1 | 23.44 mM |
References
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef]
- Neu, H.C. The Crisis in Antibiotic Resistance. Science 1993, 257, 1064–1073. [Google Scholar] [CrossRef]
- Park, A.J.; Okhovat, J.P.; Kim, J. Antimicrobial Peptides. Clin. Basic Immunodermatol. 2017. [Google Scholar] [CrossRef]
- Kenawy, E.R.; Worley, S.D.; Broughton, R. The Chemistry and Applications of Antimicrobial Polymers: A State-of-the-Art Review. Biomacromolecules 2007, 8, 1359–1384. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Brown, E.D.; Wildenhain, J.; Farha, M.A.; Wright, G.D.; Coombes, B.K.; Ejim, L.; Falconer, S.B. Combinations of Antibiotics and Nonantibiotic Drugs Enhance Antimicrobial Efficacy. Nat. Chem. Biol. 2011, 7, 348–350. [Google Scholar]
- Turner, R.J. Metal-Based Antimicrobial Strategies. Microb. Biotechnol. 2017, 10, 1062–1065. [Google Scholar] [CrossRef]
- Dupont, C.L.; Yang, S.; Palenik, B.; Bourne, P.E. Modern Proteomes Contain Putative Imprints of Ancient Shifts in Trace Metal Geochemistry. Proc. Natl. Acad. Sci. USA 2006, 103, 17822–17827. [Google Scholar] [CrossRef] [PubMed]
- Waldron, K.J.; Robinson, N.J. How Do Bacterial Cells Ensure That Metalloproteins Get the Correct Metal? Nat. Rev. Microbiol. 2009, 7, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I.; Rosato, A. A Hint to Search for Metalloproteins in Gene Banks. Bioinformatics 2004, 20, 1373–1380. [Google Scholar] [CrossRef]
- Nies, D.H. Microbial Heavy-Metal Resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Ceri, H.; Stremick, C.A.; Turner, R.J. Biofilm Susceptibility to Metal Toxicity. Environ. Microbiol. 2004, 6, 1220–1227. [Google Scholar] [CrossRef]
- Kaneko, Y.; Thoendel, M.; Olakanmi, O.; Britigan, B.E.; Singh, P.K. The Transition Metal Gallium Disrupts Pseudomonas aeruginosa Iron Metabolism and Has Antimicrobial and Antibiofilm Activity. J. Clin. Invest. 2007, 117, 877–888. [Google Scholar] [CrossRef]
- Gugala, N.; Lemire, J.A.; Turner, R.J. The Efficacy of Different Anti-Microbial Metals at Preventing the Formation of and Eradicating Bacterial Biofilms of Pathogenic Indicator Strains. J. Antibiot. 2017, 70, 775–780. [Google Scholar] [CrossRef]
- Harrison, J.J.; Turner, R.J.; Ceri, H. High-Throughput Metal Susceptibility Testing of Microbial Biofilms. BMC Microbiol. 2005, 5, 1–11. [Google Scholar] [CrossRef]
- Khan, S.T.; Musarrat, J.; Al-Khedhairy, A.A. Countering Drug Resistance, Infectious Diseases and Sepsis Using Metal and Metal Oxides Nanoparticles: Current Status. Colloids Surf. B Biointerfaces 2016, 146, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.B.; Lam, K.; Burrell, R.E. Wound Management in an Era of Increasing Bacterial Antibiotic Resistance: A Role for Topical Silver Treatment. Am. J. Infect. Control 1998, 26, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Mikolay, A.; Huggett, S.; Tikana, L.; Grass, G.; Braun, J.; Nies, D.H. Survival of Bacteria on Metallic Copper Surfaces in a Hospital Trial. Appl. Microbiol. Biotechnol. 2010, 87, 1875–1879. [Google Scholar] [CrossRef] [PubMed]
- Palza, H.; Nuñez, M.; Bastías, R.; Delgado, K. In Situ Antimicrobial Behavior of Materials with Copper-Based Additives in a Hospital Environment. Int. J. Antimicrob. Agents 2018, 51, 912–917. [Google Scholar] [CrossRef]
- Shastri, J.P.; Rupani, M.G.; Jain, R.L. Antimicrobial Activity of Nanosilver-Coated Socks Fabrics against Foot Pathogens. J. Text. Inst. 2012, 103, 1234–1243. [Google Scholar] [CrossRef]
- Atiyeh, B.S.; Costagliola, M.; Hayek, S.N.; Dibo, S.A. Effect of Silver on Burn Wound Infection Control and Healing: Review of the Literature. Burns 2007, 33, 139–148. [Google Scholar] [CrossRef]
- Borkow, G.; Zatcoff, R.C.; Gabbay, J. Reducing the Risk of Skin Pathologies in Diabetics by Using Copper Impregnated Socks. Med. Hypotheses 2009, 73, 883–886. [Google Scholar] [CrossRef]
- Li, Y.; Leung, P.; Yao, L.; Song, Q.W.; Newton, E. Antimicrobial Effect of Surgical Masks Coated with Nanoparticles. J. Hosp. Infect. 2006, 62, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Borkow, G.; Zhou, S.S.; Page, T.; Gabbay, J. A Novel Anti-Influenza Copper Oxide Containing Respiratory Face Mask. PLoS ONE 2010, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ghazvini, K.; Barati, S.; Ahrari, F.; Eslami, N.; Rajabi, O. The Antimicrobial Sensitivity of Streptococcus mutans and Streptococcus sangius to Colloidal Solutions of Different Nanoparticles Applied as Mouthwashes. Dent. Res. J. (Isfahan). 2015, 12, 44–49. [Google Scholar]
- Liu, Y.; He, L.; Mustapha, A.; Li, H.; Hu, Z.Q.; Lin, M. Antibacterial Activities of Zinc Oxide Nanoparticles against Escherichia coli O157:H7. J. Appl. Microbiol. 2009, 107, 1193–1201. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Sierra, J.F.; Ruiz, F.; Cruz Pena, D.C.; Martínez-Gutiérrez, F.; Martínez, A.E.; de Jesús Pozos Guillén, A.; Tapia-Pérez, H.; Martínez Castañón, G. The Antimicrobial Sensitivity of Streptococcus mutans to Nanoparticles of Silver, Zinc Oxide and Gold. Nanomed. Nanotechnol. Biol. Med. 2008, 4, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Radzig, M.A.; Nadtochenko, V.A.; Koksharova, O.A.; Kiwi, J.; Lipasova, V.A.; Khmel, I.A. Antibacterial Effects of Silver Nanoparticles on Gram-Negative Bacteria: Influence on the Growth and Biofilms Formation, Mechanisms of Action. Colloids Surfaces B Biointerfaces 2013, 102, 300–306. [Google Scholar] [CrossRef]
- Wahid, F.; Zhong, C.; Wang, H.; Hu, X.; Chu, L. Recent Advances in Antimicrobial Hydrogels Containing Metal Ions and Metals/Metal Oxide Nanoparticles. Polymers (Basel). 2017, 9, 636. [Google Scholar] [CrossRef]
- Palza, H. Antimicrobial Polymers with Metal Nanoparticles. Int. J. Mol. Sci. 2015, 16, 2099–2116. [Google Scholar] [CrossRef]
- Li, P.; Li, J.; Wu, C.; Wu, Q.; Li, J. Synergistic Antibacterial Effects of Beta-Lactam Antibiotic Combined with Silver Nanoparticles. Nanotechnology 2005, 16, 1912–1917. [Google Scholar] [CrossRef]
- Fayaz, A.M.; Balaji, K.; Girilal, M.; Yadav, R.; Kalaichelvan, P.T.; Venketesan, R. Biogenic Synthesis of Silver Nanoparticles and Their Synergistic Effect with Antibiotics: A Study against Gram-positive and Gram-negative Bacteria. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 103–109. [Google Scholar] [CrossRef]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial Activity of Metals: Mechanisms, Molecular Targets and Applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in Vitro Evaluating Antimicrobial Activity: A Review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Azam, A. Antimicrobial Activity of Metal Oxide Nanoparticles against Gram-positive and Gram-negative Bacteria: A Comparative Study. Int. J. Nanomedicine 2012, 7, 6003–6009. [Google Scholar] [CrossRef]
- Ruparelia, J.P.; Kumar, A.; Duttagupta, S.P. Strain Specificity in Antimicrobial Activity of Silver and Copper Nanoparticles. Acta Biomater. 2008, 4, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Vance, M.E.; Kuiken, T.; Vejerano, E.P.; McGinnis, S.P.; Hochella, M.F.; Hull, D.R. Nanotechnology in the Real World: Redeveloping the Nanomaterial Consumer Products Inventory. Beilstein J. Nanotechnol. 2015, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Ceri, H.; Turner, R.J. Multimetal Resistance and Tolerance in Microbial Biofilms. Nat. Rev. Microbiol. 2007, 5, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J. Plasmid-Determined Resistance to Antimicrobial Drugs and Toxic Metal Ions in Bacteria. Microbiol. Rev. 1983, 47, 361–409. [Google Scholar] [PubMed]
- Teitzel, G.M.; Parsek, M.R. Heavy Metal Resistance of Biofilm and Planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. Resistance of Planktonic and Biofilm-Grown Burkholderia cepacia Complex Isolates to the Transition Metal Gallium. J. Antimicrob. Chemother. 2008, 61, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Efflux-Mediated Heavy Metal Resistance in Prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Hobman, J.L.; Crossman, L.C. Bacterial Antimicrobial Metal Ion Resistance. J. Med. Microbiol. 2015, 64, 471–497. [Google Scholar] [CrossRef] [PubMed]
- Harrison, J.J.; Rabiei, M.; Turner, R.J.; Badry, E.A.; Sproule, K.M.; Ceri, H. Metal Resistance in Candida Biofilms. FEMS Microbiol. Ecol. 2006, 55, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Hur, H. Metagenomic analysis reveals the prevalence and persistence of antibiotic-and heavy metal-resistance genes in wastewater treatment plant. J. Microbiol. 2018, 56, 408–415. [Google Scholar]
- Li, A.-D.; Li, L.-G.; Zhang, T. Exploring Antibiotic Resistance Genes and Metal Resistance Genes in Plasmid Metagenomes from Wastewater Treatment Plants. Front. Microbiol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-Selection of Antibiotic and Metal Resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.S.; Peltier, G.L.; Stepanauskas, R.; McArthur, J.V. Bacterial Tolerances to Metals and Antibiotics in Metal-Contaminated and Reference Streams. FEMS Microbiol. Ecol. 2006, 58, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Pal, C.; Asiani, K.; Arya, S.; Rensing, C.; Stekel, D.J.; Larsson, D.G.J.; Hobman, J.L. Metal Resistance and Its Association with Antibiotic Resistance. In Advances in Microbial Physiology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 70, pp. 261–313. [Google Scholar]
- Um, M.M.; Brugère, H.; Kérourédan, M.; Oswald, E.; Bibbal, D. Antimicrobial Resistance Profiles of Enterohemorrhagic and Enteropathogenic Escherichia coli of Serotypes O157:H7, O26:H11, O103:H2, O111:H8, O145:H28 Compared to Escherichia coli Isolated from the Same Adult Cattle. Microb. Drug Resist. 2018, 24, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular Mechanisms of Antibiotic Resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy Metal Driven Co-Selection of Antibiotic Resistance in Soil and Water Bodies Impacted by Agriculture and Aquaculture. Front. Microbiol. 2012, 3, 1–10. [Google Scholar] [CrossRef]
- Chen, J.; Cen, T.; He, M.; Gu, A.Z.; Zhang, Y.; Li, D.; Li, X. Sub-Inhibitory Concentrations of Heavy Metals Facilitate the Horizontal Transfer of Plasmid-Mediated Antibiotic Resistance Genes in Water Environment. Environ. Pollut. 2018, 237, 74–82. [Google Scholar]
- Lemire, J.A.; Kalan, L.; Gugala, N.; Bradu, A.; Turner, R.J. Silver Oxynitrate–an Efficacious Compound for the Prevention and Eradication of Dual-Species Biofilms. Biofouling 2017, 33, 460–469. [Google Scholar] [CrossRef]
- Burmølle, M.; Webb, J.S.; Rao, D.; Hansen, L.H.; Sørensen, S.J.; Kjelleberg, S. Enhanced Biofilm Formation and Increased Resistance to Antimicrobial Agents and Bacterial Invasion Are Caused by Synergistic Interactions in Multispecies Biofilms. Appl. Environ. Microbiol. 2006, 72, 3916–3923. [Google Scholar] [CrossRef]
- Sohlenkamp, C.; Geiger, O. Bacterial Membrane Lipids: Diversity in Structures and Pathways. FEMS Microbiol. Rev. 2015, 40, 133–159. [Google Scholar] [CrossRef]
- Politano, A.D.; Campbell, K.T.; Rosenberger, L.H.; Sawyer, R.G. Use of Silver in the Prevention and Treatment of Infections: Silver Review. Surg. Infect. (Larchmt). 2013, 14, 8–20. [Google Scholar] [CrossRef]
- Alexander, J.W. History of the Medical Use of Silver. Surg. Infect. (Larchmt). 2009, 10, 289–292. [Google Scholar] [CrossRef]
- Melaiye, A.; Youngs, W.J. Silver and Its Application as an Antimicrobial Agent. Expert Opin. Ther. Pat. 2005, 15, 125–130. [Google Scholar] [CrossRef]
- O’Gorman, J.; Humphreys, H. Application of Copper to Prevent and Control Infection. Where Are We Now? J. Hosp. Infect. 2012, 81, 217–223. [Google Scholar] [CrossRef]
- Yasuyuki, M.; Kunihiro, K.; Kurissery, S.; Kanavillil, N.; Sato, Y.; Kikuchi, Y. Antibacterial Properties of Nine Pure Metals: A Laboratory Study Using Staphylococcus aureus and Escherichia coli. Biofouling 2010, 26, 851–858. [Google Scholar] [CrossRef]
- Li, Y.; Zamble, D.B. Nickel Homeostasis and Nickel Regulation: An Overview. Chem. Rev. 2009, 109, 4617–4643. [Google Scholar] [CrossRef]
- Irving, B.H.; Williams, R.J.P. The Stability of Transition-Metal Complexes. J. Chem. Soc. 1953, 0, 3192–3210. [Google Scholar] [CrossRef]
- Patel, J.B.; Cockerill, F.R.; Bradford, P.A.; Eliopoulos, G.M.; Hindler, J.A.; Jenkins, S.G.; Lewis, J.S.; Limbago, B. M02-A12: Performance Standards for Antimicrobial Disk Susceptibility Tests; Approved Standard—Twelfth Edition; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2015. [Google Scholar]
- Werthén, M.; Henriksson, L.; Jensen, P.Ø.; Sternberg, C.; Givskov, M.; Bjarnsholt, T. An in Vitro Model of Bacterial Infections in Wounds and Other Soft Tissues. APMIS 2010, 118, 156–164. [Google Scholar] [CrossRef]
- Lemire, J.A.; Kalan, L.; Bradu, A.; Turner, R.J. Silver Oxynitrate, an Unexplored Silver Compound with Antimicrobial and Antibiofilm Activity. Antimicrob. Agents Chemother. 2015, 59, 4031–4039. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gugala, N.; Vu, D.; Parkins, M.D.; Turner, R.J. Specificity in the Susceptibilities of Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus Clinical Isolates to Six Metal Antimicrobials. Antibiotics 2019, 8, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8020051
Gugala N, Vu D, Parkins MD, Turner RJ. Specificity in the Susceptibilities of Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus Clinical Isolates to Six Metal Antimicrobials. Antibiotics. 2019; 8(2):51. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8020051
Chicago/Turabian StyleGugala, Natalie, Dennis Vu, Michael D. Parkins, and Raymond J. Turner. 2019. "Specificity in the Susceptibilities of Escherichia coli, Pseudomonas aeruginosa and Staphylococcus aureus Clinical Isolates to Six Metal Antimicrobials" Antibiotics 8, no. 2: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8020051