Tackling Intrinsic Antibiotic Resistance in Serratia marcescens with a Combination of Ampicillin/Sulbactam and Phage SALSA


Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of S. marcescens Clinical Isolates
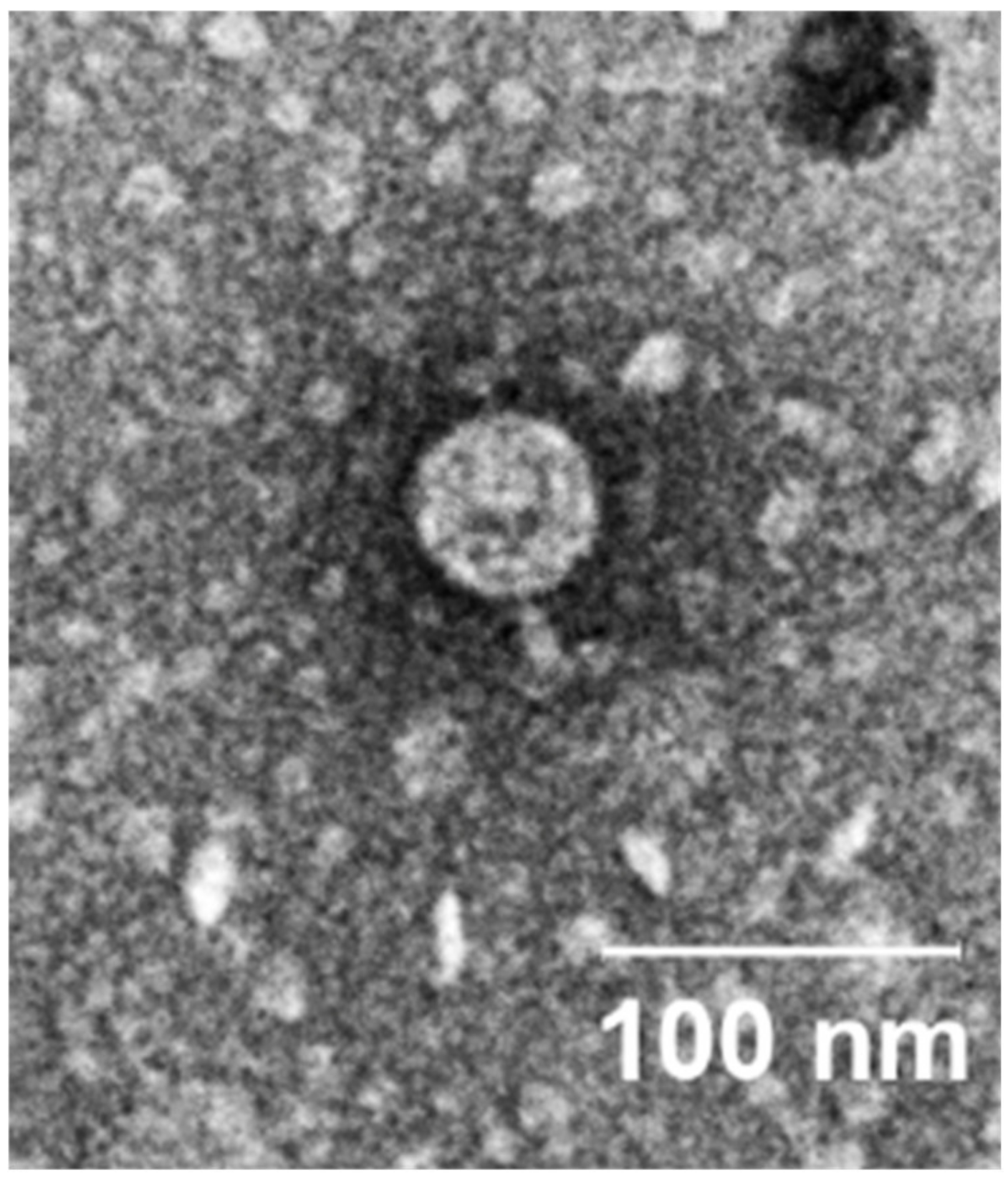
2.2. Phenotypic Characterization of vB_SmaP-SALSA
2.3. Genome Analysis of Phage SALSA
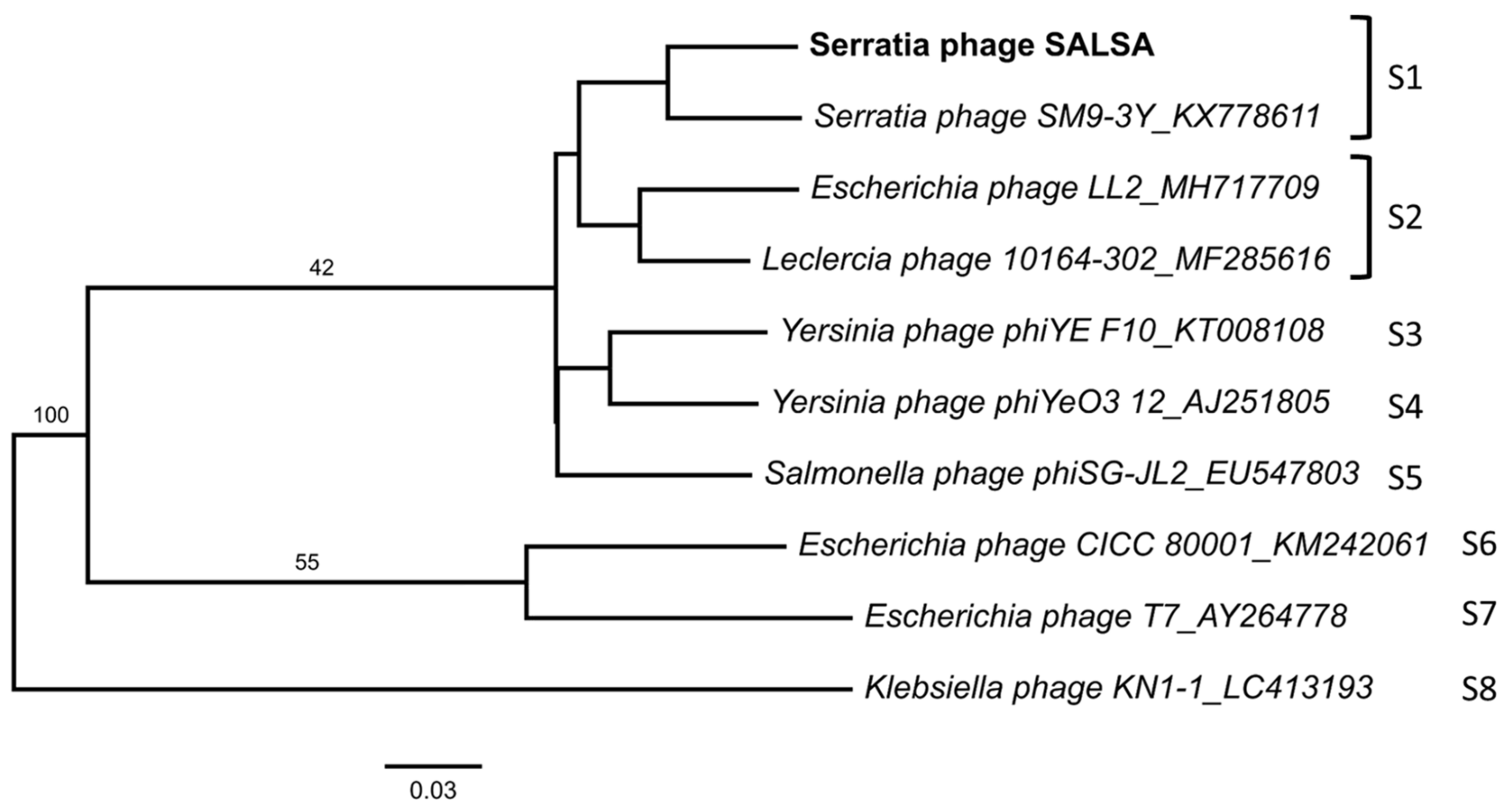
2.4. Diversity of Known Serratia Phages
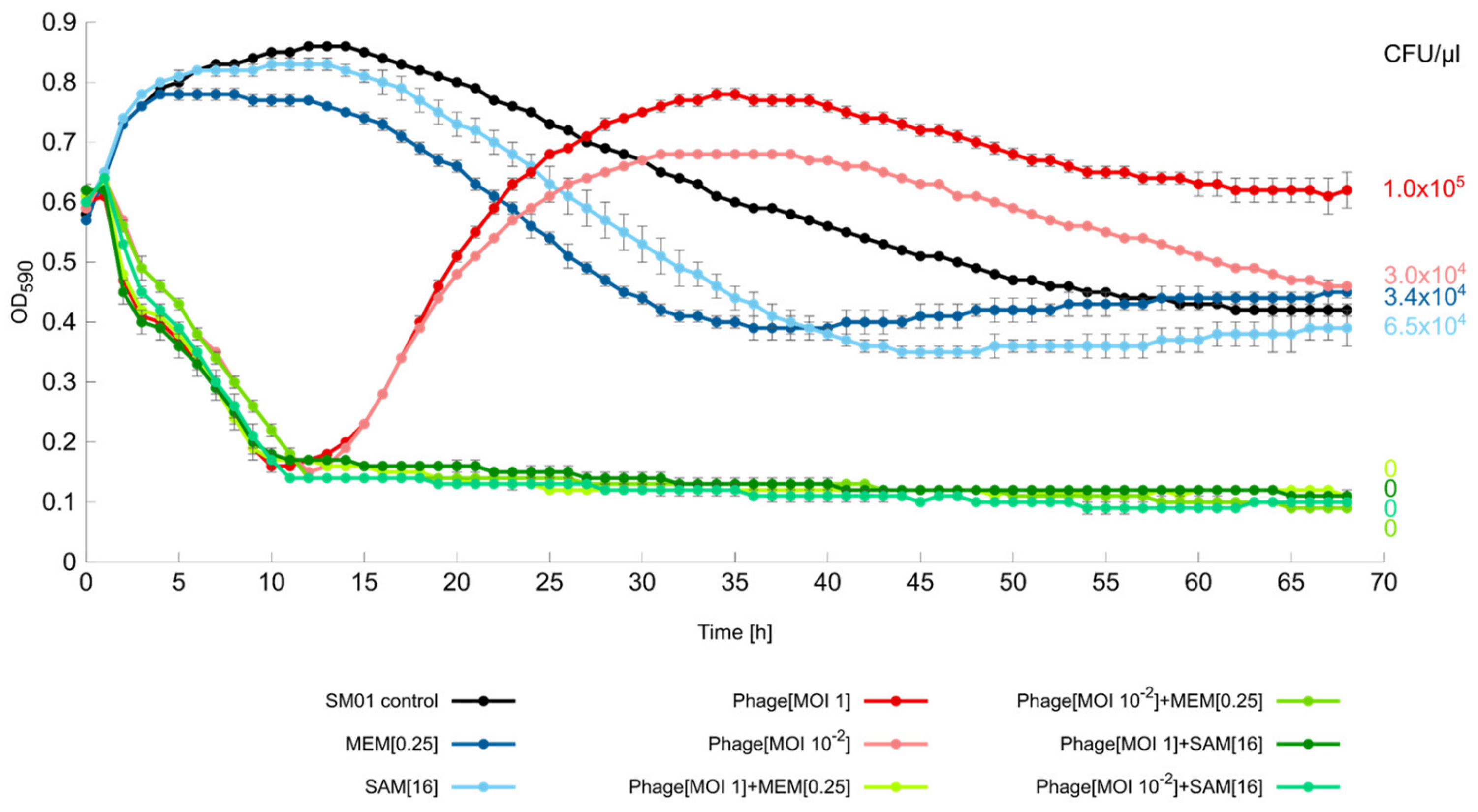
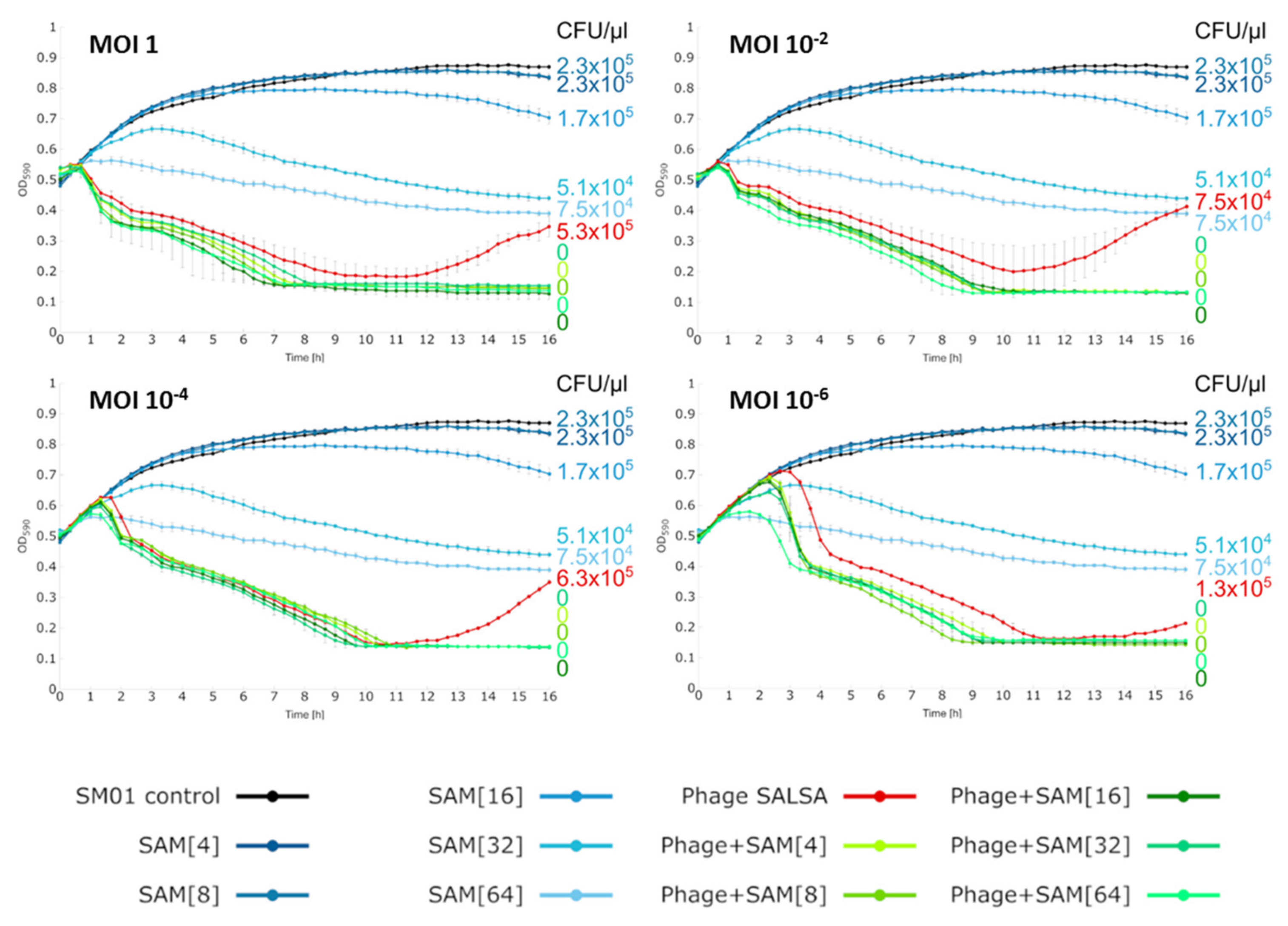
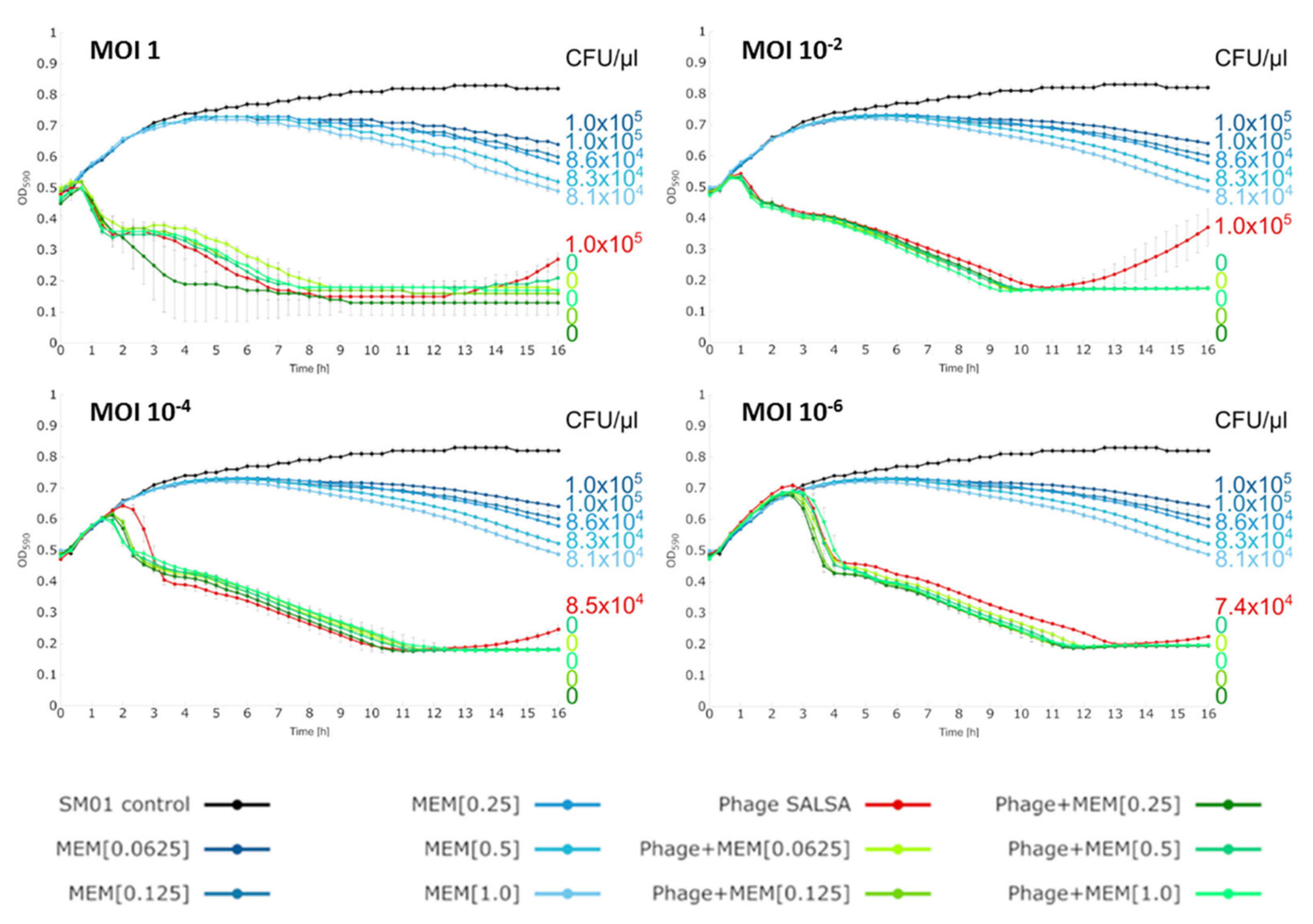
2.5. Liquid Infection Assays
3. Materials and Methods
3.1. Bacterial Strains and Phage Isolation
3.2. Characterization of the Clinical Isolates of S. marcescens
3.3. Host Range
3.4. Transmission Electron Microscopy (TEM)
3.5. Phage Genome Sequencing and Analysis
3.6. Liquid Infection Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aslam, S.; Courtwright, A.M.; Koval, C.; Lehman, S.M.; Morales, S.; Furr, C.L.; Rosas, F.; Brownstein, M.J.; Fackler, J.R.; Sisson, B.M.; et al. Early clinical experience of bacteriophage therapy in 3 lung transplant recipients. Am. J. Transplant. 2019, 19, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Verbeken, G.; Pirnay, J.-P.; Lavigne, R.; Jennes, S.; De Vos, D.; Casteels, M.; Huys, I. Call for a dedicated European legal framework for bacteriophage therapy. Arch. Immunol. Ther. Exp. (Warsz.) 2014, 62, 117–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutateladze, M.; Adamia, R. Bacteriophages as potential new therapeutics to replace or supplement antibiotics. Trends Biotechnol. 2010, 28, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, F. Resistance development to bacteriophages occurring during bacteriophage therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Tagliaferri, T.L.; Jansen, M.; Horz, H.-P. Fighting pathogenic bacteria on two fronts: Phages and antibiotics as combined strategy. Front. Cell. Infect. Microbiol. 2019, 9, 22. [Google Scholar] [CrossRef]
- Jansen, M.; Wahida, A.; Latz, S.; Krüttgen, A.; Häfner, H.; Buhl, E.M.; Ritter, K.; Horz, H.-P. Enhanced antibacterial effect of the novel T4-like bacteriophage KARL-1 in combination with antibiotics against multi-drug resistant Acinetobacter baumannii. Sci. Rep. 2018, 8, 14140. [Google Scholar] [CrossRef] [Green Version]
- Torres-Barceló, C.; Hochberg, M.E. Evolutionary rationale for phages as complements of antibiotics. Trends Microbiol. 2016, 24, 249–256. [Google Scholar] [CrossRef]
- Macdougall, C. Beyond susceptible and resistant, Part I: Treatment of infections due to gram-negative organisms with inducible β-Lactamases. J. Pediatr. Pharmacol. Ther. JPPT Off. J. PPAG 2011, 16, 23–30. [Google Scholar]
- Cristina, M.; Sartini, M.; Spagnolo, A. Serratia marcescens Infections in neonatal intensive care units (NICUs). Int. J. Environ. Res. Public Health 2019, 16, 610. [Google Scholar] [CrossRef] [Green Version]
- Iguchi, A.; Nagaya, Y.; Pradel, E.; Ooka, T.; Ogura, Y.; Katsura, K.; Kurokawa, K.; Oshima, K.; Hattori, M.; Parkhill, J.; et al. Genome evolution and plasticity of Serratia marcescens, an important multidrug-resistant nosocomial pathogen. Genome Biol. Evol. 2014, 6, 2096–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastmeier, P.; Loui, A.; Stamm-Balderjahn, S.; Hansen, S.; Zuschneid, I.; Sohr, D.; Behnke, M.; Obladen, M.; Vonberg, R.-P.; Rüden, H. Outbreaks in neonatal intensive care units—They are not like others. Am. J. Infect. Control. 2007, 35, 172–176. [Google Scholar] [CrossRef] [PubMed]
- Mahlen, S.D. Serratia Infections: From military experiments to current practice. Clin. Microbiol. Rev. 2011, 24, 755–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maragakis, L.L.; Winkler, A.; Tucker, M.G.; Cosgrove, S.E.; Ross, T.; Lawson, E.; Carroll, K.C.; Perl, T.M. Outbreak of multidrug-resistant Serratia marcescens infection in a neonatal intensive care unit. Infect. Control. Hosp. Epidemiol. 2008, 29, 418–423. [Google Scholar] [CrossRef]
- Krzyżaniak, N.; Pawłowska, I.; Bajorek, B. Review of drug utilization patterns in NICUs worldwide. J. Clin. Pharm. Ther. 2016, 41, 612–620. [Google Scholar] [CrossRef]
- Abedon, S.T. Lysis from without. Bacteriophage 2011, 1, 46–49. [Google Scholar] [CrossRef]
- Teng, T.; Zhang, G.; Fan, X.; Zhang, Z.; Zhang, L.; Wu, D.; Chen, S.; Li, Y.; Jin, J. Complete genome sequence analysis of PS2, a novel T4-like bacteriophage that infects Serratia marcescens clinical isolates. Arch. Virol. 2018, 163, 1997–2000. [Google Scholar] [CrossRef]
- Tian, C.; Zhao, J.; Zhang, Z.; Chen, X.; Wei, X.; Li, H.; Lin, W.; Ke, Y.; Hu, L.; Jiang, A.; et al. Identification and molecular characterization of Serratia marcescens phages vB_SmaA_2050H1 and vB_SmaM_2050HW. Arch. Virol. 2019, 164, 1085–1094. [Google Scholar] [CrossRef]
- Yosef, I.; Goren, M.G.; Globus, R.; Molshanski-Mor, S.; Qimron, U. Extending the host range of bacteriophage particles for DNA Transduction. Mol. Cell 2017, 66, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Huss, P.; Raman, S. Engineered bacteriophages as programmable biocontrol agents. Curr. Opin. Biotechnol. 2020, 61, 116–121. [Google Scholar] [CrossRef]
- Bilen, M.; Dufour, J.-C.; Lagier, J.-C.; Cadoret, F.; Daoud, Z.; Dubourg, G.; Raoult, D. The contribution of culturomics to the repertoire of isolated human bacterial and archaeal species. Microbiome 2018, 6, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.-W. Frequency of morphological phage descriptions in the year 2000. Arch. Virol. 2001, 146, 843–857. [Google Scholar] [CrossRef]
- Hao, Y.; Shi, H.; Li, J.; Zhao, C.; Huang, H.; Sun, Y. Complete genome sequence of bacteriophage SM9-3Y infecting Serratia marcescens. Genome Announc. 2018, 6, e0127-17. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. VICTOR: Genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 2017, 33, 3396–3404. [Google Scholar] [CrossRef] [Green Version]
- Pajunen, M.I.; Kiljunen, S.J.; Söderholm, M.E.-L.; Skurnik, M. Complete genomic sequence of the lytic bacteriophage φYeO3-12 of Yersinia enterocolitica serotype O:3. J. Bacteriol. 2001, 183, 1928–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.-J.; Cho, S.-H.; Kim, T.-E.; Won, Y.-J.; Jeong, J.; Park, S.C.; Kim, J.-H.; Yoo, H.-S.; Park, Y.-H.; Kim, S.-J. Characterization of a T7-Like Lytic Bacteriophage (φSG-JL2) of Salmonella enterica Serovar Gallinarum Biovar Gallinarum. Appl. Environ. Microbiol. 2008, 74, 6970–6979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denyes, J.M.; Krell, P.J.; Manderville, R.A.; Ackermann, H.-W.; She, Y.-M.; Kropinski, A.M. The genome and proteome of Serratia bacteriophage η which forms unstable lysogens. Virol. J. 2014, 11, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casey, E.; Fitzgerald, B.; Mahony, J.; Lugli, G.A.; Ventura, M.; van Sinderen, D. Genome sequence of Serratia marcescens phage BF. Genome Announc. 2017, 5, e00211-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, S.; Nguyen, Q.; Newkirk, H.; Liu, M.; Cahill, J.; Ramsey, J. Complete Genome sequence of Serratia marcescens Myophage MyoSmar. Microbiol. Resour. Announc. 2019, 8, e01046-19. [Google Scholar] [CrossRef] [Green Version]
- Price, L.; Rohren, M.; Newkirk, H.; Liu, M.; Ramsey, J. Complete genome sequence of Serratia marcescens Myophage Moabite. Microbiol. Resour. Announc. 2019, 8, e00741-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, K.; Freeman, M.; Newkirk, H.; Liu, M.; Cahill, J.; Ramsey, J. Complete genome sequence of Serratia marcescens phage MTx. Microbiol. Resour. Announc. 2019, 8, e00573-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, S.; Atkison, C.; Moreland, R.; Liu, M.; Ramsey, J.; Leavitt, J. Complete genome sequence of Serratia phage Muldoon. Microbiol. Resour. Announc. 2020, 9, e01418-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snowden, J.; Newkirk, H.; Moreland, R.; Liu, M.; Ramsey, J.; Leavitt, J. Complete genome sequence of Serratia marcescens Siphophage Slocum. Microbiol. Resour. Announc. 2020, 9, e01421-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, E.A.; Hopson, H.; Rodriguez, A.; Kongari, R.; Bonasera, R.; Hernandez-Morales, A.C.; Liu, M. Complete genome sequence of Serratia marcescens Siphophage Serbin. Microbiol. Resour. Announc. 2019, 8, e00042-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehler, B.T.; Hopson, H.; Kongari, R.; Bonasera, R.; Hernandez-Morales, A.C.; Liu, M. Complete genome sequence of Serratia marcescens Siphophage Scapp. Microbiol. Resour. Announc. 2019, 8, e00417-19. [Google Scholar] [CrossRef] [Green Version]
- Melbern, L.; Broussard, K.; Moreland, R.; Liu, M.; Ramsey, J.; Leavitt, J. Complete genome sequence of Serratia marcescens Podophage Pila. Microbiol. Resour. Announc. 2020, 9, e01420-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bockoven, R.; Gutierrez, J.; Newkirk, H.; Liu, M.; Cahill, J.; Ramsey, J. Complete genome sequence of Serratia marcescens Podophage Parlo. Microbiol. Resour. Announc. 2019, 8, e00569-19. [Google Scholar] [CrossRef] [Green Version]
- Matilla, M.A.; Salmond, G.P.C. Bacteriophage ϕMAM1, a Viunalikevirus, is a broad-host-range, high-efficiency generalized transducer that infects environmental and clinical isolates of the enterobacterial genera Serratia and Kluyvera. Appl. Environ. Microbiol. 2014, 80, 6446–6457. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Ma, T.; Zhang, X.; Huang, Y.; Mi, Z.; Sun, Q.; An, X.; Fan, H.; Wu, S.; Wei, L.; et al. First complete genome sequence of a virulent bacteriophage infecting the opportunistic pathogen Serratia rubidaea. Arch. Virol. 2017, 162, 2021–2028. [Google Scholar] [CrossRef]
- Malone, L.M.; Warring, S.L.; Jackson, S.A.; Warnecke, C.; Gardner, P.P.; Gumy, L.F.; Fineran, P.C. A jumbo phage that forms a nucleus-like structure evades CRISPR–Cas DNA targeting but is vulnerable to type III RNA-based immunity. Nat. Microbiol. 2020, 5, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Akusobi, C.; Fang, X.; Salmond, G.P.C. Environmental T4-family bacteriophages evolve to escape abortive infection via multiple routes in a bacterial host employing “altruistic suicide” through Type III Toxin-Antitoxin Systems. Front. Microbiol. 2017, 8, 1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; Di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the host immune system and bacteriophage Is essential for successful phage therapy against an acute respiratory pathogen. Cell Host Microbe 2017, 22, 38–47. [Google Scholar] [CrossRef]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.-A. Synergistic interaction between phage therapy and antibiotics clears Pseudomonas Aeruginosa infection in endocarditis and reduces virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez, C.A.; Koskella, B. Phage resistance evolution in vitro is not reflective of in vivo outcome in a plant-bacteria-phage system. Evol. Int. J. Org. Evol. 2019, 73, 2461–2475. [Google Scholar] [CrossRef]
- Huang, T.; Zheng, Y.; Yan, Y.; Yang, L.; Yao, Y.; Zheng, J.; Wu, L.; Wang, X.; Chen, Y.; Xing, J.; et al. Probing minority population of antibiotic-resistant bacteria. Biosens. Bioelectron. 2016, 80, 323–330. [Google Scholar] [CrossRef]
- Davis, K.M.; Isberg, R.R. Defining heterogeneity within bacterial populations via single cell approaches. BioEssays 2016, 38, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.E.; Otoupal, P.B.; Chatterjee, A. Transcriptome-level signatures in gene expression and gene expression variability during bacterial adaptive evolution. mSphere 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Latz, S.; Krüttgen, A.; Häfner, H.; Buhl, E.; Ritter, K.; Horz, H.-P. Differential effect of newly isolated phages belonging to PB1-like, phiKZ-like and LUZ24-like viruses against multi-drug resistant Pseudomonas aeruginosa under varying growth conditions. Viruses 2017, 9, 315. [Google Scholar] [CrossRef] [Green Version]
- Burmeister, A.R.; Fortier, A.; Roush, C.; Lessing, A.J.; Bender, R.G.; Barahman, R.; Grant, R.; Chan, B.K.; Turner, P.E. Pleiotropy complicates a trade-off between phage resistance and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 11207–11216. [Google Scholar] [CrossRef]
- Xie, Y.; Wahab, L.; Gill, J. Development and validation of a microtiter plate-based assay for determination of bacteriophage host range and virulence. Viruses 2018, 10, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 70, pp. 217–248. ISBN 9780123809919. [Google Scholar]
- Roberts, A.E.L.; Kragh, K.N.; Bjarnsholt, T.; Diggle, S.P. The limitations of in vitro experimentation in understanding biofilms and chronic infection. J. Mol. Biol. 2015, 427, 3646–3661. [Google Scholar] [CrossRef]
- Wang, S.; Yin, B.; Yu, L.; Dang, M.; Guo, Z.; Yan, G.; Hu, D.; Gu, J.; Du, C.; Feng, X.; et al. Overexpression of AmpC promotes bacteriophage lysis of ampicillin-resistant Escherichia coli. Front. Microbiol. 2020, 10, 2973. [Google Scholar] [CrossRef] [Green Version]
- David, M.D.; Weller, T.M.A.; Lambert, P.; Fraise, A.P. An outbreak of Serratia marcescens on the neonatal unit: A tale of two clones. J. Hosp. Infect. 2006, 63, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Herbert, S.; Halvorsen, D.S.; Leong, T.; Franklin, C.; Harrington, G.; Spelman, D. Large outbreak of infection and colonization with gram-negative pathogens carrying the metallo-β-Lactamase Gene bla IMP-4 at a 320-bed tertiary hospital in Australia. Infect. Control. Hosp. Epidemiol. 2007, 28, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Nastro, M.; Monge, R.; Zintgraff, J.; Vaulet, L.G.; Boutureira, M.; Famiglietti, A.; Rodriguez, C.H. First nosocomial outbreak of VIM-16-producing Serratia marcescens in Argentina. Clin. Microbiol. Infect. 2013, 19, 617–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachino, J.; Yoshida, H.; Yamane, K.; Suzuki, S.; Matsui, M.; Yamagishi, T.; Tsutsui, A.; Konda, T.; Shibayama, K.; Arakawa, Y. SMB-1, a Novel subclass B3 metallo-β-lactamase, associated with IS CR1 and a class 1 integron, from a carbapenem-resistant Serratia marcescens clinical isolate. Antimicrob. Agents Chemother. 2011, 55, 5143–5149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutter, E. Phage host range and efficiency of plating. In Bacteriophages; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; Volume 501, pp. 141–149. ISBN 978-1-60327-164-6. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [Green Version]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Related Phage | Query Cover | Identity | Accession-No. |
|---|---|---|---|
| Serratia phage SM9-3Y | 95% | 96.4% | KX778611.3 |
| Escherichia phage LL2 | 88% | 96.2% | MH717709.1 |
| Leclercia phage 10164-302 | 89% | 96.6% | MF285616.1 |
| Yersinia phage phiYeO3-12 | 90% | 96.3% | AJ251805.1 |
| Salmonella phage phiSG-JL2 | 87% | 96.3% | EU547803.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, L.; Jansen, M.; Krüttgen, A.; Buhl, E.M.; Horz, H.-P. Tackling Intrinsic Antibiotic Resistance in Serratia marcescens with a Combination of Ampicillin/Sulbactam and Phage SALSA. Antibiotics 2020, 9, 371. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070371
Weber L, Jansen M, Krüttgen A, Buhl EM, Horz H-P. Tackling Intrinsic Antibiotic Resistance in Serratia marcescens with a Combination of Ampicillin/Sulbactam and Phage SALSA. Antibiotics. 2020; 9(7):371. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070371
Chicago/Turabian StyleWeber, Lorenz, Mathias Jansen, Alex Krüttgen, Eva Miriam Buhl, and Hans-Peter Horz. 2020. "Tackling Intrinsic Antibiotic Resistance in Serratia marcescens with a Combination of Ampicillin/Sulbactam and Phage SALSA" Antibiotics 9, no. 7: 371. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070371