Ethyl Acetate Fraction of Helianthus tuberosus L. Induces Anti-Diabetic, and Wound-Healing Activities in Insulin-Resistant Human Liver Cancer and Mouse Fibroblast Cells


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Cell Lines
2.2. Preparation of Plant Extract and their Active Fractions
2.3. Estimation of Total Phenol and Flavonoid Content
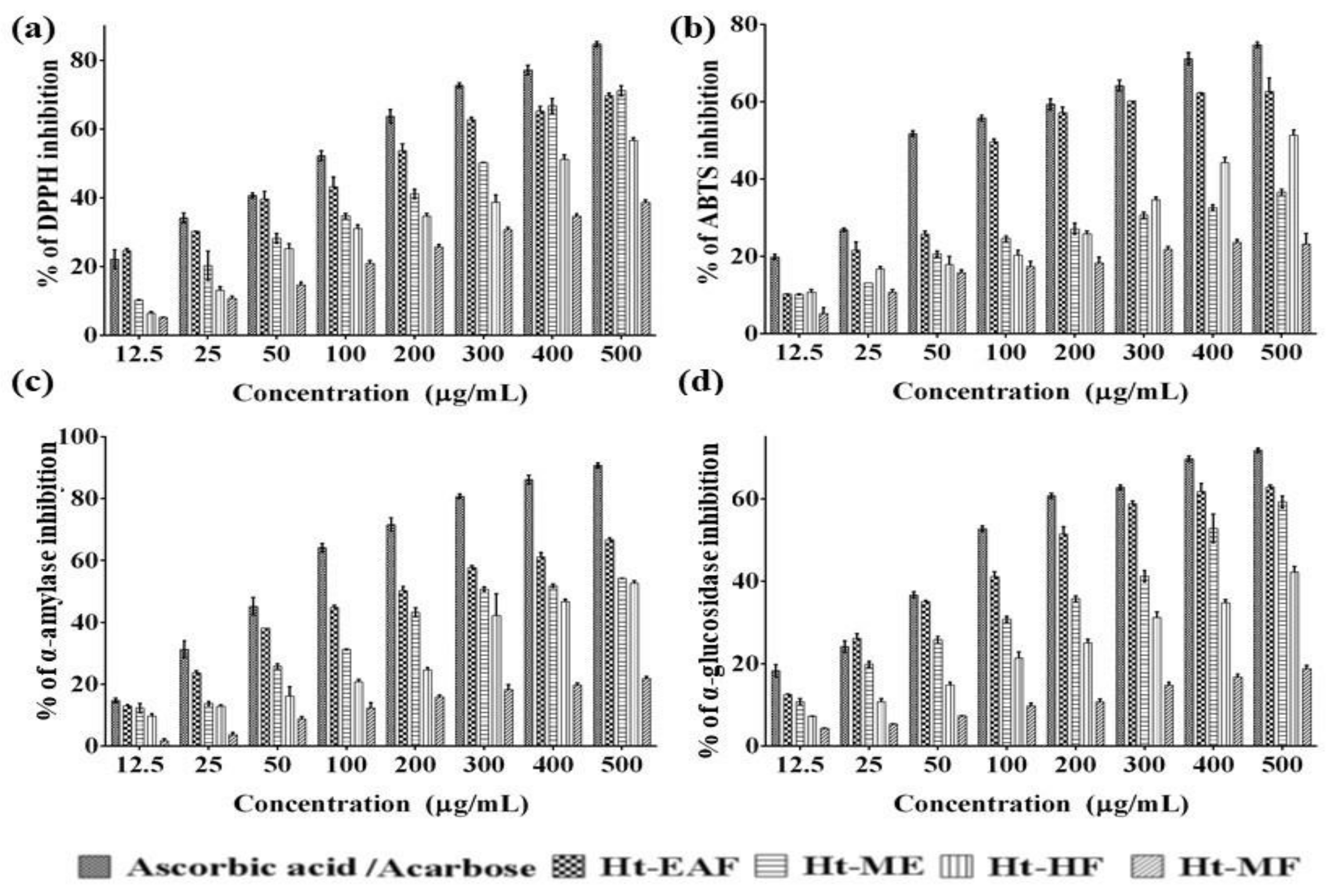
2.4. Determination of In Vitro Antioxidant Activity
2.5. α-Amylase and α-Glucosidase Inhibition Assay
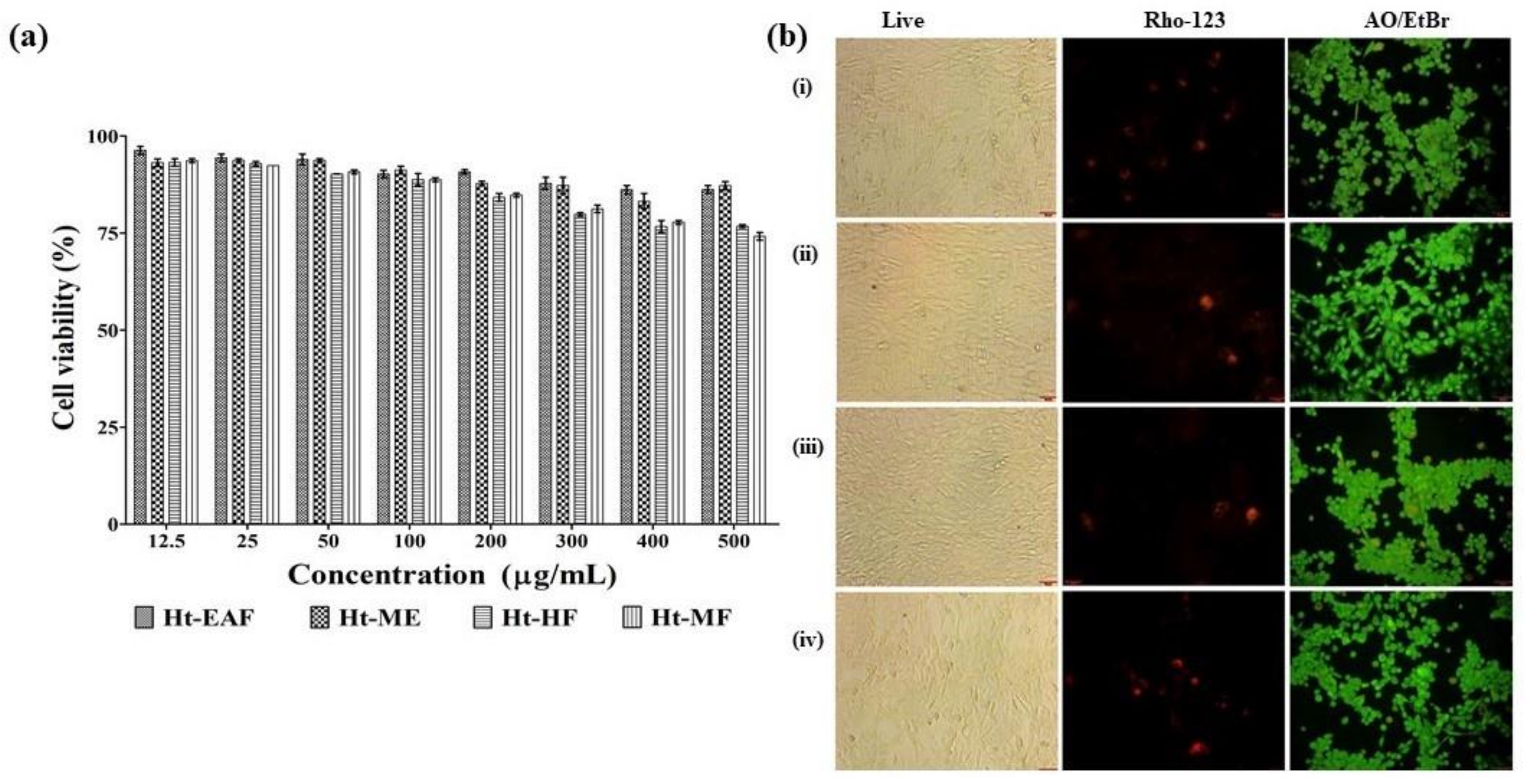
2.6. Biocompatible Nature of Ethyl Acetate Fraction of H. tuberosus
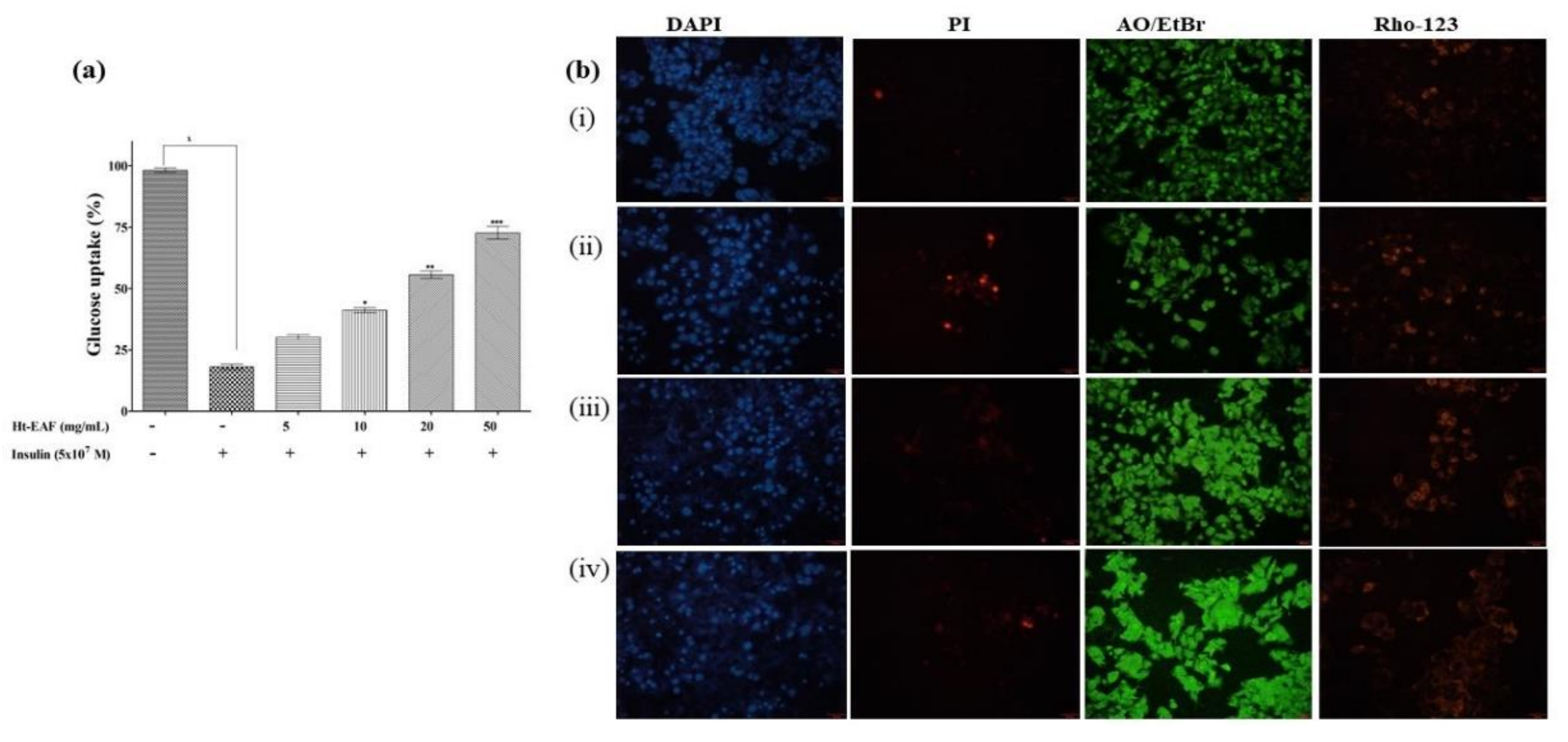
2.7. In Vitro Anti-Diabetic Nature of Ethyl Acetate Fraction of H. tuberosus
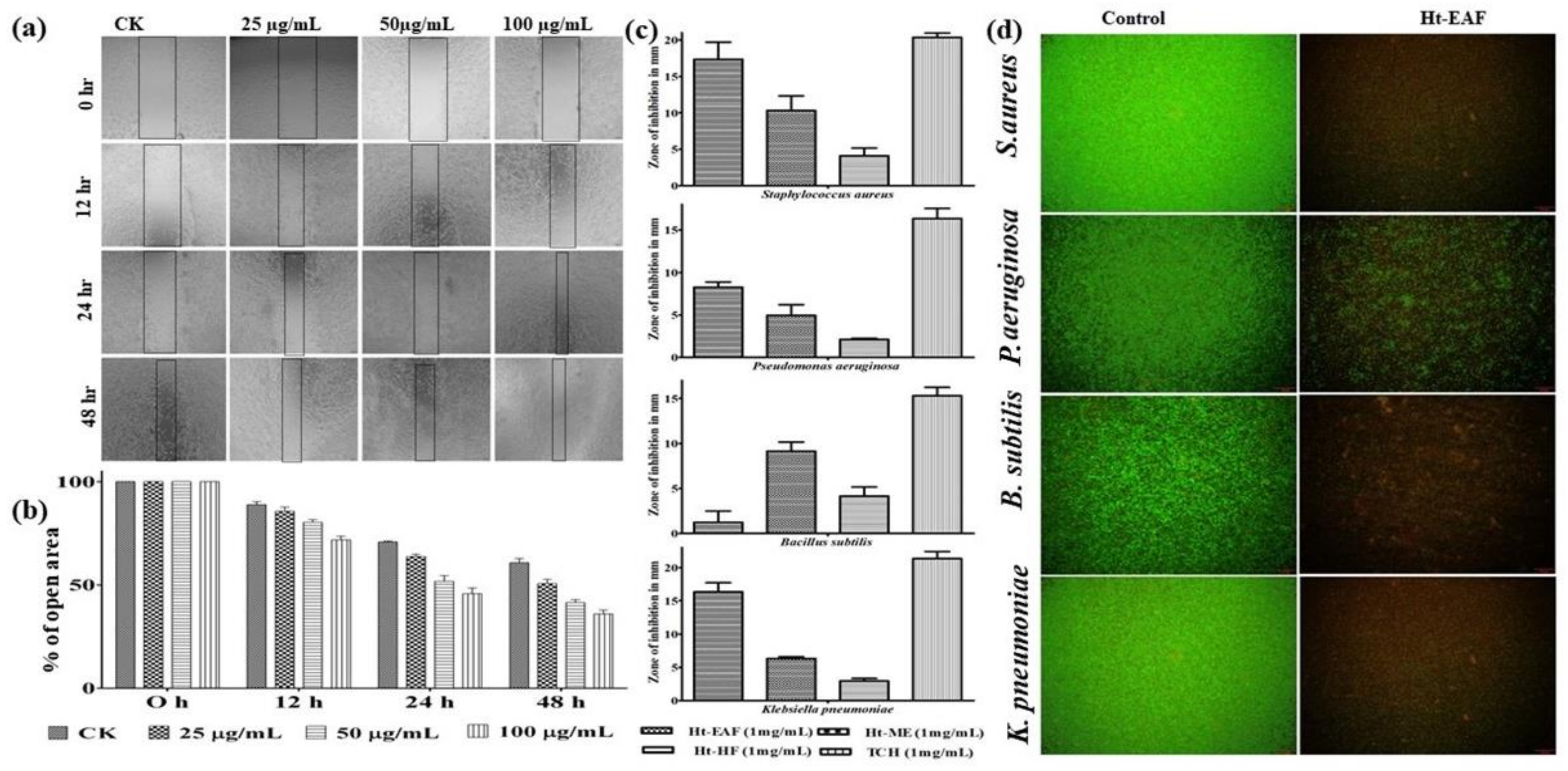
2.8. In Vitro Wound Scratch Assay
2.9. Anti-Bacterial Activity
2.10. UPLC-QTOF-MS/MS Analysis
2.11. GC-MS Analysis
2.12. Molecular Docking Study
2.13. Statistical Analysis
3. Results and Discussion
3.1. Phytochemical Quantification
3.2. Radical Scavenging Potential
3.3. In Vitro Anti-Diabetic Efficacy
3.4. Biocompatibility Nature of H. tuberosus
3.5. Anti-Diabetic Effects of H. tuberosus
3.6. Wound-Healing Ability in Fibroblast Cells
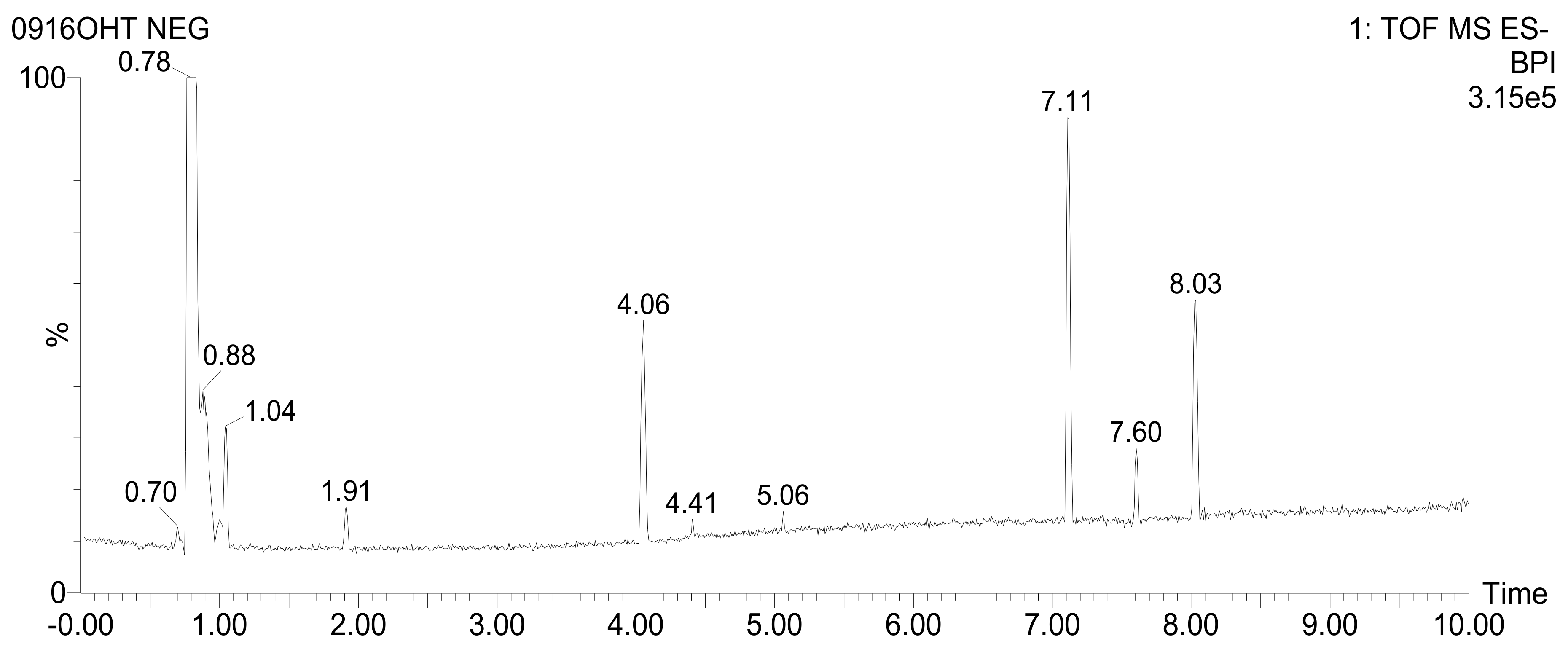
3.7. UPLC-QTOF-MS/MS Analysis
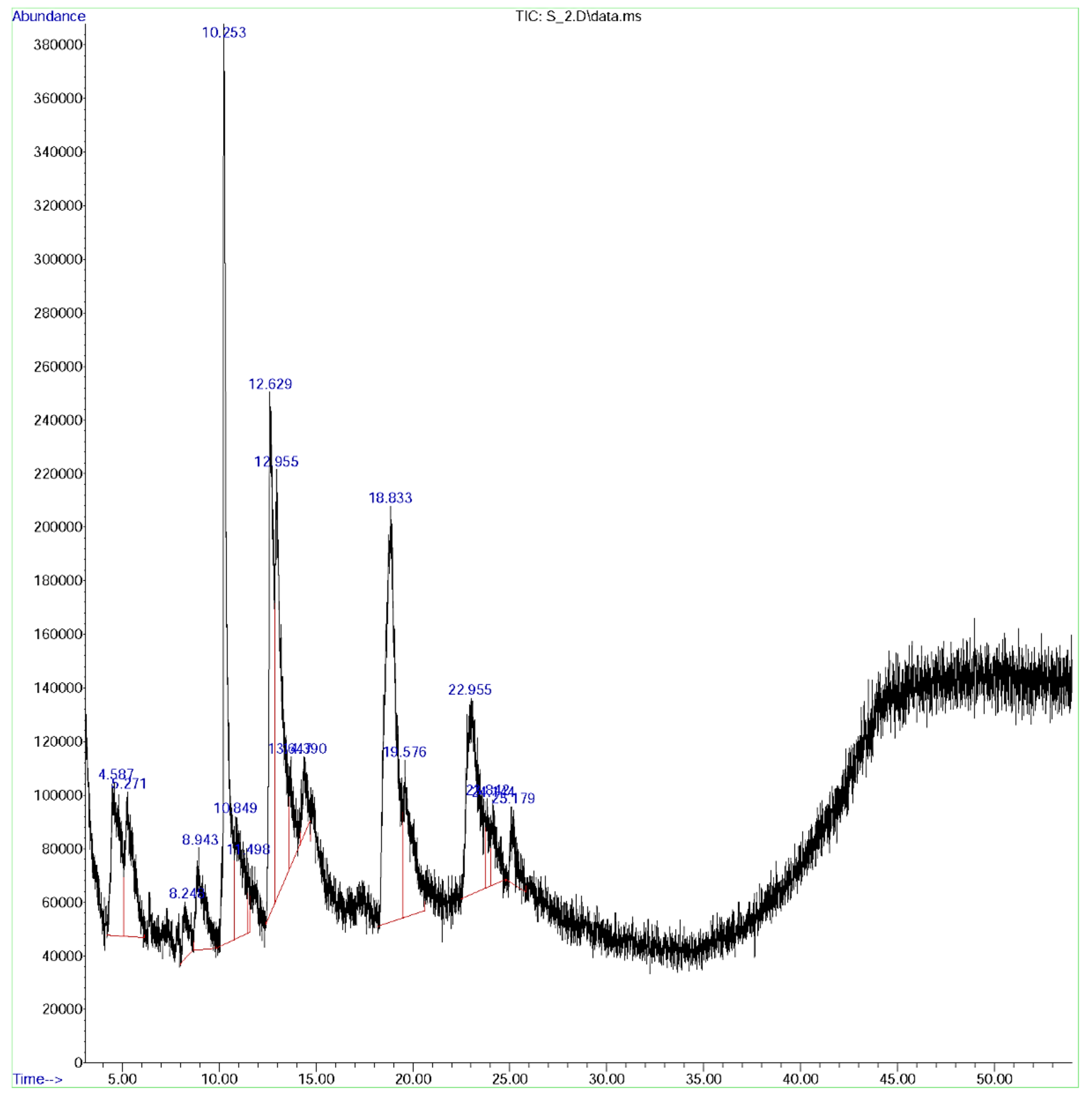
3.8. GC-MS Analysis
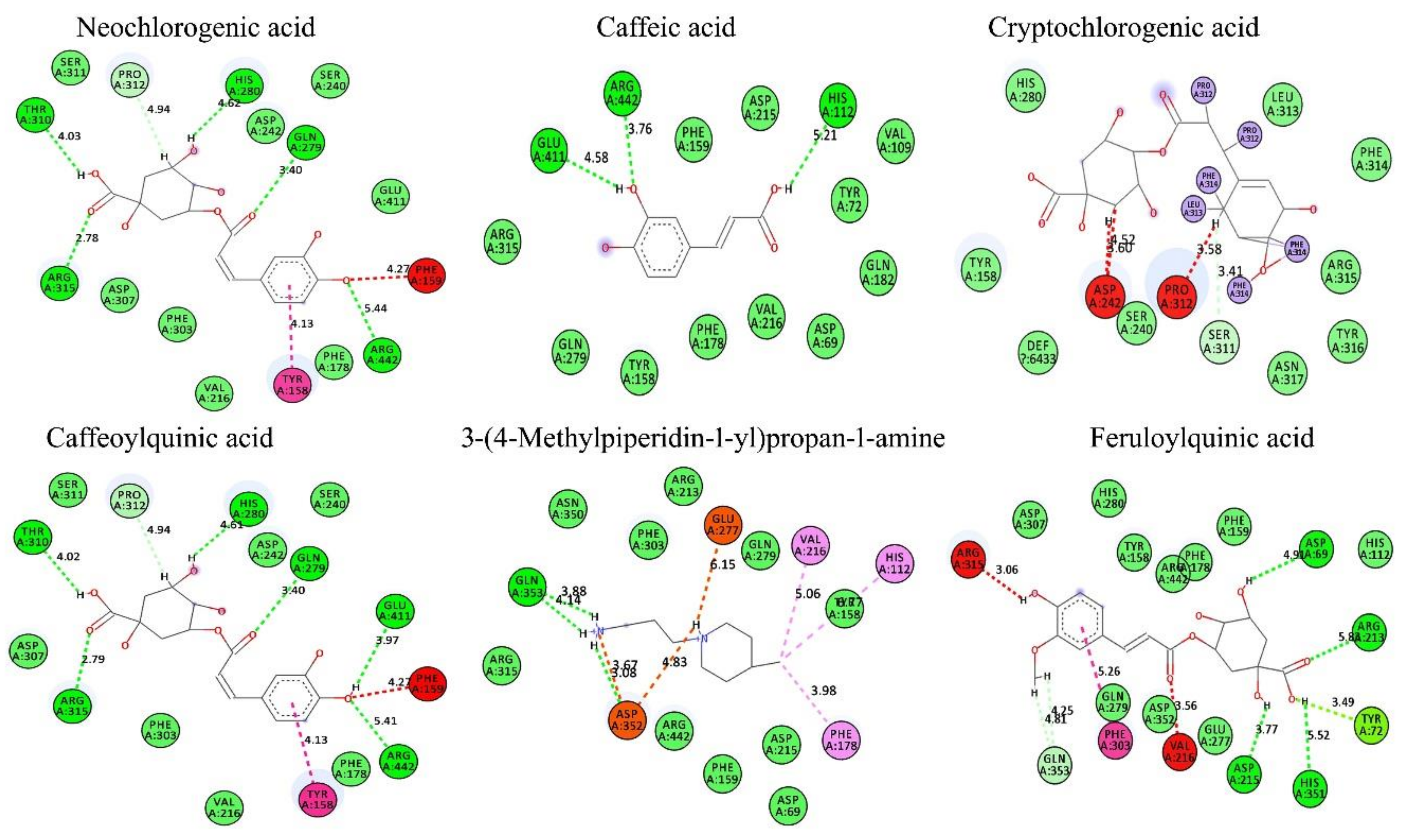
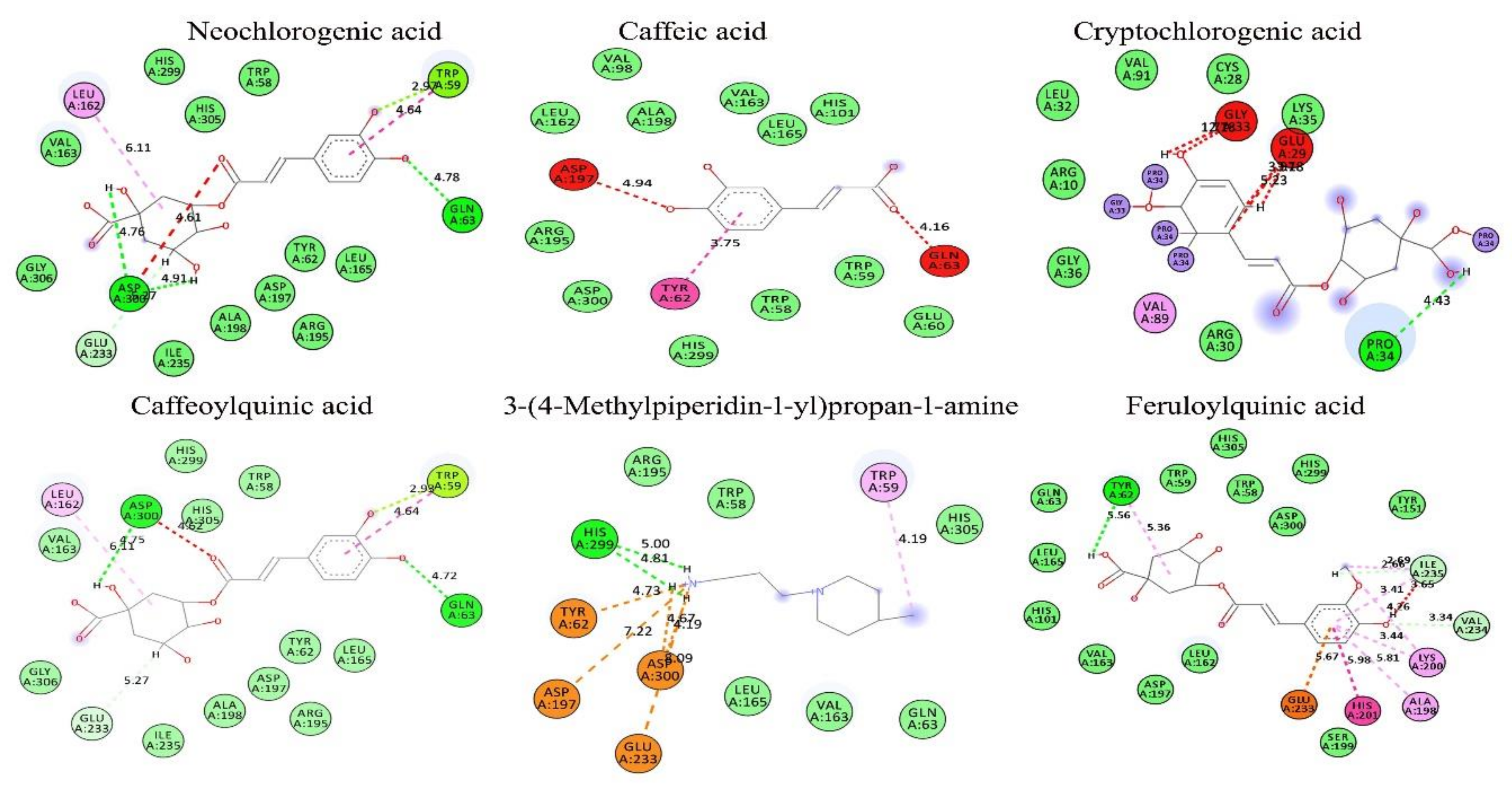
3.9. In Silico Conformation of Anti-Diabetic Efficacy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Freeman, A. Fast Food: Oppression Through Poor Nutrition. California Law Rev. 2007, 95, 2221. [Google Scholar] [CrossRef]
- Forouhi, N.G.; Misra, A.; Mohan, V.; Taylor, R.; Yancy, W. Dietary and nutritional approaches for prevention and management of type 2 diabetes. BMJ 2018, 361, k2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvis, N. Nutritional recommendations for individuals with diabetes mellitus. S. Afr. Med. J. 1992, 81, 162–166. [Google Scholar] [PubMed]
- Bethel, M.A.; Engel, S.S.; Stevens, S.R.; Lokhnygina, Y.; Ding, J.; Josse, R.G.; Alvarsson, M.; Hramiak, I.; Green, J.B.; Peterson, E.D.; et al. Progression of glucose-lowering diabetes therapy in TECOS. Endocrinol. Diabetes Metab. 2019, 2, e00053. [Google Scholar] [CrossRef] [Green Version]
- Patwardhan, B.; Warude, D.; Pushpangadan, P.; Bhatt, N. Ayurveda and traditional Chinese medicine: A comparative overview. Evid Based Complementary Altern. Med. 2005, 2, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Abima Shazhni, J.R.; Renu, A.; Vijayaraghavan, P. Insights of antidiabetic, anti-inflammatory and hepatoprotective properties of antimicrobial secondary metabolites of corm extract from Caladium × hortulanum. Saudi J. Biol. Sci. 2018, 25, 1755–1761. [Google Scholar] [CrossRef]
- Moradabadi, L.; Montasser Kouhsari, S.; Fehresti Sani, M. Hypoglycemic effects of three medicinal plants in experimental diabetes: Inhibition of rat intestinal α-glucosidase and enhanced pancreatic insulin and cardiac glut-4 mrnas expression. Iran. J. Pharm. Res. IJPR 2013, 12, 387–397. [Google Scholar]
- Patel, D.K.; Prasad, S.K.; Kumar, R.; Hemalatha, S. An overview on antidiabetic medicinal plants having insulin mimetic property. Asian Pac. J. Trop. Biomed. 2012, 2, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Sun, X.; Jiang, X.; Wang, L.; Tian, J.; Li, L.; Zhao, M.; Zhong, Q. Characterization of the Tibet plateau Jerusalem artichoke (Helianthus tuberosus L.) transcriptome by de novo assembly to discover genes associated with fructan synthesis and SSR analysis. Hereditas 2019, 156, 9. [Google Scholar] [CrossRef]
- Matsuura, H.; Yoshihara, T.; Ichihara, A.; Kikuta, Y.; Koda, Y. Tuber-forming substances in jerusalem artichoke (Helianthus tuberosus L.). Biosci. Biotechnol. Biochem. 1993, 57, 1253–1256. [Google Scholar] [CrossRef] [Green Version]
- Gupta, D.; Chaturvedi, N. Impact of Processing on Inulin and sugars content of Jerusalem Artichoke Tuber. Res. J. Pharm. Technol. 2020, 13, 3143–3146. [Google Scholar] [CrossRef]
- Pan, L.; Sinden, M.R.; Kennedy, A.H.; Chai, H.; Watson, L.E.; Graham, T.L.; Kinghorn, A.D. Bioactive constituents of Helianthus tuberosus (Jerusalem artichoke). Phytochem. Lett. 2009, 2, 15–18. [Google Scholar] [CrossRef]
- Aslan, M.; Orhan, N.; Orhan, D.D.; d Ergun, F. Hypoglycemic activity and antioxidant potential of some medicinal plants traditionally used in Turkey for diabetes. J. Ethnopharmacol. 2010, 128, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Gengaihi, S.; Aboul-Enein, A.; Abou Baker, D. Molecular characterizations and antimicrobial activities of chicory and jerusalem artichoke plants. Int. J. Acad. Res. 2009, 1, 66–71. [Google Scholar]
- Teke, G.N.; Lunga, P.K.; Wabo, H.K.; Kuiate, J.-R.; Vilarem, G.; Giacinti, G.; Kikuchi, H.; Oshima, Y. Antimicrobial and antioxidant properties of methanol extract, fractions and compounds from the stem bark of Entada abyssinica Stend ex A. Satabie. BMC Complementary Altern. Med. 2011, 11, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, N.; Khan, M.R.; Shabbir, M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complementary Altern. Med. 2012, 12, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnao, M.B.; Cano, A.; Acosta, M. The hydrophilic and lipophilic contribution to total antioxidant activity. Food Chem. 2001, 73, 239–244. [Google Scholar] [CrossRef]
- Dehghan, H.; Salehi, P.; Amiri, M.S. Bioassay-guided purification of α-amylase, α-glucosidase inhibitors and DPPH radical scavengers from roots of Rheum turkestanicum. Ind. Crop. Prod. 2018, 117, 303–309. [Google Scholar] [CrossRef]
- Sathiyaseelan, A.; Saravanakumar, K.; Mariadoss, A.V.A.; Wang, M.-H. Biocompatible fungal chitosan encapsulated phytogenic silver nanoparticles enhanced antidiabetic, antioxidant and antibacterial activity. Int. J. Biol. Macromol. 2020, 153, 63–71. [Google Scholar] [CrossRef]
- Mariadoss, A.V.A.; Saravanakumar, K.; Sathiyaseelan, A.; Wang, M.-H. Preparation, characterization and anti-cancer activity of graphene oxide--silver nanocomposite. J. Photochem. Photobiol. B Biol. 2020, 210, 111984. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Sathiyaseelan, A.; Mariadoss, A.V.A.; Jeevithan, E.; Hu, X.; Shin, S.; Wang, M.-H. Dual stimuli-responsive release of aptamer AS1411 decorated erlotinib loaded chitosan nanoparticles for non-small-cell lung carcinoma therapy. Carbohydr. Polym. 2020, 245, 116407. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Teng, H.; Cao, H. Chlorogenic acid and caffeic acid from Sonchus oleraceus Linn synergistically attenuate insulin resistance and modulate glucose uptake in HepG2 cells. Food Chem. Toxicol. 2019, 127, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Sathiyaseelan, A.; Anand Mariadoss, A.V.; Wang, M.-H. Antioxidant and antidiabetic properties of biocompatible ceria oxide (CeO2) nanoparticles in mouse fibroblast NIH3T3 and insulin resistant HepG2 cells. Ceram. Int. 2020, in press. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Mariadoss, A.V.A.; Sathiyaseelan, A.; Wang, M.-H. Synthesis and characterization of nano-chitosan capped gold nanoparticles with multifunctional bioactive properties. Int. J. Biol. Macromol. 2020, 165, 747–757. [Google Scholar] [CrossRef]
- Mariadoss, A.V.A.; Saravanakumar, K.; Sathiyaseelan, A.; Venkatachalam, K.; Wang, M.-H. Folic acid functionalized starch encapsulated green synthesized copper oxide nanoparticles for targeted drug delivery in breast cancer therapy. Int. J. Biol. Macromol. 2020, 164, 2073–2084. [Google Scholar] [CrossRef] [PubMed]
- Sathiyaseelan, A.; Saravanakumar, K.; Mariadoss, A.V.; Ramachandran, C.; Hu, X.; Oh, D.H.; Wang, M.H. Chitosan-tea tree oil nanoemulsion and calcium chloride tailored edible coating increase the shelf life of fresh cut red bell pepper. Prog. Org. Coat. 2020, 106010. [Google Scholar] [CrossRef]
- Mariadoss, A.V.A.; Vinyagam, R.; Rajamanickam, V.; Sankaran, V.; Venkatesan, S.; David, E. Pharmacological aspects and potential use of phloretin: A systemic review. Mini Rev. Med. Chem. 2019, 19, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.J.; Long, X.H.; Li, E.Z. Evaluation of antifungal phenolics from helianthus tuberosus l. leaves against phytophthora capsici leonian by chemometric analysis. Molecules 2019, 24, 4300. [Google Scholar] [CrossRef] [Green Version]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: A cellular mechanism review. Nutr. Metab. 2015, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Nizioł-Łukaszewska, Z.; Furman-Toczek, D.; Zagórska-Dziok, M. Antioxidant activity and cytotoxicity of Jerusalem artichoke tubers and leaves extract on HaCaT and BJ fibroblast cells. Lipids Health Dis. 2018, 17, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suluvoy, J.K.; Berlin Grace, V.M. Phytochemical profile and free radical nitric oxide (NO) scavenging activity of Averrhoa bilimbi L. fruit extract. 3 Biotechnology 2017, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complementary Med. 2017, 8, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Kooti, W.; Farokhipour, M.; Asadzadeh, Z.; Ashtary-Larky, D.; Asadi-Samani, M. The role of medicinal plants in the treatment of diabetes: A systematic review. Electron. Physician 2016, 8, 1832–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, R.; Kayani, W.K.; Ahmed, T.; Malik, F.; Hussain, S.; Ashfaq, M.; Ali, H.; Rubnawaz, S.; Green, B.D.; Calderwood, D.; et al. Assessment of antidiabetic potential and phytochemical profiling of Rhazya stricta root extracts. BMC Complement Med. Ther. 2020, 20, 293. [Google Scholar] [CrossRef] [PubMed]
- Jdir, H.; Khemakham, B.; Chakroun, M.; Zouari, S.; Ali, Y.B.; Zouari, N. Diplotaxis simplex suppresses postprandial hyperglycemia in mice by inhibiting key-enzymes linked to type 2 diabetes. Rev. Bras. Farmacogn. 2015, 25, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Sathish, G.; Jayanthi, M.; Muthukumaran, J.; Muruganathan, U.; Ramachandran, V. Ameliorating effect of eugenol on hyperglycemia by attenuating the key enzymes of glucose metabolism in streptozotocin-induced diabetic rats. Mol. Cell. Biochem. 2014, 385, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.B.; Alimova, Y.; Myers, T.M.; Ebersole, J.L. Short- and medium-chain fatty acids exhibit antimicrobial activity for oral microorganisms. Arch. Oral Biol. 2011, 56, 650–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, H.Y.; Kim, M.; Seo, C.R.; Yoo, H.J.; Lee, S.-H.; Lee, J.H. The effects of Jerusalem artichoke and fermented soybean powder mixture supplementation on blood glucose and oxidative stress in subjects with prediabetes or newly diagnosed type 2 diabetes. Nutr. Diabetes 2018, 8, 42. [Google Scholar] [CrossRef]
- Addis, R.; Cruciani, S.; Santaniello, S.; Bellu, E.; Sarais, G.; Ventura, C.; Maioli, M.; Pintore, G. Fibroblast Proliferation and Migration in Wound Healing by Phytochemicals: Evidence for a Novel Synergic Outcome. Int. J. Med. Sci. 2020, 17, 1030–1042. [Google Scholar] [CrossRef] [Green Version]
- Nouvong, A.; Ambrus, A.M.; Zhang, E.R.; Hultman, L.; Coller, H.A. Reactive oxygen species and bacterial biofilms in diabetic wound healing. Physiol. Genom. 2016, 48, 889–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogba, O.M.; Nsan, E.; Eyam, E.S. Aerobic bacteria associated with diabetic foot ulcers and their susceptibility pattern. Biomed. Dermatol. 2019, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Baharfar, R.; Azimi, R.; Mohseni, M. Antioxidant and antibacterial activity of flavonoid-, polyphenol- and anthocyanin-rich extracts from Thymus kotschyanus boiss & hohen aerial parts. J. Food Sci. Technol. 2015, 52, 6777–6783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariadoss, A.V.A.; Ramachandran, V.; Shalini, V.; Agilan, B.; Franklin, J.H.; Sanjay, K.; Alaa, Y.G.; Tawfiq, M.A.-A.; Ernest, D. Green synthesis, characterization and antibacterial activity of silver nanoparticles by Malus domestica and its cytotoxic effect on (MCF-7) cell line. Microb. Pathog. 2019, 135, 103609. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Long, X.; Liu, Z.; Shao, H.; Liu, L. Analysis of phenolic acids of Jerusalem artichoke (Helianthus tuberosus L.) responding to salt-stress by liquid chromatography/tandem mass spectrometry. Sci. World J. 2014, 2014, 568043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virgen-Ortíz, J.J.; Ibarra-Junquera, V.; Escalante-Minakata, P.; Centeno-Leija, S.; Serrano-Posada, H.; de Jesús Ornelas-Paz, J.; Pérez-Martínez, J.D.; Osuna-Castro, J.A. Identification and functional characterization of a fructooligosaccharides-forming enzyme from aspergillus aculeatus. Appl. Biochem. Biotechnol. 2016, 179, 497–513. [Google Scholar] [CrossRef] [PubMed]
- Kaszás, L.; Alshaal, T.; El-Ramady, H.; Kovács, Z.; Koroknai, J.; Elhawat, N.; Nagy, É.; Cziáky, Z.; Fári, M.; Domokos-Szabolcsy, É. Identification of Bioactive Phytochemicals in Leaf Protein Concentrate of Jerusalem Artichoke (Helianthus tuberosus L.). Plants 2020, 9, 889. [Google Scholar] [CrossRef]
- Piraud, M.; Vianey-Saban, C.; Petritis, K.; Elfakir, C.; Steghens, J.P.; Morla, A.; Bouchu, D. ESI-MS/MS analysis of underivatised amino acids: A new tool for the diagnosis of inherited disorders of amino acid metabolism. Fragmentation study of 79 molecules of biological interest in positive and negative ionisation mode. Rapid Commun. Mass Spectrom. RCM 2003, 17, 1297–1311. [Google Scholar] [CrossRef]
- Bianco, A.; Buiarelli, F.; Cartoni, G.; Coccioli, F.; Muzzalupo, I.; Polidori, A.; Uccella, N. Analysis by hplc-ms/ms of biophenolic components in olives and oils. Anal. Lett. 2001, 34, 1033–1051. [Google Scholar] [CrossRef]
- Zhang, L.; Tu, Z.-C.; Xie, X.; Wang, H.; Wang, H.; Wang, Z.-X.; Sha, X.-M.; Lu, Y. Jackfruit (Artocarpus heterophyllus Lam.) peel: A better source of antioxidants and a-glucosidase inhibitors than pulp, flake and seed, and phytochemical profile by HPLC-QTOF-MS/MS. Food Chem. 2017, 234, 303–313. [Google Scholar] [CrossRef]
- Ruan, J.; Yan, J.; Zheng, D.; Sun, F.; Wang, J.; Han, L.; Zhang, Y.; Wang, T. Comprehensive chemical profiling in the ethanol extract of pluchea indica aerial parts by liquid chromatography/mass spectrometry analysis of its silica gel column chromatography fractions. Molecules 2019, 24, 2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhammad, N.; Hejazi, V.; Abbas, T.; Ali, A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Kumawat, V.S.; Kaur, G. Insulinotropic and antidiabetic effects of β-caryophyllene with l-arginine in type 2 diabetic rats. J. Food Biochem. 2020, 44, e13156. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-L.; Peng, B.-J.; Zhong, Y.-L.; Liu, Y.-L.; Song, Z.; Wang, Z. Effect of 5-caffeoylquinic acid on the NF-κB signaling pathway, peroxisome proliferator-activated receptor gamma 2, and macrophage infiltration in high-fat diet-fed Sprague–Dawley rat adipose tissue. Food Funct. 2015, 6, 2779–2786. [Google Scholar] [CrossRef] [PubMed]
- Berthet, M.; Cheviet, T.; Dujardin, G.; Parrot, I.; Martinez, J. Isoxazolidine: A privileged scaffold for organic and medicinal chemistry. Chem. Rev. 2016, 116, 15235–15283. [Google Scholar] [CrossRef]
- Zaki, M.; Oukhrib, A.; Akssira, M.; Berteina-Raboin, S. Synthesis of novel spiro-isoxazoline and spiro-isoxazolidine derivatives of tomentosin. RSC Adv. 2017, 7, 6523–6529. [Google Scholar] [CrossRef] [Green Version]
- Mosbah, H.; Chahdoura, H.; Mannai, A.; Snoussi, M.; Aouadi, K.; Abreu, R.M.V.; Bouslama, A.; Achour, L.; Selmi, B. Biological activities evaluation of enantiopure isoxazolidine derivatives: In vitro, in vivo and in silico studies. Appl. Biochem. Biotechnol. 2019, 187, 1113–1130. [Google Scholar] [CrossRef] [Green Version]
- Hall, I.H.; Wong, O.T.; Simlot, R.; Izydore, R.A. Effects of isoxazolidine or triazolidine on rat serum lipids in vivo and LDL and HDL binding and degradation in human and rodent cultured cells in vitro. Res. Commun Chem. Pathol. Pharmacol. 1992, 77, 327–346. [Google Scholar]
- Arslan, S.; Loğoğlu, E.; Oktemer, A. Antimicrobial activity studies on some piperidine and pyrrolidine substituted halogenobenzene derivatives. J. Enzym. Inhib. Med. Chem. 2006, 21, 211–214. [Google Scholar] [CrossRef]
- Buko, V.; Zavodnik, I.; Kanuka, O.; Belonovskaya, E.; Naruta, E.; Lukivskaya, O.; Kirko, S.; Budryn, G.; Żyżelewicz, D.; Oracz, J.; et al. Antidiabetic effects and erythrocyte stabilization by red cabbage extract in streptozotocin-treated rats. Food Funct. 2018, 9, 1850–1863. [Google Scholar] [CrossRef]
- Ghosh, K.; Shetty, S.; Jijina, F.; Mohanty, D. Role of epsilon amino caproic acid in the management of haemophilic patients with inhibitors. Haemoph. J. World Fed. Hemoph. 2004, 10, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Heerding, D.A.; Rhodes, N.; Leber, J.D.; Clark, T.J.; Keenan, R.M.; Lafrance, L.V.; Li, M.; Safonov, I.G.; Takata, D.T.; Venslavsky, J.W.; et al. Identification of 4-(2-(4-Amino-1,2,5-oxadiazol-3-yl)-1-ethyl-7-{[(3S)-3-piperidinylmethyl]oxy}-1H-imidazo[4,5-c]pyridin-4-yl)-2-methyl-3-butyn-2-ol (GSK690693), a Novel Inhibitor of AKT Kinase. J. Med. Chem. 2008, 51, 5663–5679. [Google Scholar] [CrossRef] [PubMed]
- Hanaya, T.; Itoh, C.J.C. An efficient synthesis of antibiotic sf 2312 (3 Dihydroxyphosphoryl-1,5 dihydrox™ 2 pyrrolidone). Heterocycles 2011, 42, 1675–1683. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.F.; Lemos, T.G.; de Mattos, M.C.; Segundo, T.A.; Santiago, G.M.; Braz-Filho, R. New enamine derivatives of lapachol and biological activity. An. Acad. Bras. Cienc. 2002, 74, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.K.; Nam, P.-W.; Lee, S.-J.; Lee, K.-G. Antioxidant activities of volatile and non-volatile fractions of selected traditionally brewed Korean rice wines. J. Inst. Brew. 2014, 120, 537–542. [Google Scholar] [CrossRef]
- Carlotto, J.; da Silva, L.M.; Dartora, N.; Maria-Ferreira, D.; Sabry Dde, A.; Filho, A.P.; de Paula Werner, M.F.; Sassaki, G.L.; Gorin, P.A.; Iacomini, M.; et al. Identification of a dicaffeoylquinic acid isomer from Arctium lappa with a potent anti-ulcer activity. Talanta 2015, 135, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, I.; Peroulis, N.; Pantazis, P.; Hadzopoulou-Cladaras, M. Camphene, a plant-derived monoterpene, reduces plasma cholesterol and triglycerides in hyperlipidemic rats independently of hmg-coa reductase activity. PLoS ONE 2011, 6, e20516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Kumar, R.; Pant, A.; Kumar, M.; Isidorov, V.; Szczepaniak, L. Chemical composition, anti-inflammatory, analgesic, antipyretic, myorelaxant, antibacterial and antifungal activity of rabdosia rugosus wall. (Syn. Plectranthus rugosus Wall.). J. Med. Herbs Ethnomed. 2019, 5, 8–15. [Google Scholar] [CrossRef]
- Montes-Avila, J.; Sarmiento Sanchez, J.I.; Delgado-Vargas, F.; Rivero, I.; Diaz-Camacho, S.; Uribe-Beltrán, M. Antioxidant activity and antimicrobial evaluation of 1-benzyl-1,2,3-triazole. Acta Univ. 2016, 26, 63–67. [Google Scholar] [CrossRef]
- Lipinski, C.A. Drug-like properties and the causes of poor solubility and poor permeability. J. Pharmacol. Toxicol. Methods 2000, 44, 235–249. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Phenolic Content (mg GAE/g of Extract) | Total Flavonoid Content (mg QE/g of Extract) | |
|---|---|---|
| Ht-EAF | 69.55 ± 0.36 | 21.03 ± 0.97 |
| Ht-HF | 17.11 ± 1.05 | 11.48 ± 0.11 |
| Ht-MF | 10.29 ± 0.82 | 6.27 ± 1.09 |
| Ht-ME | 43.07 ± 0.05 | 19.85 ± 0.21 |
| RT (min) | Tentative Identification | Formula | m/z [M-H]- | Mass Error (ppm) | Response | Fragmentation (m/z) | Reference |
|---|---|---|---|---|---|---|---|
| 0.79 | Sucrose | C12H22O11 | 341.1095 | 1.9 | 3592134 | 179.0563 | [47] |
| 1.04 | Fructosylnystose | C30H52O26 | 827.2669 | −0.3 | 2910 | 179.0560, 341.1090, 503.1615, 665.2151 | [47] |
| 1.07 | Salicylic acid β-D-glucoside | C13H16O8 | 299.0772 | −0.1 | 315 | 137.0246, 180.0653 | [48] |
| 1.30 | Phenylalanine | C9H11NO2 | 164.0717 | −0.3 | 15377 | 147.0454 | [49] |
| 1.30 | Cinnamic acid | C9H8O2 | 147.0457 | 3.5 | 2185 | - | [50] |
| 1.50 | Neochlorogenic acid | C16H18O9 | 353.0879 | 0.2 | 4678 | 179.0345, 191.0557 | [48] |
| 1.75 | Tryptophan | C11H12N2O2 | 203.0823 | −1.5 | 4696 | 116.0516 | [51] |
| 1.91 | Chlorogenic acid | C16H18O9 | 353.0876 | −0.7 | 45770 | 135.0454, 179.0348, 191.0557 | [48] |
| 2.37 | Caffeic acid | C9H8O4 | 179.0347 | −0.3 | 2330 | 135.0457 | [50] |
| 2.54 | 4-O-(4-Coumaroyl)quinic acid | C16H18O8 | 337.0920 | −2.6 | 187 | 191.0561 | [48] |
| 2.86 | Feruloylquinic acid | C17H20O9 | 367.1028 | −1.9 | 5135 | 134.0378, 173.0455, 191.0558, 193.0506 | [48] |
| 3.91 | Dicaffeoylquinic acid isomer 1 | C25H24O12 | 515.1196 | 0.3 | 9330 | 135.0460, 173.0455, 179.0349, 191.0557, 353.0873 | [48] |
| 3.91 | Salvianolic acid derivative isomer 1 | C27H22O12 | 537.1017 | −4.1 | 1294 | 135.0457, 161.0245, 201.0171, 375.0706 | [49] |
| 4.06 | Dicaffeoylquinic acid isomer 2 | C25H24O12 | 515.1196 | 0.2 | 94316 | 135.0457, 173.0456, 179.0348, 191.0560, 353.0878 | [49] |
| 4.06 | Cryptochlorogenic Acid | C16H18O9 | 353.0878 | −0.1 | 11708 | 135.0457, 173.0456, 179.0348, 191.0560 | [49] |
| 4.06 | Salvianolic acid derivative isomer 2 | C27H22O12 | 537.1016 | −4.1 | 19809 | 135.0457, 161.0245, 179.0348, 201.0168, 375.0698 | [49] |
| 4.41 | Dicaffeoylquinic acid isomer 3 | C25H24O12 | 515.1188 | −1.3 | 25919 | 135.0457, 173.0455, 179.0347, 191.0560, 353.0879 | [48] |
| 4.41 | Salvianolic acid derivative isomer 3 | C27H22O12 | 537.1014 | −4.6 | 3150 | 135.0457, 161.0245, 179.0347, 201.0172, 375.0695 | [49] |
| 7.11 | Unknown | C41H56O7 | 659.3939 | −2.1 | 207461 | 599.3732 | - |
| 7.60 | Unknown | C13H25NO3 | 242.1760 | −0.5 | 83368 | 224.1656 | - |
| 8.03 | Pinellic acid | C18H34O5 | 329.2335 | 0.4 | 167137 | 171.1028, 211.1336, 229.1440 | [52] |
| Peak | Rt Value | Area % | Name of Constituent | Formula | Pharmacological Activity |
|---|---|---|---|---|---|
| 1. | 4.58 | 5.37 | Caffeoylquinic acid | C16H18O9 | Activator of NF-κB signaling and macrophage infiltration [55] |
| 2. | 5.271 | 4.47 | Isoxazolidine | C3H7NO | Anti-cancer [56,57], anti-diabetic [58], anti-fungal hypolipidemic activity [59] |
| 3. | 8.921 | 10.87 | ß-Bourbonene | C15H24 | Anti-diabetic and insulinotropic [54] |
| 4. | 8.943 | 3.33 | 3-(4-Methyl piperidin-1-yl)propan-1-amine | C9H20N2 | Anti-Alzheimer’s agent [60] |
| 5. | 10.849 | 13.77 | Feruloylquinic acid | C17H20O9 | Anti-diabetic cardiovascular disease [61] |
| 6. | 10.253 | 4.13 | Carbamic acid | CH3NO2 | Anti-microbial, antifibrinolytics [62] |
| 7. | 11.489 | 0.39 | 4-Amino-1,2,5-oxadiazol-3-ol | C2H3N3O2 | Akt kinase inhibitor [63] |
| 8. | 12.629 | 11.96 | Pent-4-enamide | C5H9NO | Anti-bacterial agent [64] |
| 9. | 13.645 | 1.98 | Morpholinoacetonitrile | C6H10N2O | anti-inflammatory, anti-cancer [65] |
| 10. | 14.378 | 2.52 | 3-Pyrrolidinol | C4H9NO | Antioxidant [66] |
| 11. | 18.833 | 19.91 | 1,4 Dicaffeoyl quinic acid | C25H24O12 | Antioxidant, anti-ulcer [67] |
| 12. | 19.577 | 5.85 | Camphene | C10H16 | Anti-hyperlipidemic and anti-inflammatory [68] |
| 13. | 22.957 | 10.20 | Hexanoic acid | C6H12O2 | Anti-microbial [39] |
| 14. | 23.011 | 13.48 | α-Murrolene | C15H24 | Analgesic, anti-inflammatory, anti-bacterial, and anti-fungal activity [69] |
| 15. | 25.182 | 1.66 | Cyclopentanol | C5H10O | Oxidative stabilizer, anti-oxidants [70] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariadoss, A.V.A.; Park, S.; Saravanakumar, K.; Sathiyaseelan, A.; Wang, M.-H. Ethyl Acetate Fraction of Helianthus tuberosus L. Induces Anti-Diabetic, and Wound-Healing Activities in Insulin-Resistant Human Liver Cancer and Mouse Fibroblast Cells. Antioxidants 2021, 10, 99. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010099
Mariadoss AVA, Park S, Saravanakumar K, Sathiyaseelan A, Wang M-H. Ethyl Acetate Fraction of Helianthus tuberosus L. Induces Anti-Diabetic, and Wound-Healing Activities in Insulin-Resistant Human Liver Cancer and Mouse Fibroblast Cells. Antioxidants. 2021; 10(1):99. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010099
Chicago/Turabian StyleMariadoss, Arokia Vijaya Anand, SeonJu Park, Kandasamy Saravanakumar, Anbazhagan Sathiyaseelan, and Myeong-Hyeon Wang. 2021. "Ethyl Acetate Fraction of Helianthus tuberosus L. Induces Anti-Diabetic, and Wound-Healing Activities in Insulin-Resistant Human Liver Cancer and Mouse Fibroblast Cells" Antioxidants 10, no. 1: 99. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10010099