Characterization of the Metabolic Response of Streptomyces clavuligerus to Shear Stress in Stirred Tanks and Single-Use 2D Rocking Motion Bioreactors for Clavulanic Acid Production

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
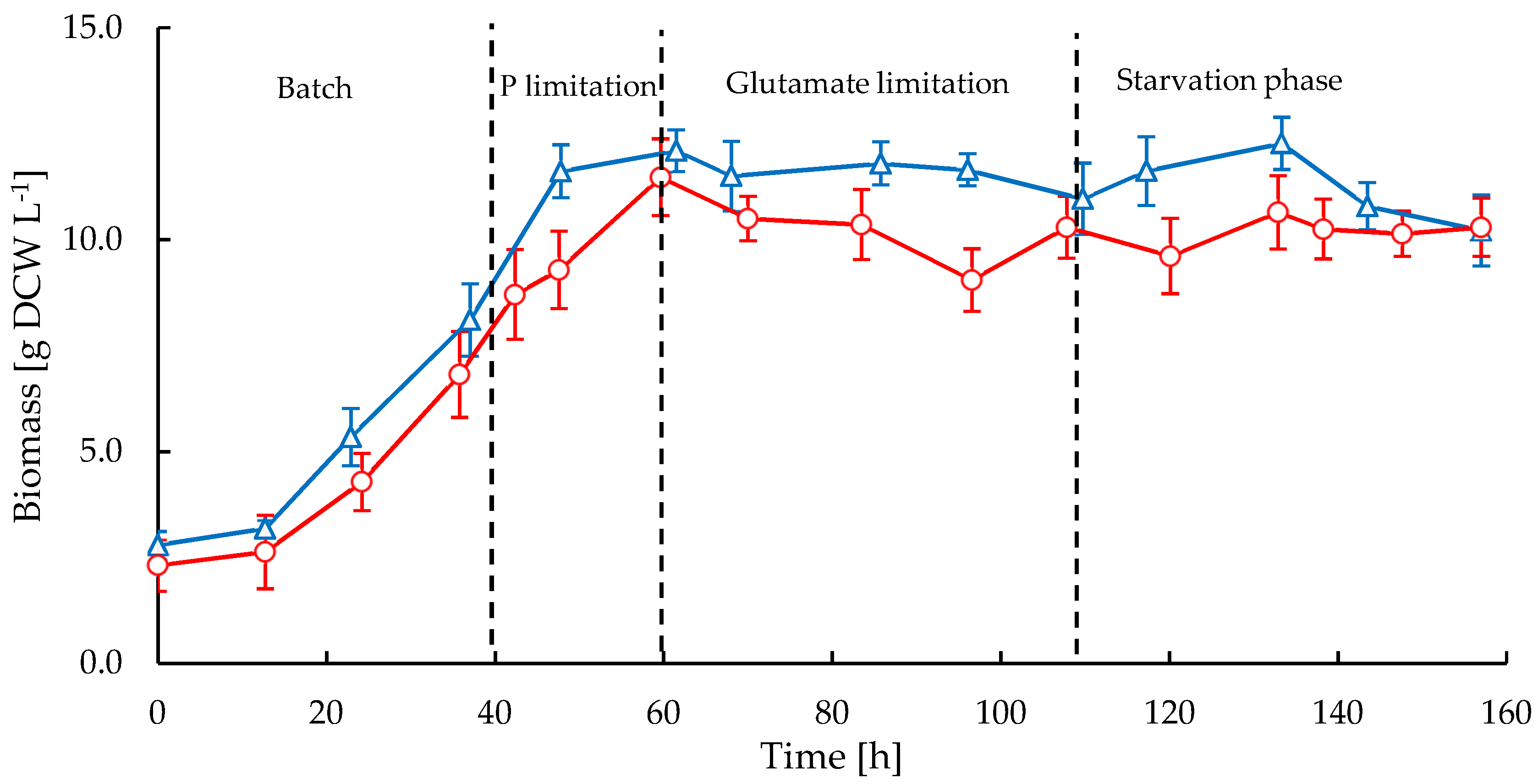
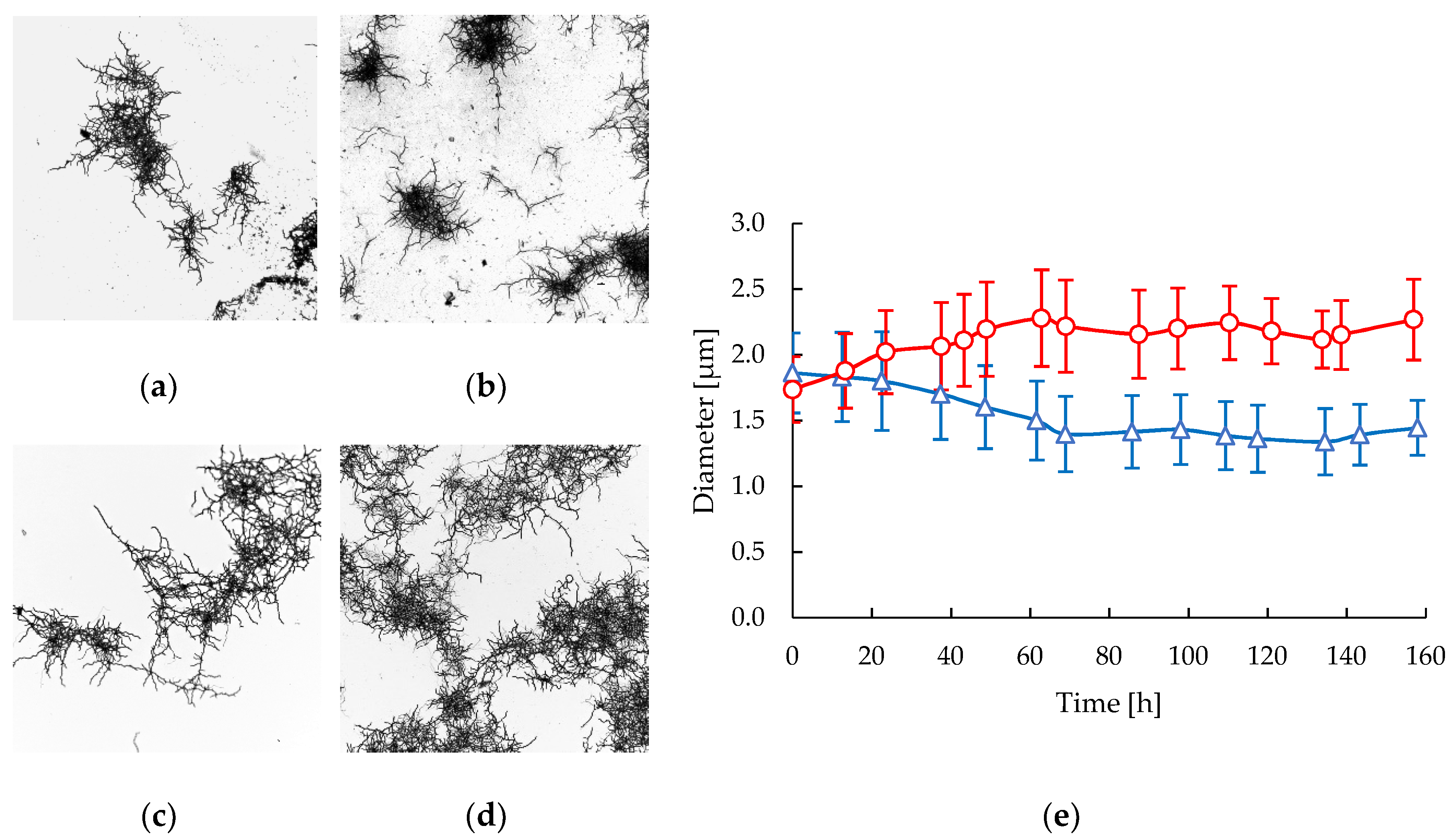
2.1. Biomass and Morphology
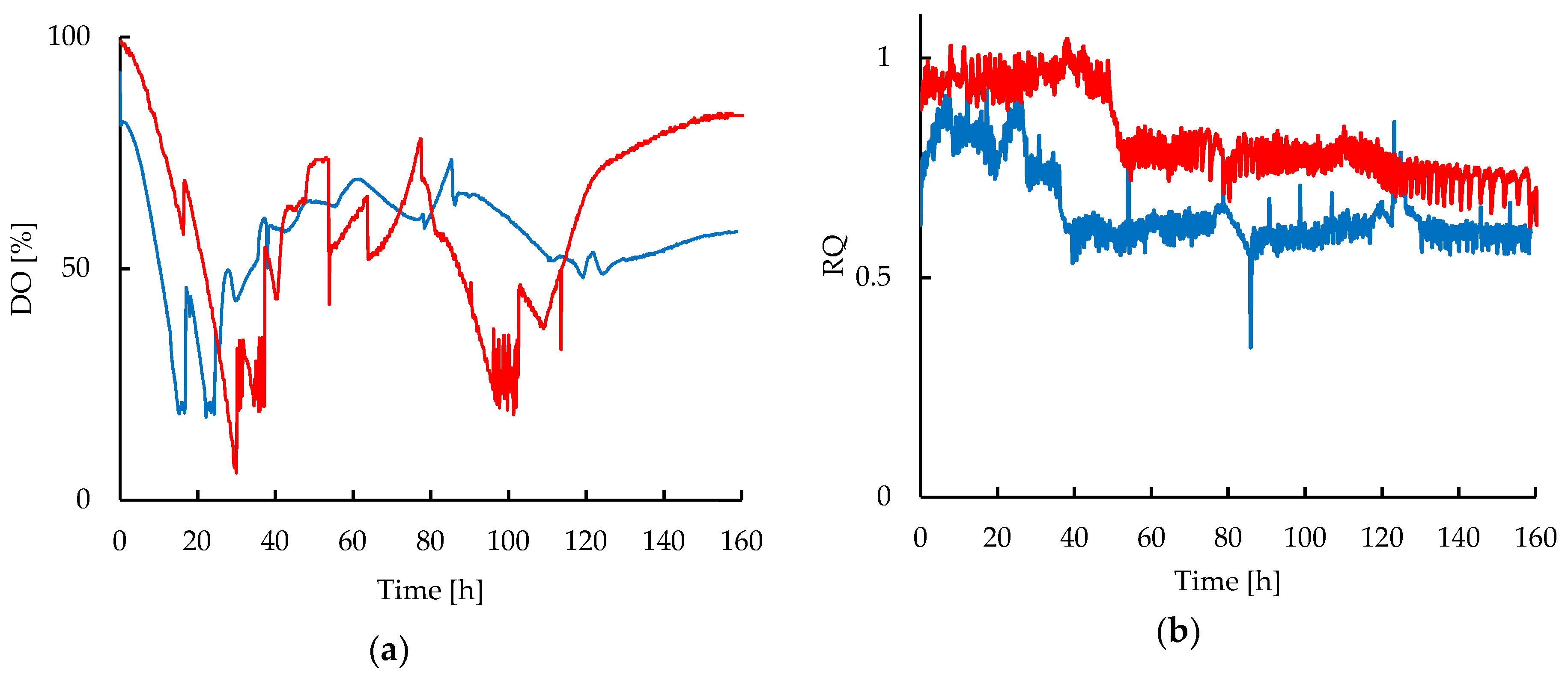
2.2. Shear Stress and Oxygen Transfer
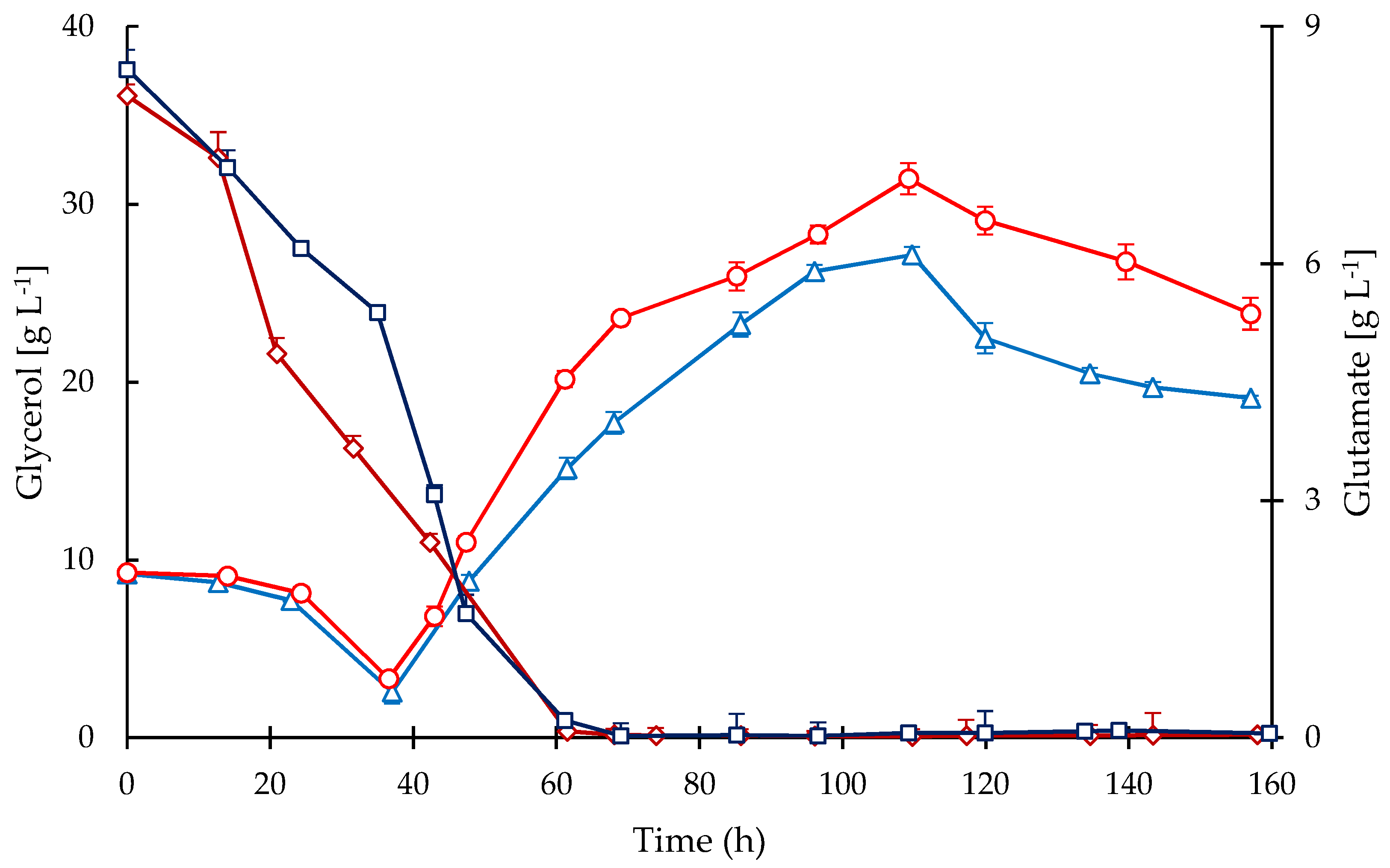
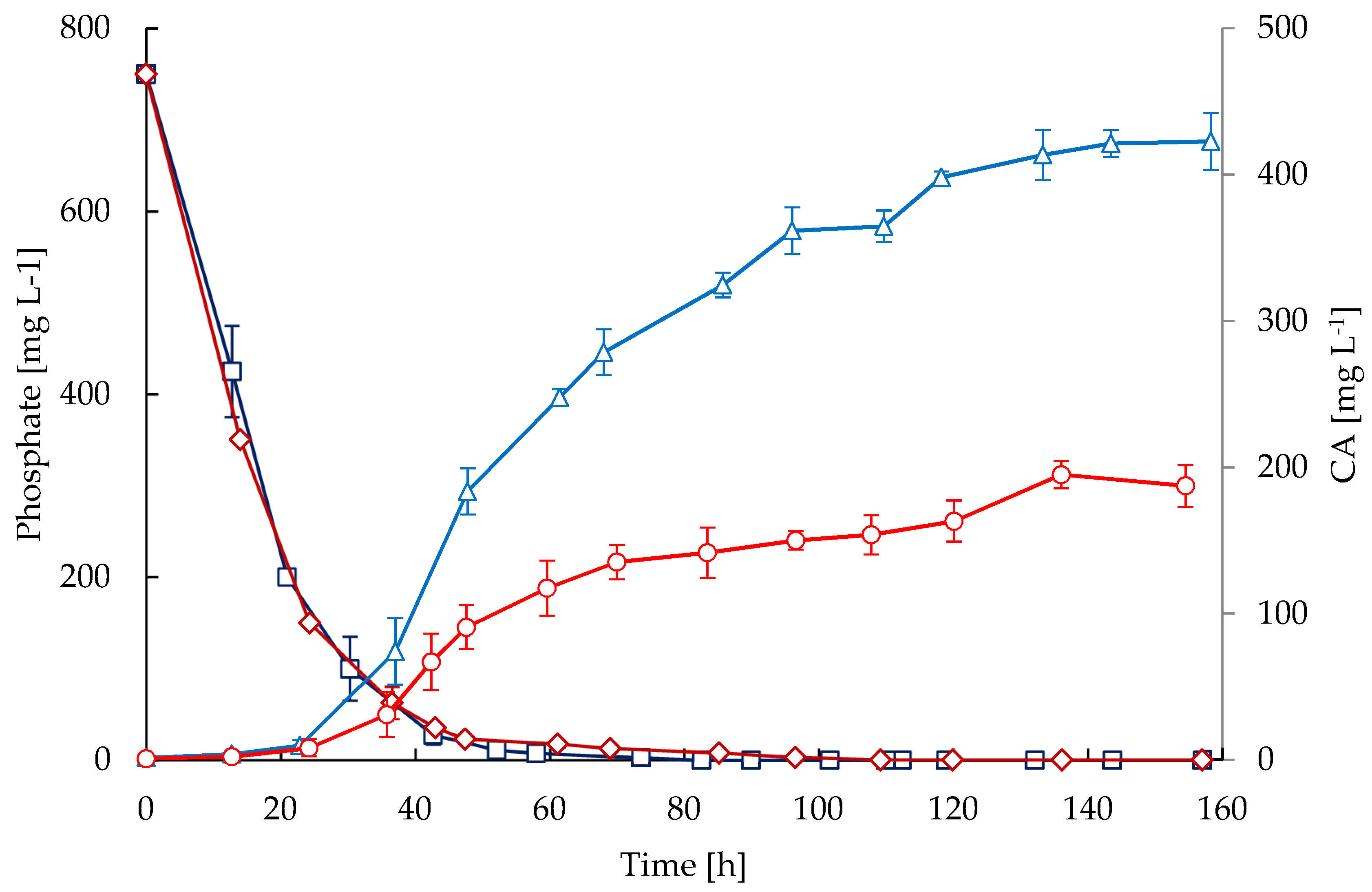
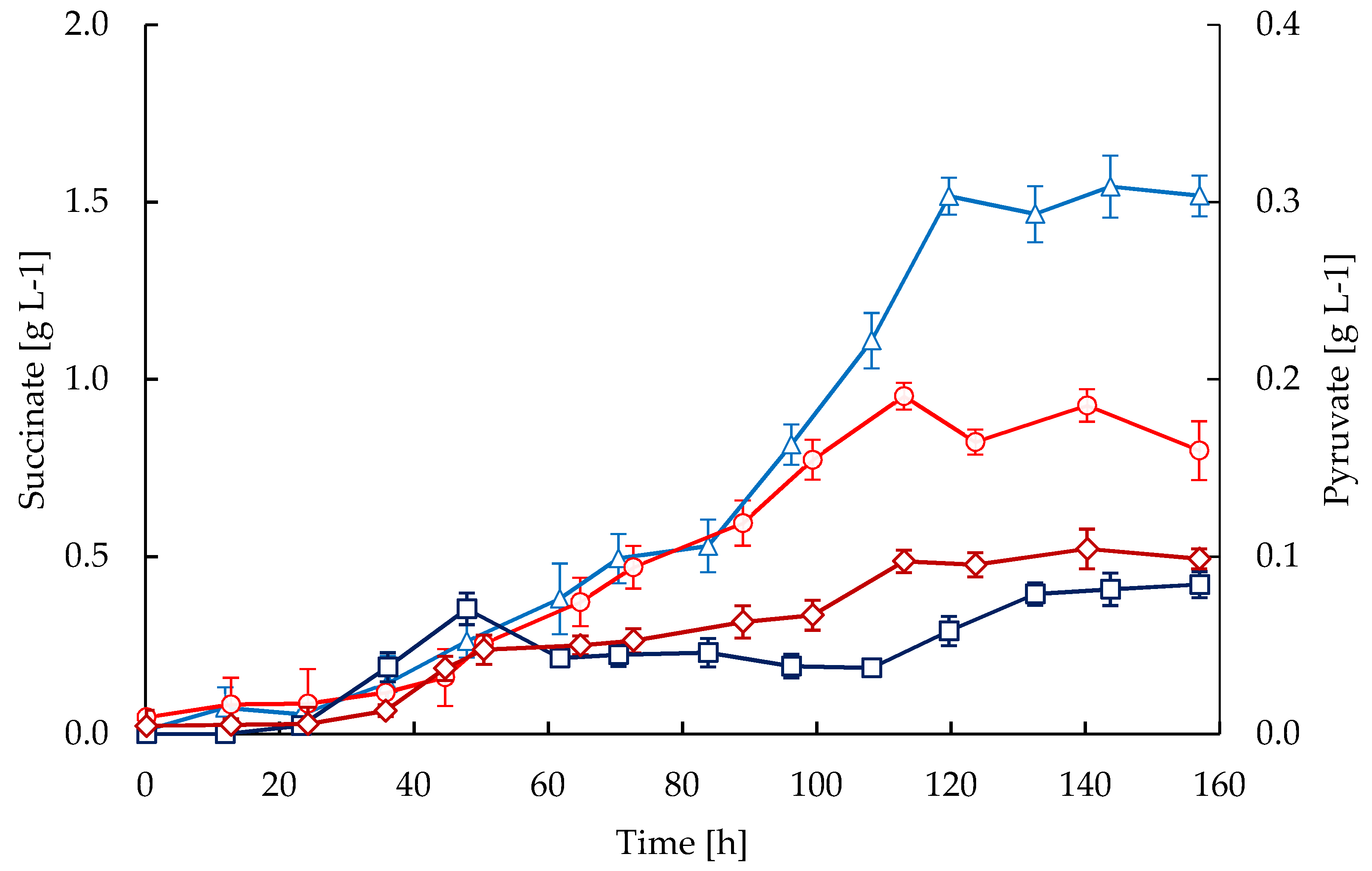
2.3. Substrates and Products
3. Discussion
4. Materials and Methods
4.1. Strain and Cultivation Procedures
4.2. Mycelium Measurement and Product Quantification
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ramirez-Malule, H. Bibliometric Analysis of Global Research on Clavulanic Acid. Antibiotics 2018, 7, 102. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ríos, D.; Ramírez-Malule, H. Bibliometric analysis of recent research on multidrug and antibiotics resistance (2017–2018). J. Appl. Pharm. Sci. 2019, 9, 112–116. [Google Scholar] [CrossRef]
- Cerri, M.O.; Badino, A.C. Shear conditions in clavulanic acid production by Streptomyces clavuligerus in stirred tank and airlift bioreactors. Bioprocess Biosyst. Eng. 2012, 35, 977–984. [Google Scholar] [CrossRef]
- Rosa, J.C.; Baptista Neto, A.; Hokka, C.O.; Badino, A.C. Influence of dissolved oxygen and shear conditions on clavulanic acid production by Streptomyces clavuligerus. Bioprocess Biosyst. Eng. 2005, 27, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ríos, D.; Ramírez-Malule, H.; Neubauer, P.; Junne, S.; Ríos-Estepa, R. Degradation kinetics of clavulanic acid in fermentation broths at low temperatures. Antibiotics 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ríos, D.; Ramírez-Malule, H.; Neubauer, P.; Junne, S.; Ríos-Estepa, R. Data of clavulanic acid and clavulanate-imidazole stability at low temperatures. Data Br. 2019, 23, 103775. [Google Scholar] [CrossRef] [PubMed]
- Ser, H.; Law, J.W.; Chaiyakunapruk, N.; Jacop, S.A.; Palanisamy, U.D.; Chan, K.-G.; Goh, B.-H.; Lee, L. Fermentation conditions that affect clavulanic acid production in Streptomyces clavuligerus: A systematic review. Front. Microbiol. 2016, 7, 522. [Google Scholar] [CrossRef]
- Qin, R.; Zhong, C.; Zong, G.; Fu, J.; Pang, X.; Cao, G. Improvement of clavulanic acid production in Streptomyces clavuligerus F613-1 by using a claR-neo reporter strategy. Electron. J. Biotechnol. 2017, 28, 41–46. [Google Scholar] [CrossRef]
- Jnawali, H.N.; Yoo, J.C.; Sohng, J.K. Improvement of clavulanic acid production in Streptomyces clavuligerus by genetic manipulation of structural biosynthesis genes. Biotechnol. Lett. 2011, 33, 1221–1226. [Google Scholar] [CrossRef]
- Kizildoğan, A.K.; Jaccard, G.V.; Mutlu, A.; Sertdemir, I.; Özcengiz, G. Genetic engineering of an industrial strain of Streptomyces clavuligerus for further enhancement of clavulanic acid production. Turk. J. Biol. 2017, 41, 342–353. [Google Scholar] [CrossRef]
- Olmos, E.; Mehmood, N.; Haj Husein, L.; Goergen, J.L.; Fick, M.; Delaunay, S. Effects of bioreactor hydrodynamics on the physiology of Streptomyces. Bioprocess Biosyst. Eng. 2013, 36, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Manteca, Á.; Yagüe, P. Streptomyces differentiation in liquid cultures as a trigger of secondary metabolism. Antibiotics 2018, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Lin, S.; Xia, X.X.; Cong, F.S.; Zhong, J.J. Significance of agitation-induced shear stress on mycelium morphology and lavendamycin production by engineered Streptomyces flocculus. Appl. Microbiol. Biotechnol. 2014, 98, 4399–4407. [Google Scholar] [CrossRef] [PubMed]
- Zacchetti, B.; Smits, P.; Claessen, D. Dynamics of pellet fragmentation and aggregation in liquid-grown cultures of Streptomyces lividans. Front. Microbiol. 2018, 9, 943. [Google Scholar] [CrossRef] [PubMed]
- Manteca, A.; Alvarez, R.; Salazar, N.; Yagüe, P.; Sanchez, J. Mycelium differentiation and antibiotic production in submerged cultures of Streptomyces coelicolor. Appl. Environ. Microbiol. 2008, 74, 3877–3886. [Google Scholar] [CrossRef]
- Ramirez-Malule, H.; Junne, S.; Nicolás Cruz-Bournazou, M.; Neubauer, P.; Ríos-Estepa, R. Streptomyces clavuligerus shows a strong association between TCA cycle intermediate accumulation and clavulanic acid biosynthesis. Appl. Microbiol. Biotechnol. 2018, 102, 4009–4023. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.S.; Vieira, L.M.; Pons, M.N.; Fonseca, M.M.R.; Menezes, J.C. Morphology and viability analysis of Streptomyces clavuligerus in industrial cultivation systems. Bioprocess Biosyst. Eng. 2004, 26, 177–184. [Google Scholar] [CrossRef]
- Eibl, R.; Kaiser, S.; Lombriser, R. Disposable bioreactors: The current state-of-the-art and recommended applications in biotechnology. Appl. Microbiol. Biotechnol. 2010, 41–49. [Google Scholar] [CrossRef]
- Junne, S.; Solymosi, T.; Oosterhuis, N.; Neubauer, P. Cultivation of cells and microorganisms in wave-mixed disposable bag bioreactors at different scales. Chem. Ing. Tech. 2012, 85, 57–66. [Google Scholar] [CrossRef]
- DeFife, K.; Leigh, P. Versatility of a single-use bioreactor platform for culture of diverse cell types. BioPharm Int. 2009, 22, 30–37. [Google Scholar]
- Löffelholz, C.; Kaiser, S.C.; Kraume, M.; Eibl, R.; Eibl, D. Dynamic Single-Use Bioreactors Used in Modern Liter- and m3- Scale Biotechnological Processes: Engineering Characteristics and Scaling Up. In Disposable Bioreactors II; Eibl, D., Eibl, R., Eds.; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 138, pp. 1–6. [Google Scholar]
- Odeleye, A.O.O.; Marsh, D.T.J.; Osborne, M.D.; Lye, G.J.; Micheletti, M. On the fluid dynamics of a laboratory scale single-use stirred bioreactor. Chem. Eng. Sci. 2014, 111, 299–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillig, F.; Porscha, N.; Junne, S.; Neubauer, P. Growth and docosahexaenoic acid production performance of the heterotrophic marine microalgae Crypthecodinium cohnii in the wave-mixed single-use reactor CELL-tainer. Eng. Life Sci. 2014, 14, 254–263. [Google Scholar] [CrossRef]
- Westbrook, A.; Scharer, J.; Moo-young, M.; Oosterhuis, N.; Chou, C.P. Application of a two-dimensional disposable rocking bioreactor to bacterial cultivation for recombinant protein production. Biochem. Eng. J. 2014, 88, 154–161. [Google Scholar] [CrossRef]
- Kurt, T.; Maria, A.; Ardébol, M.; Turan, Z.; Neubauer, P.; Junne, S.; Meyer, V. Rocking Aspergillus: Morphology-controlled cultivation of Aspergillus niger in a wave-mixed bioreactor for the production of secondary metabolites. Microb. Cell Fact. 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bushell, M.E.; Kirk, S.; Zhao, H.-J.; Avignone-Rossa, C.A. Manipulation of the physiology of clavulanic acid biosynthesis with the aid of metabolic flux analysis. Enzyme Microb. Technol. 2006, 39, 149–157. [Google Scholar] [CrossRef]
- Daub, A.; Böhm, M.; Delueg, S.; Mühlmann, M.; Schneider, G.; Büchs, J. Characterization of hydromechanical stress in aerated stirred tanks up to 40 m3 scale by measurement of maximum stable drop size. J. Biol. Eng. 2014, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Baldi, S.; Yianneskis, M. On the quantification of energy dissipation in the impeller stream of a stirred vessel from fluctuating velocity gradient measurements. Chem. Eng. Sci. 2004, 59, 2659–2671. [Google Scholar] [CrossRef]
- Öncul, A.A.; Kalmbach, A.; Genzel, Y.; Reichl, U.; Thévenin, D. Characterization of flow conditions in 2 L and 20 L wave bioreactors using computational fluid dynamics. Biotechnol. Prog. 2010, 26, 101–110. [Google Scholar] [CrossRef]
- Rollins, M.J.; Jensen, S.E.; Westlake, D.W.S. Effect of dissolved oxygen level on ACV synthetase synthesis and activity during growth of Streptomyces clavuligerus. Appl. Microbiol. Biotechnol. 1991, 35, 83–88. [Google Scholar] [CrossRef]
- Duobin, M.; Yuping, M.; Lujing, G.; Aijing, Z.; Jianqiang, Z.; Chunping, X. Fermentation characteristics in stirred-tank reactor of exopolysaccharides with hypolipidemic activity produced by Pleurotus geesteranus 5#. An. Acad. Bras. Cienc. 2013, 85, 1473–1481. [Google Scholar] [CrossRef]
- Chisti, Y. Hydrodynamic damage to animal cells. Crit. Rev. Biotechnol. 2001, 21, 67–110. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, J.C.; Araujo, M.L.G.C.; Hokka, C.O.; Badino, A.C. Influence of glycerol and ornithine feeding on clavulanic acid production by Streptomyces clavuligerus. Braz. J. Chem. Eng. 2010, 27, 499–506. [Google Scholar] [CrossRef]
- Costa, C.L.L.; Badino, A.C. Production of clavulanic acid by Streptomyces clavuligerus in batch cultures without and with glycerol pulses under different temperature conditions. Biochem. Eng. J. 2012, 69, 1–7. [Google Scholar] [CrossRef]
- Zhan, C.; Hagrot, E.; Brandt, L.; Chotteau, V. Study of hydrodynamics in wave bioreactors by computational fluid dynamics reveals a resonance phenomenon. Chem. Eng. Sci. 2019, 193, 53–65. [Google Scholar] [CrossRef]
- Buffo, M.M.; Corrêa, L.J.; Esperança, M.N.; Cruz, A.J.G.; Farinas, C.S.; Badino, A.C. Influence of dual-impeller type and configuration on oxygen transfer, power consumption, and shear rate in a stirred tank bioreactor. Biochem. Eng. J. 2016, 114. [Google Scholar] [CrossRef]
- Campesi, A.; Cerri, M.O.; Hokka, C.O.; Badino, A.C. Determination of the average shear rate in a stirred and aerated tank bioreactor. Bioprocess Biosyst. Eng. 2009, 32, 241–248. [Google Scholar] [CrossRef]
- Yagüe, P.; López-García, M.T.; Rioseras, B.; Sánchez, J.; Manteca, Á. Pre-sporulation stages of Streptomyces differentiation: State-of-the-art and future perspectives. FEMS Microbiol. Lett. 2013, 342, 79–88. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–44. [Google Scholar] [CrossRef]
- Veiter, L.; Rajamanickam, V.; Herwig, C. The filamentous fungal pellet—Relationship between morphology and productivity. Appl. Microbiol. Biotechnol. 2018, 102, 2997–3006. [Google Scholar] [CrossRef]
- Yin, P.; Wang, Y.H.; Zhang, S.L.; Chu, J.; Zhuang, Y.P.; Chen, N.; Li, X.F.; Wu, Y.B. Effect of mycelial morphology on bioreactor performance and avermectin production of Streptomyces avermitilis in submerged cultivations. J. Chin. Inst. Chem. Eng. 2008, 39, 609–615. [Google Scholar] [CrossRef]
- Kumar, P.; Dubey, K.K. Mycelium transformation of Streptomyces toxytricini into pellet: Role of culture conditions and kinetics. Bioresour. Technol. 2017, 228, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Ohta, N.; Park, Y.S.; Yahiro, K.; Okabe, M. Comparison of neomycin production from Streptomyces fradiae cultivation using soybean oil as the sole carbon source in an air-lift bioreactor and a stirred-tank reactor. J. Ferment. Bioeng. 1995, 79, 443–448. [Google Scholar] [CrossRef]
- Elsayed, E.A.; Farid, M.A.; El-Enshasy, H.A. Enhanced Natamycin production by Streptomyces natalensis in shake-flasks and stirred tank bioreactor under batch and fed-batch conditions. BMC Biotechnol. 2019, 19, 46. [Google Scholar] [CrossRef] [PubMed]
- El Enshasy, H.; Farid, M.; Elsayed, E. Influence of inoculum type and cultivation conditions on natamycin production by Streptomyces natalensis. J. Basic Microbiol. 2000, 40, 333–342. [Google Scholar] [CrossRef]
- Mehmood, N.; Olmos, E.; Marchal, P.; Goergen, J.L.; Delaunay, S. Relation between pristinamycins production by Streptomyces pristinaespiralis, power dissipation and volumetric gas-liquid mass transfer coefficient, kLa. Process Biochem. 2010, 45, 1779–1786. [Google Scholar] [CrossRef]
- Saudagar, P.S.; Singhal, R.S. Optimization of nutritional requirements and feeding strategies for clavulanic acid production by Streptomyces clavuligerus. Bioresour. Technol. 2007, 98, 2010–2017. [Google Scholar] [CrossRef] [PubMed]
- Arulanantham, H.; Kershaw, N.J.; Hewitson, K.S.; Hughes, C.E.; Thirkettle, J.E.; Schofield, C.J. ORF17 from the clavulanic acid biosynthesis gene cluster catalyzes the ATP-dependent formation of N-glycyl-clavaminic acid. J. Biol. Chem. 2006, 281, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yu, C.; Yao, J.; Wang, Z.; Lu, S. An Online Respiratory quotient-feedback strategy of feeding yeast extract for efficient arachidonic acid production by Mortierella alpina. Front. Bioeng. Biotechnol. 2018, 5, 83. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, D.A. Primary metabolism and its control in streptomycetes: A most unusual group of bacteria. Adv. Microb. Physiol. 2000, 42, 47–238. [Google Scholar] [PubMed]
- Gamboa-Suasnavart, R.A.; Valdez-Cruz, N.A.; Gaytan-Ortega, G.; Cereceda-Reynoso, G.I.; Cabrera-Santos, D.; López-Griego, L.; Klöckner, W.; Büchs, J.; Trujillo-Roldán, M.A. The metabolic switch can be activated in a recombinant strain of Streptomyces lividans by a low oxygen transfer rate in shake flasks. Microb. Cell Fact. 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Viollier, P.H.; Minas, W.; Dale, G.E.; Folcher, M.; Thompson, C.J. Role of Acid Metabolism in Streptomyces coelicolor morphological differentiation and antibiotic biosynthesis. J. Bacteriol. 2001, 183, 3184–3192. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Shi, Z.; Gao, P.; Feng, H.; Duan, Z.; Mao, Z. On-line optimization of glutamate production based on balanced metabolic control by RQ. Bioprocess Biosyst. Eng. 2006, 29, 109–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roubos, J.A.; Krabben, P.; De Laat, W.; Heijnen, J.J. Clavulanic acid degradation in Streptomyces clavuligerus fed-batch cultivations. Biotechnol. Prog. 2002, 18, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B. Energy dissipation and shear rate with geometry of baffled surface aerator. Chem. Eng. Res. Bull. 2010, 14, 92–96. [Google Scholar] [CrossRef]
- Devi, T.T.; Kumar, B. Mass transfer and power characteristics of stirred tank with Rushton and curved blade impeller. Eng. Sci. Technol. Int. J. 2017, 20, 730–737. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.S.; Porcel, E.R.; López, J.C.; Sevilla, J.F.; Chisti, Y. Shear rate in stirred tank and bubble column bioreactors. Chem. Eng. J. 2006, 124, 1–5. [Google Scholar] [CrossRef]
- Eibl, D.; Eibl, R. Disposable Bioreactors II; Scheper, T., Ed.; Advances in Biochemical Engineering/Biotechnology; Springer: Berlin/Heidelberg, Germany, 2014; Volume 138, pp. 11–38. [Google Scholar]
- Rasband, W.S. ImageJ; US National Institutes of Health: Bethesda, MD, USA, 2018.
- Ramirez-Malule, H.; Junne, S.; López, C.; Zapata, J.; Sáez, A.; Neubauer, P.; Rios-Estepa, R. An improved HPLC-DAD method for clavulanic acid quantification in fermentation broths of Streptomyces clavuligerus. J. Pharm. Biomed. Anal. 2016, 120, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Junne, S.; Klingner, A.; Kabisch, J.; Schweder, T.; Neubauer, P. A two-compartment bioreactor system made of commercial parts for bioprocess scale-down studies: Impact of oscillations on Bacillus subtilis fed-batch cultivations. Biotechnol. J. 2011, 6, 1009–1017. [Google Scholar] [CrossRef]
- Lemoine, A.; Maya Martnez-Iturralde, N.; Spann, R.; Neubauer, P.; Junne, S. Response of Corynebacterium glutamicum exposed to oscillating cultivation conditions in a two- and a novel three-compartment scale-down bioreactor. Biotechnol. Bioeng. 2015, 112, 1220–1231. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioreactor | DCW (g·L−1) | V (L) | N (rpm) | kLa (h−1) | τmax (Pa) | µapp (Pa s) |
|---|---|---|---|---|---|---|
| CT | 0.5 | 5.0 | 12 | 73.25 ± 7.40 | 0.066 ± 0.010 | 0.0011 ± 0.0005 |
| 2.1 | 5.0 | 15 | 43.90 ± 0.92 | 0.102 ± 0.068 | 0.0012 ± 0.0001 | |
| 3.4 | 5.0 | 17 | 61.71 ± 8.13 | 0.222 ± 0.016 | 0.0021± 0.0013 | |
| 4.7 | 5.1 | 20 | 80.52 ± 8.86 | 0.147 ± 0.041 | 0.0041 ± 0.0015 | |
| 9.3 | 5.8 | 25 | 208.10 ± 28.79 | 0.572 ± 0.033 | 0.0055 ± 0.0005 | |
| 10.1 | 6.1 | 22 | 121.88 ± 3.26 | 0.747 ± 0.079 | 0.0083 ± 0.0007 | |
| 10.3 | 7.8 | 22 | 83.73 ± 7.15 | 0.634 ± 0.029 | 0.0096 ± 0.0007 | |
| STR | 1.9 | 5.0 | 300 | 39.11 ± 2.80 | 1.472 ± 0.061 | 0.0013 ± 0.0001 |
| 4.3 | 5.0 | 320 | 35.49 ± 1.99 | 3.091 ± 0.228 | 0.0034 ± 0.0003 | |
| 6.5 | 5.0 | 410 | 50.57 ± 2.40 | 5.047 ± 0.538 | 0.0051 ± 0.0006 | |
| 7.7 | 5.2 | 450 | 58.61 ± 3.46 | 5.883 ± 0.645 | 0.0058 ± 0.0016 | |
| 9.3 | 6.4 | 500 | 61.92 ± 3.78 | 6.460 ± 0.381 | 0.0088 ± 0.0017 | |
| 12.1 | 7.8 | 500 | 54.85 ± 0.26 | 7.563 ± 0.255 | 0.0145 ± 0.0010 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Ríos, D.; Junne, S.; Neubauer, P.; Ochoa, S.; Ríos-Estepa, R.; Ramírez-Malule, H. Characterization of the Metabolic Response of Streptomyces clavuligerus to Shear Stress in Stirred Tanks and Single-Use 2D Rocking Motion Bioreactors for Clavulanic Acid Production. Antibiotics 2019, 8, 168. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040168
Gómez-Ríos D, Junne S, Neubauer P, Ochoa S, Ríos-Estepa R, Ramírez-Malule H. Characterization of the Metabolic Response of Streptomyces clavuligerus to Shear Stress in Stirred Tanks and Single-Use 2D Rocking Motion Bioreactors for Clavulanic Acid Production. Antibiotics. 2019; 8(4):168. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040168
Chicago/Turabian StyleGómez-Ríos, David, Stefan Junne, Peter Neubauer, Silvia Ochoa, Rigoberto Ríos-Estepa, and Howard Ramírez-Malule. 2019. "Characterization of the Metabolic Response of Streptomyces clavuligerus to Shear Stress in Stirred Tanks and Single-Use 2D Rocking Motion Bioreactors for Clavulanic Acid Production" Antibiotics 8, no. 4: 168. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics8040168