
Mechanisms Modified by (−)-Epicatechin and Taxifolin Relevant for the Treatment of Hypertension and Viral Infection: Knowledge from Preclinical Studies


Abstract
:1. Introduction
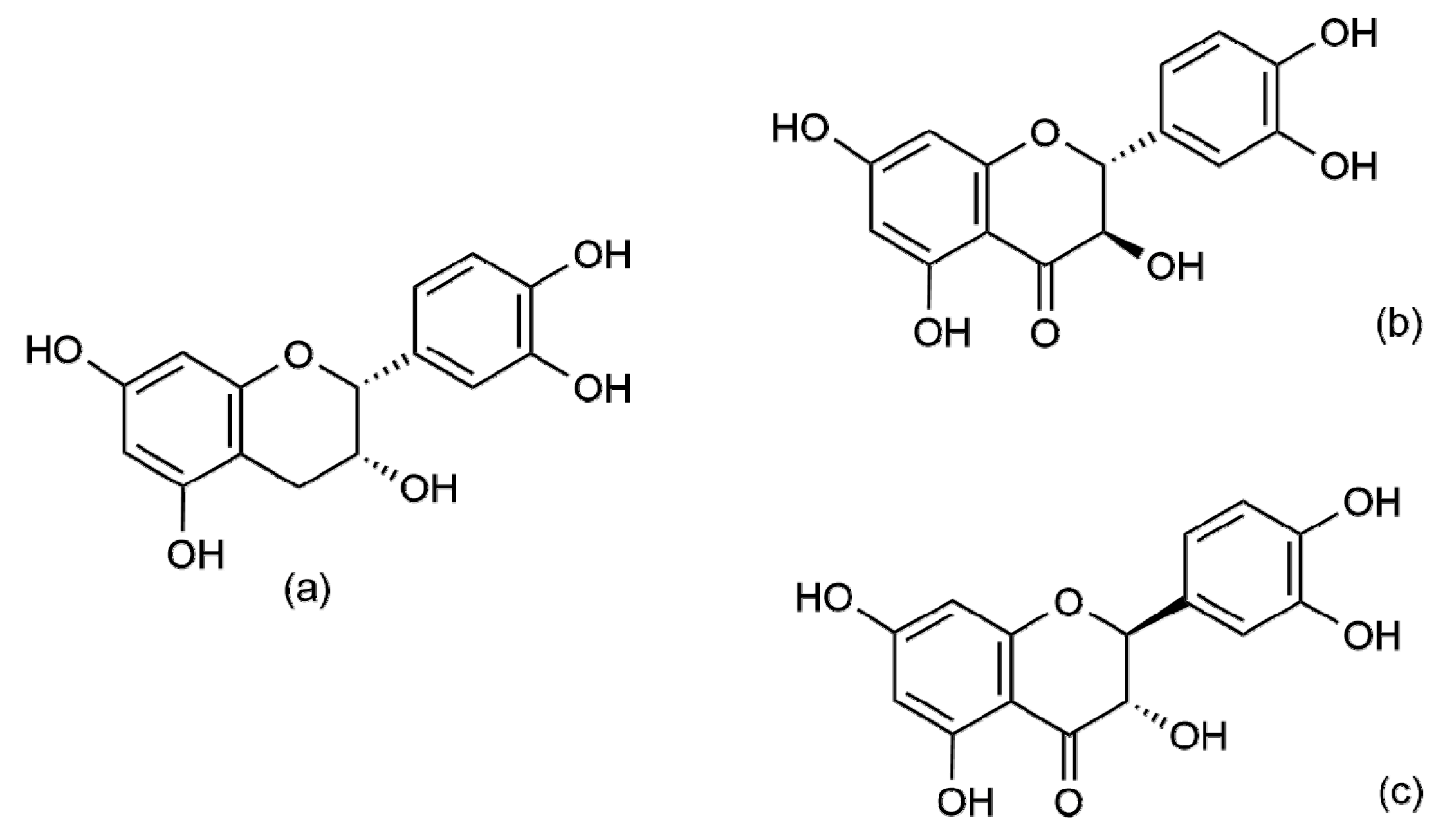
2. Flavonoids EC and TX
3. Absorption and Toxicity of EC and TX
4. Biological Effects of EC and TX
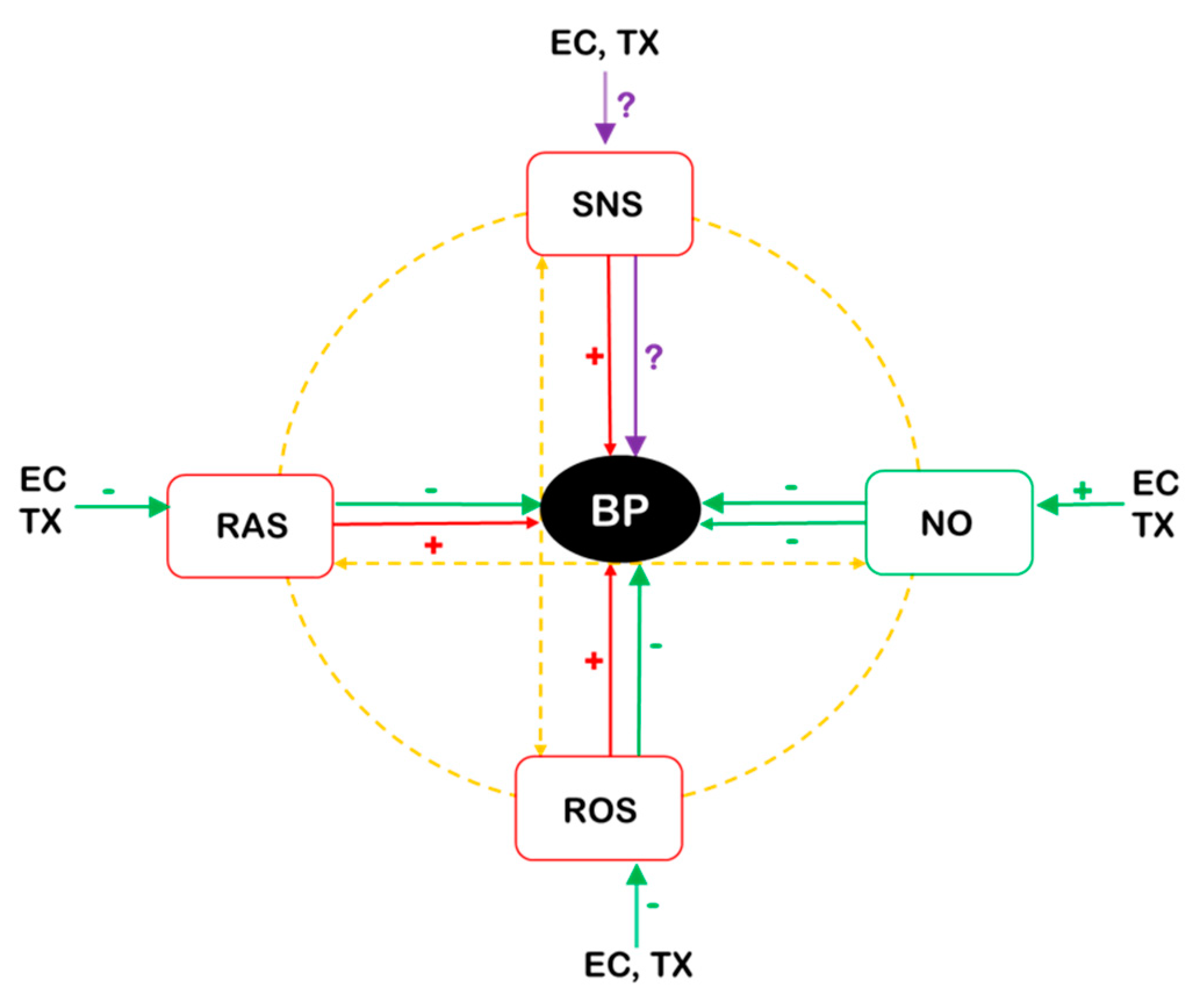
5. Cellular and Molecular Mechanisms of EC and TX Action Relevant to Cardiovascular Function
5.1. EC- and TX-Mediated Defense against Oxidative Stress
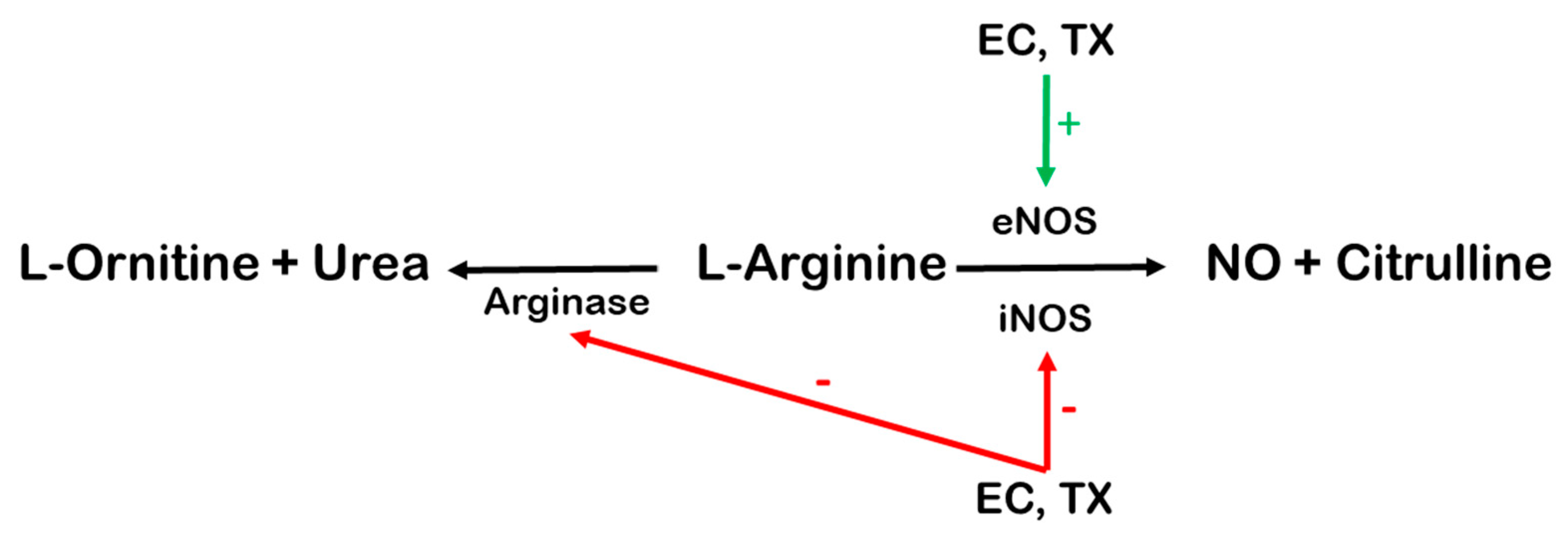
5.2. NO-Mediated Effects of EC and TX
5.3. Anti-Inflammatory Effects of EC and TX
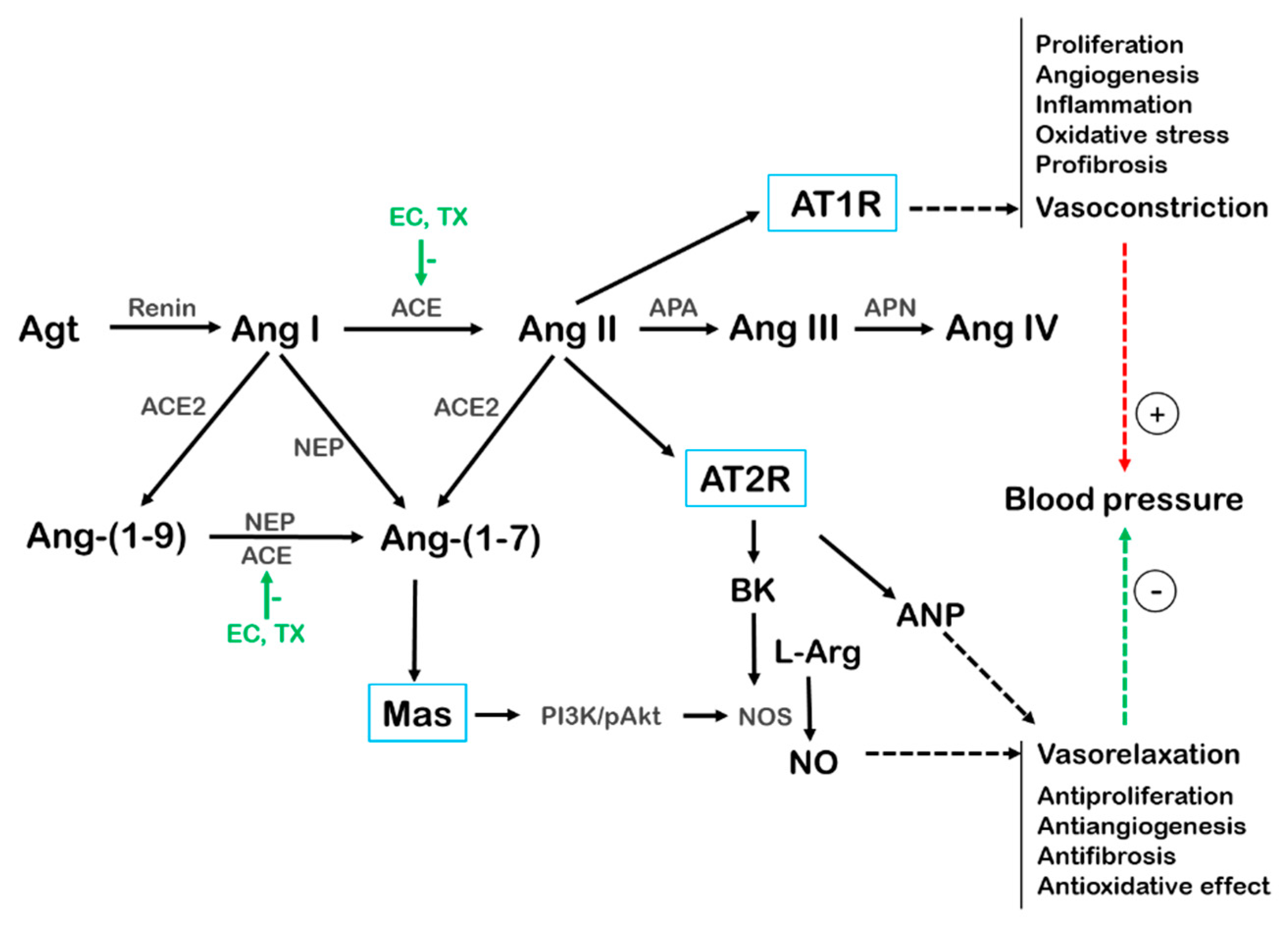
5.4. EC and TX-Mediated Effects on the SNS and the RAS
5.5. Mechanisms Involved in the Vascular Action of EC and TX
5.6. EC and TX Effects on Blood Cells and Rheology
6. Mechanisms of EC and TX Action in Experimental Models of Hypertension
6.1. EC and TX Action in Spontaneously Hypertensive and Borderline Hypertensive Rats
6.2. EC and TX Action in NO-Deficient Hypertension
6.3. Mechanisms of EC and TX Action in Other Models of Hypertension
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACE | angiotensin-converting enzyme |
| Ang II | angiotensin II |
| ARE | antioxidant response element |
| ATR | angiotensin receptor |
| BHRs | borderline hypertensive rats |
| CAT | catalase |
| COVID-19 | coronavirus disease |
| COX | cyclooxygenase |
| CVDs | cardiovascular diseases |
| DOCA | deoxycorticosterone acetate |
| EC | (−)-epicatechin |
| EDCFs | endothelium-derived contracting factors |
| eNOS | endothelial NO synthase activity |
| GPx | glutathione peroxidase |
| HO-1 | hemoxygenase-1 |
| HUVEC | human umbilical veins endothelial cells |
| IL | interleukin |
| IUPAC | International Union of Pure and Applied Chemistry |
| Keap1 | Kelch-like ECH-associating protein 1 |
| L-NAME | NG-nitro-L-arginine methyl ester |
| MAPK | mitogen-activated protein kinase |
| Mpro | main protease |
| NAD(P)H | nicotinamide adenine dinucleotide phosphate |
| NEP | neutral endopeptidase |
| NF-κB | nuclear factor-κB |
| NLRP3 | nucleotide-binding oligomerization domain-, leucine-rich repeat- and pyrin domain-containing protein 3 |
| NLRC4 | neuronal apoptosis inhibitory protein (NAIP)/NLR family caspase activation and recruitment domain–containing protein 4 |
| NO | nitric oxide |
| NODH | NO-deficient hypertension |
| eNOS | endothelial nitric oxide synthase |
| iNOS | inducible nitric oxide synthase |
| NOS | nitric oxide synthase |
| NOX | nicotinamide adenine dinucleotide phosphate oxidase |
| Nrf2 | nuclear factor erythroid-2 related factor 2 |
| PI3K/Akt/PKA | phosphatidylinositol-3-kinase/protein kinase B/protein kinase A |
| RAS | renin-angiotensin system |
| ROS | reactive oxygen species |
| SARS-CoV-2 | severe acute respiratory syndrome corona virus 2 |
| SHRs | spontaneously hypertensive rats |
| SOD | superoxide dismutase |
| SNS | sympathetic nervous system |
| TNFα | tumor necrosis factor α |
| TX | (+)-taxifolin, (−)-taxifolin or their racemic mixture |
| XO | xanthine oxidase |
References
- Forouzanfar, M.H.; Afshin, A.; Alexander, L.T.; Anderson, H.R.; Bhutta, Z.A.; Biryukov, S.; Carrero, J.J. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: A systematic analysis for the global burden of disease study 2015. Lancet 2016, 388, 1659–1724. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Bentham, J.; Di Cesare, M.; Bixby, H.; Danaei, G.; Cowan, M.J.; Cho, B. Worldwide trends in blood pressure from 1975 to 2015: A pooled analysis of 1479 population-based measurement studies with 19·1 million participants. Lancet 2017, 389, 37–55. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.; Qin, M.; Shen, B.; Cai, Y.; Liu, T.; Yang, F.; Gong, W.; Liu, X.; Liang, J.; Zhao, Q.; et al. Association of cardiac injury with mortality in hospitalized patients with COVID-19 in Wuhan, China. JAMA Cardiol. 2020, 5, 802–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Fan, Y.; Chen, M.; Wu, X.; Zhang, L.; He, T.; Wang, H.; Wan, J.; Wang, X.; Lu, Z. Cardiovascular implications of fatal outcomes of patients with coronavirus disease 2019 (COVID-19). JAMA Cardiol. 2020, 5, 811–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Taddei, S.; Bruno, R.M.; Masi, S.; Solini, A. ESC Cardiomed. In Epidemiology and Pathophysiology of Hypertension; Oxford University Press: Oxford, UK, 2020. [Google Scholar] [CrossRef]
- Bernatova, I. Endothelial dysfunction in experimental models of arterial hypertension: Cause or consequence? BioMed Res. Int. 2014, 2014, 598271. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.M.; Wysocki, J.; Batlle, D. Interaction of SARS-CoV-2 and other coronavirus with ACE (angiotensin-converting enzyme)-2 as their main receptor: Therapeutic implications. Hypertension 2020, 76, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef]
- Grassi, D.; Desideri, G.; Croce, G.; Tiberti, S.; Aggio, A.; Ferri, C. Flavonoids, vascular function and cardiovascular protection. Curr. Pharm. Des. 2009, 15, 1072–1084. [Google Scholar] [CrossRef]
- Rees, A.; Dodd, G.F.; Spencer, J.P.E. The effects of flavonoids on cardiovascular health: A review of human intervention trials and implications for cerebrovascular function. Nutrients 2018, 10, 1852. [Google Scholar] [CrossRef] [Green Version]
- Diniz, L.R.L.; Bezerra Filho, C.D.S.M.; Fielding, B.C.; De Sousa, D.P. Natural antioxidants: A review of studies on human and animal coronavirus. Oxid. Med. Cell. Longev. 2020, 2020, 3173281. [Google Scholar] [CrossRef]
- Solnier, J.; Fladerer, J.P. Flavonoids: A complementary approach to conventional therapy of COVID-19? Phytochem. Rev. 2020, 1–23. [Google Scholar] [CrossRef]
- Lin, Y.T.; Wu, Y.H.; Tseng, C.K.; Lin, C.K.; Chen, W.C.; Hsu, Y.C.; Lee, J.C. Green tea phenolic epicatechins inhibit hepatitis C virus replication via cycloxygenase-2 and attenuate virus-induced inflammation. PLoS ONE 2013, 8, e54466. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, P.G.; Ferraz, A.C.; Figueiredo, J.E.; Lima, C.F.; Rodrigues, V.G.; Taranto, A.G.; Ferreira, J.M.S.; Brandão, G.C.; Vieira-Filho, S.A.; Duarte, L.P.; et al. Detection of the antiviral activity of epicatechin isolated from Salacia crassifolia (Celastraceae) against Mayaro virus based on protein C homology modelling and virtual screening. Arch. Virol. 2018, 163, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Evaluation of green tea polyphenols as novel corona virus (SARS CoV-2) main protease (Mpro) inhibitors-an in silico docking and molecular dynamics simulation study. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, N.; Chowdhury, P.; Goswami, A.K.; Das, A.; Chetia, D.; Gogoi, B. Computational guided identification of a citrus flavonoid as potential inhibitor of SARS-CoV-2 main protease. Mol. Divers. 2020, 1–15. [Google Scholar] [CrossRef]
- Fischer, A.; Sellner, M.; Neranjan, S.; Smieško, M.; Lill, M.A. Potential inhibitors for novel coronavirus protease identified by virtual screening of 606 million compounds. Int. J. Mol. Sci. 2020, 21, 3626. [Google Scholar] [CrossRef]
- Epicatechin C15H1406-PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Epicatechin (accessed on 30 January 2021).
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vega-Villa, K.R.; Remsberg, C.M.; Takemoto, J.K.; Ohgami, Y.; Yáñez, J.A.; Andrews, P.K.; Davies, N.M. Stereospecific pharmacokinetics of racemic homoeriodictyol, isosakuranetin, and taxifolin in rats and their disposition in fruit. Chirality 2011, 23, 339–348. [Google Scholar] [CrossRef]
- Miller, K.B.; Hurst, W.J.; Flannigan, N.; Ou, B.; Lee, C.Y.; Smith, N.; Stuart, D.A. Survey of commercially available chocolate- and cocoa-containing products in the United States. 2. Comparison of flavan-3-ol content with nonfat cocoa solids, total polyphenols, and percent cacao. J. Agric. Food Chem. 2009, 57, 9169–9180. [Google Scholar] [CrossRef] [PubMed]
- Alanon, M.E.; Castle, S.M.; Siswanto, P.J.; Cifuentes-Gomez, T.; Spencer, J.P. Assessment of flavanol stereoisomers and caffeine and theobromine content in commercial chocolates. Food Chem. 2016, 208, 177–184. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vågen, I.M. Onions: A source of unique dietary flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef]
- Zhang, W.; Han, F.; He, J.; Duan, C. HPLC-DAD-ESI-MS/MS analysis and antioxidant activities of nonanthocyanin phenolics in mulberry (Morus alba L.). J. Food Sci. 2008, 73, C512–C518. [Google Scholar] [CrossRef]
- Lantto, T.A.; Dorman, H.J.D.; Shikov, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Tikhonov, V.P.; Hiltunen, R.; Raasmaja, A. Chemical composition, antioxidative activity and cell viability effects of a Siberian pine (Pinus sibirica Du Tour) extract. Food Chem. 2009, 112, 936–943. [Google Scholar] [CrossRef]
- Gerhäuser, C. Beer constituents as potential cancer chemopreventive agents. Eur. J. Cancer 2005, 41, 1941–1954. [Google Scholar] [CrossRef] [PubMed]
- Johannot, L.; Somerset, S.M. Age-related variations in flavonoid intake and sources in the Australian population. Public Health Nutr. 2006, 9, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Chen, Y.M.; Huang, J.; Fang, Y.J.; Huang, W.Q.; Yan, B.; Lu, M.S.; Pan, Z.Z.; Zhang, C.X. Flavonoid intake from vegetables and fruits is inversely associated with colorectal cancer risk: A case-control study in China. Br. J. Nutr. 2016, 116, 1275–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, O.K.; Chung, S.J.; Song, W.O. Estimated dietary flavonoid intake and major food sources of U.S. adults. J. Nutr. 2007, 137, 1244–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollenberg, K. Vascular action of cocoa flavanols in humans: The roots of the story. J. Cardiovasc. Pharmacol. 2006, 47, S99–S102. [Google Scholar] [CrossRef]
- Baba, S.; Osakabe, N.; Natsume, M.; Muto, Y.; Takizawa, T.; Terao, J. In vivo comparison of the bioavailability of (+)-catechin, (−)-epicatechin and their mixture in orally administered rats. J. Nutr. 2001, 131, 2885–2891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Z.; Wang, F.; Dai, S.; Lu, J.; Wu, X.; Zhang, J. Profiling and identification of (−)-epicatechin metabolites in rats using ultra-high performance liquid chromatography coupled with linear trap-Orbitrap mass spectrometer. Drug Test. Anal. 2017, 9, 1224–1235. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Borges, G.; Momma, T.Y.; Spencer, J.P.; Keen, C.L.; Crozier, A.; Schroeter, H. The metabolome of [2-(14) C] (−)-epicatechin in humans: Implications for the assessment of efficacy, safety, and mechanisms of action of polyphenolic bioactives. Sci. Rep. 2016, 6, 29034. [Google Scholar] [CrossRef] [Green Version]
- Faria, A.; Pestana, D.; Teixeira, D.; Couraud, P.O.; Romero, I.; Weksler, B.; Mateus, N.; De Freitas, V.; Calhau, C. Insights into the putative catechin and epicatechin transport across blood-brain barrier. Food Funct. 2011, 2, 39–44. [Google Scholar] [CrossRef]
- Faria, A.; Meireles, M.; Fernandes, I.; Santos-Buelga, C.; Gonzalez-Manzano, S.; Duenas, M.; De Freitas, V.; Mateus, N.; Calhau, C. Flavonoid metabolites transport across a human BBB model. Food Chem. 2014, 149, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, Q.L.; Zhang, X.Y.; Lv, C.; Li, J.; Yuan, Y.; Yin, F.X. Pharmacokinetics and blood-brain barrier penetration of (+)-catechin and (−)-epicatechin in rats by microdialysis sampling coupled to high-performance liquid chromatography with chemiluminescence detection. J. Agric. Food Chem. 2012, 60, 9377–9383. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, J.I.; Balz, M.; Kimball, J.; Ensunsa, J.L.; Fong, R.; Momma, T.Y.; Kwik-Uribe, C.; Schroeter, H.; Keen, C.L. Safety and efficacy of cocoa flavanol intake in healthy adults: A randomized, controlled, double-masked trial. Am. J. Clin. Nutr. 2015, 102, 1425–1435. [Google Scholar] [CrossRef] [Green Version]
- Kluknavsky, M.; Balis, P.; Puzserova, A.; Radosinska, J.; Berenyiova, A.; Drobna, M.; Lukac, S.; Muchova, J.; Bernatova, I. (−)-Epicatechin prevents blood pressure increase and reduces locomotor hyperactivity in young spontaneously hypertensive rats. Oxid. Med. Cell. Longev. 2016, 2016, 6949020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.Q.; Wang, Y.J.; Zhou, X.F. Effects of (−) Epicatechin on the pathology of APP/PS1 transgenic mice. Front. Neurol. 2014, 5, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuang, H.; Tang, Z.; Zhang, C.; Wang, Z.; Li, W.; Yang, C.; Wang, Q.; Yang, B.; Kong, A.N. Taxifolin Activates the Nrf2 anti-oxidative stress pathway in mouse skin epidermal JB6 P+ cells through epigenetic modifications. Int. J. Mol. Sci. 2017, 18, 1546. [Google Scholar] [CrossRef] [Green Version]
- Voskoboinikova, I.V.; Tjukavkina, N.A.; Geodakyan, S.V.; Kolesnik, Y.A.; Kolhir, V.K.; Zjuzin, V.A.; Sokolov, S.J. Experimental pharmacokinetics of biologically active plant phenolic compounds III. Pharmacokinetics of dihydroquercetin. Phytother. Res. 1993, 7, 208–210. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products; Nutrition and Allergies (NDA); Turck, D.; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; et al. Scientific Opinion on taxifolin-rich extract from Dahurian Larch (Larix gmelinii). EFSA J. 2017, 15. [CrossRef]
- Pozharitskaya, O.N.; Karlina, M.V.; Shikov, A.N.; Kosman, V.M.; Makarova, M.N.; Makarov, V.G. Determination and pharmacokinetic study of taxifolin in rabbit plasma by high-performance liquid chromatography. Phytomedicine 2009, 16, 244–251. [Google Scholar] [CrossRef]
- Booth, A.N.; Deeds, F. The toxicity and metabolism of dihydroquercetin. J. Am. Pharm. Assoc. 1958, 47, 183–184. [Google Scholar] [CrossRef]
- Yang, P.; Xu, F.; Li, H.F.; Wang, Y.; Li, F.C.; Shang, M.Y.; Liu, G.X.; Wang, X.; Cai, S.Q. Detection of 191 taxifolin metabolites and their distribution in rats using HPLC-ESI-IT-TOF-MS(n). Molecules 2016, 21, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhanataev, A.K.; Kulakova, A.V.; Nasonova, V.V.; Durnev, A.D. In vivo study of dihydroquercetin genotoxicity. Bull. Exp. Biol. Med. 2008, 145, 338–340. [Google Scholar] [CrossRef]
- Celyico, S.S. Sub-Acute Toxicity Study of Lavitol (Dyhydroquercetin) in the Wistar Rat; Amur State Medical Academy: Blagoveshchensk, Russia, 2008. [Google Scholar]
- Schauss, A.G.; Tselyico, S.S.; Kuznetsova, V.A.; Yegorova, I. Toxicological and genotoxicity assessment of a dihydroquercetin-rich dahurian larch tree (Larix gmelinii Rupr) extract (Lavitol). Int. J. Toxicol. 2015, 34, 162–181. [Google Scholar] [CrossRef]
- Zu, Y.; Wu, W.; Zhao, X.; Li, Y.; Wang, W.; Zhong, C.; Zhang, Y.; Zhao, X. Enhancement of solubility, antioxidant ability and bioavailability of taxifolin nanoparticles by liquid antisolvent precipitation technique. Int. J. Pharm. 2014, 471, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Grimm, T.; Skrabala, R.; Chovanová, Z.; Muchová, J.; Sumegová, K.; Liptáková, A.; Duracková, Z.; Högger, P. Single and multiple dose pharmacokinetics of maritime pine bark extract (pycnogenol) after oral administration to healthy volunteers. BMC Clin. Pharmacol. 2006, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Zinchenko, V.P.; Kim, Y.A.; Tarakhovskii, Y.S.; Bronnikov, G.E. Biological activity of water-soluble nanostructures of dihydroquercetin with cyclodextrins. Biophysics 2011, 56, 418. [Google Scholar] [CrossRef]
- Shikov, A.N.; Pozharitskaya, O.N.; Miroshnyk, I.; Mirza, S.; Urakova, I.N.; Hirsjärvi, S.; Makarov, V.G.; Heinämäki, J.; Yliruusi, J.; Hiltunen, R. Nanodispersions of taxifolin: Impact of solid-state properties on dissolution behavior. Int. J. Pharm. 2009, 377, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ruiz, A.G.; Ganem, A.; Olivares-Corichi, I.M.; García-Sánchez, J.R. Lecithin-chitosan—TPGS nanoparticles as nanocarriers of (−)-epicatechin enhanced its anticancer activity in breast cancer cells. RSC Adv. 2018, 8, 34773–34782. [Google Scholar] [CrossRef] [Green Version]
- Rusznyak, S.T.; Szent-Gyorgyi, A. Vitamin P: Flavonols as Vitamins. Nature 1936, 138, 27. [Google Scholar] [CrossRef]
- Bernatova, I. Biological activities of (−)-epicatechin and (−)-epicatechin-containing foods: Focus on cardiovascular and neuropsychological health. Biotechnol. Adv. 2018, 36, 666–681. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [Green Version]
- Maher, P. The potential of flavonoids for the treatment of neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 3056. [Google Scholar] [CrossRef] [Green Version]
- Garate-Carrillo, A.; Navarrete-Yañez, V.; Ortiz-Vilchis, P.; Guevara, G.; Castillo, C.; Mendoza-Lorenzo, P.; Ceballos, G.; Ortiz-Flores, M.; Najera, N.; Bustamante-Pozo, M.M.; et al. Arginase inhibition by (−)-epicatechin reverses endothelial cell aging. Eur. J. Pharmacol. 2020, 885, 173442. [Google Scholar] [CrossRef] [PubMed]
- Galleano, M.; Bernatova, I.; Puzserova, A.; Balis, P.; Sestakova, N.; Pechanova, O.; Fraga, C.G. (−)-Epicatechin reduces blood pressure and improves vasorelaxation in spontaneously hypertensive rats by NO-mediated mechanism. IUBMB Life 2013, 65, 710–715. [Google Scholar] [CrossRef]
- Kluknavsky, M.; Balis, P.; Skratek, M.; Manka, J.; Bernatova, I. (−)-Epicatechin reduces the blood pressure of young borderline hypertensive rats during the post-treatment period. Antioxidants 2020, 9, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Guzmán, M.; Jiménez, R.; Sánchez, M.; Zarzuelo, M.J.; Galindo, P.; Quintela, A.M.; López-Sepulveda, R.; Romero, M.; Tamargo, J.; Vargas, F.; et al. Epicatechin lowers blood pressure, restores endothelial function, and decreases oxidative stress and endothelin-1 and NADPH oxidase activity in DOCA-salt hypertension. Free Radic. Biol. Med. 2012, 52, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Prince, P.D.; Lanzi, C.R.; Toblli, J.E.; Elesgaray, R.; Oteiza, P.I.; Fraga, C.G.; Galleano, M. Dietary (−)-epicatechin mitigates oxidative stress, NO metabolism alterations, and inflammation in renal cortex from fructose-fed rats. Free Radic. Biol. Med. 2015, 90, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Piotrkowski, B.; Calabro, V.; Galleano, M.; Fraga, C.G. (−)-Epicatechin prevents alterations in the metabolism of superoxide anion and nitric oxide in the hearts of L-NAME-treated rats. Food Funct. 2015, 6, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, M.B.; Aliev, O.I.; Sidekhmenova, A.V.; Shamanaev, A.Y.; Anishchenko, A.M.; Nosarev, A.V.; Pushkina, E.A. Modes of hypotensive action of dihydroquercetin in arterial hypertension. Bull. Exp. Biol. Med. 2017, 162, 353–356. [Google Scholar] [CrossRef]
- Kim, K.J.; Hwang, E.S.; Kim, M.J.; Park, J.H.; Kim, D.O. Antihypertensive effects of polyphenolic extract from Korean Red Pine (Pinus densiflora Sieb. et Zucc.) bark in spontaneously hypertensive rats. Antioxidants 2020, 9, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, C.J.; Kubo, E.; Fujii, K.; Nishimura, Y.; Kobuchi, S.; Ohkita, M.; Yoshimura, M.; Kiso, Y.; Matsumura, Y. Antihypertensive effect of French maritime pine bark extract (Flavangenol): Possible involvement of endothelial nitric oxide-dependent vasorelaxation. J. Hypertens. 2009, 27, 92–101. [Google Scholar] [CrossRef]
- Aggio, A.; Grassi, D.; Onori, E.; D’Alessandro, A.; Masedu, F.; Valenti, M.; Ferri, C. Endothelium/nitric oxide mechanism mediates vasorelaxation and counteracts vasoconstriction induced by low concentration of flavanols. Eur. J. Nutr. 2013, 52, 263–272. [Google Scholar] [CrossRef]
- MacRae, K.; Connolly, K.; Vella, R.; Fenning, A. Epicatechin’s cardiovascular protective effects are mediated via opioid receptors and nitric oxide. Eur. J. Nutr. 2019, 58, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Novakovic, A.; Marinko, M.; Vranic, A.; Jankovic, G.; Milojevic, P.; Stojanovic, I.; Nenezic, D.; Ugresic, N.; Kanjuh, V.; Yang, Q.; et al. Mechanisms underlying the vasorelaxation of human internal mammary artery induced by (−)-epicatechin. Eur. J. Pharmacol. 2015, 762, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Marinko, M.; Jankovic, G.; Nenezic, D.; Milojevic, P.; Stojanovic, I.; Kanjuh, V.; Novakovic, A. (−)-Epicatechin-induced relaxation of isolated human saphenous vein: Roles of K+ and Ca2+ channels. Phytother. Res. 2018, 32, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Plotnikov, M.B.; Aliev, O.I.; Sidekhmenova, A.V.; Shamanaev, A.Y.; Anishchenko, A.M.; Fomina, T.I.; Chernysheva, G.A.; Smol’yakova, V.I.; Arkhipov, A.M. Dihydroquercetin improves microvascularization and microcirculation in the brain cortex of SHR rats during the development of arterial hypertension. Bull. Exp. Biol. Med. 2017, 163, 57–60. [Google Scholar] [CrossRef]
- Shah, Z.A.; Li, R.C.; Ahmad, A.S.; Kensler, T.W.; Yamamoto, M.; Biswal, S.; Doré, S. The flavanol (−)-epicatechin prevents stroke damage through the Nrf2/HO1 pathway. J. Cereb. Blood Flow Metab. 2010, 30, 1951–1961. [Google Scholar] [CrossRef] [Green Version]
- Calabró, V.; Piotrkowski, B.; Fischerman, L.; Vazquez Prieto, M.A.; Galleano, M.; Fraga, C.G. Modifications in nitric oxide and superoxide anion metabolism induced by fructose overload in rat heart are prevented by (−)-epicatechin. Food Funct. 2016, 7, 1876–1883. [Google Scholar] [CrossRef]
- Yamazaki, K.G.; Taub, P.R.; Barraza-Hidalgo, M.; Rivas, M.M.; Zambon, A.C.; Ceballos, G.; Villarreal, F.J. Effects of (−)-epicatechin on myocardial infarct size and left ventricular remodeling after permanent coronary occlusion. J. Am. Coll. Cardiol. 2010, 55, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wang, T.; Fu, F.F.; El-Kassaby, Y.A.; Wang, G. Temporospatial flavonoids metabolism variation in Ginkgo biloba leaves. Front. Genet. 2020, 11, 1503. [Google Scholar] [CrossRef]
- Shu, Z.; Yang, Y.; Yang, L.; Jiang, H.; Yu, X.; Wang, Y. Cardioprotective effects of dihydroquercetin against ischemia reperfusion injury by inhibiting oxidative stress and endoplasmic reticulum stress-induced apoptosis via the PI3K/Akt pathway. Food Funct. 2019, 10, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Cremonini, E.; Bettaieb, A.; Haj, F.G.; Fraga, C.G.; Oteiza, P.I. (−)-Epicatechin improves insulin sensitivity in high fat diet-fed mice. Arch. Biochem. Biophys. 2016, 599, 13–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Salmeán, G.; Ortiz-Vilchis, P.; Vacaseydel, C.M.; Rubio-Gayosso, I.; Meaney, E.; Villarreal, F.; Ramírez-Sánchez, I.; Ceballos, G. Acute effects of an oral supplement of (−)-epicatechin on postprandial fat and carbohydrate metabolism in normal and overweight subjects. Food Funct. 2014, 5, 521–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Xu, N.; Zhao, W.; Su, J.; Liang, M.; Xie, Z.; Wu, X.; Li, Q. (−)-Epicatechin regulates blood lipids and attenuates hepatic steatosis in rats fed high-fat dietdagger. Mol. Nutr. Food Res. 2017, 61, 1700303. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Salmeán, G.; Ortiz-Vilchis, P.; Vacaseydel, C.M.; Garduño-Siciliano, L.; Chamorro-Cevallos, G.; Meaney, E.; Villafaña, S.; Villarreal, F.; Ceballos, G.; Ramírez-Sánchez, I. Effects of (−)-epicatechin on a diet-induced rat model of cardiometabolic risk factors. Eur. J. Pharmacol. 2014, 728, 24–30. [Google Scholar] [CrossRef]
- Varela, C.E.; Rodriguez, A.; Romero-Valdovinos, M.; Mendoza-Lorenzo, P.; Mansour, C.; Ceballos, G.; Villarreal, F.; Ramirez-Sanchez, I. Browning effects of (−)-epicatechin on adipocytes and white adipose tissue. Eur. J. Pharmacol. 2017, 811, 48–59. [Google Scholar] [CrossRef]
- Yoon, K.D.; Lee, J.Y.; Kim, T.Y.; Kang, H.; Ha, K.S.; Ham, T.H.; Ryu, S.N.; Kang, M.Y.; Kim, Y.H.; Kwon, Y.I. In vitro and in vivo anti-hyperglycemic activities of taxifolin and its derivatives isolated from Pigmented Rice (Oryzae sativa L. cv. Superhongmi). J. Agric. Food Chem. 2020, 68, 742–750. [Google Scholar] [CrossRef]
- Salama, S.A.; Kabel, A.M. Taxifolin ameliorates iron overload-induced hepatocellular injury: Modulating PI3K/AKT and p38 MAPK signaling, inflammatory response, and hepatocellular regeneration. Chem. Biol. Interact. 2020, 330, 109230. [Google Scholar] [CrossRef] [PubMed]
- Shariati, S.; Kalantar, H.; Pashmforoosh, M.; Mansouri, E.; Khodayar, M.J. Epicatechin protective effects on bleomycin-induced pulmonary oxidative stress and fibrosis in mice. Biomed. Pharmacother. 2019, 114, 108776. [Google Scholar] [CrossRef]
- Unver, E.; Tosun, M.; Olmez, H.; Kuzucu, M.; Cimen, F.K.; Suleyman, Z. The effect of taxifolin on cisplatin-induced pulmonary damage in rats: A biochemical and histopathological evaluation. Mediators Inflamm. 2019, 2019, 3740867. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.C.D.P.; Seito, L.N.; Di Stasi, L.C.; Akiko Hiruma-Lima, C.; Pellizzon, C.H. Epicatechin used in the treatment of intestinal inflammatory disease: An analysis by experimental models. Evid. Based Complement. Alternat. Med. 2012, 2012, 508902. [Google Scholar] [CrossRef] [Green Version]
- Prince, P.D.; Fischerman, L.; Toblli, J.E.; Fraga, C.G.; Galleano, M. LPS-induced renal inflammation is prevented by (−)-epicatechin in rats. Redox Biol. 2017, 11, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Wang, Z.; Oteiza, P.I. (−)-Epicatechin mitigates high fat diet-induced neuroinflammation and altered behavior in mice. Food Funct. 2020, 11, 5065–5076. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Saito, S.; Tanaka, M.; Yamakage, H.; Kusakabe, T.; Shimatsu, A.; Ihara, M.; Satoh-Asahara, N. Pleiotropic neuroprotective effects of taxifolin in cerebral amyloid angiopathy. Proc. Natl. Acad. Sci. USA 2019, 116, 10031–10038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Song, J.; Zhang, M.; Wang, H.; Zhang, Y.; Suo, H. Comparison of in vitro and in vivo antioxidant activities of six flavonoids with similar structures. Antioxidants 2020, 9, 732. [Google Scholar] [CrossRef]
- Plotnikov, M.B.; Plotnikov, D.M.; Alifirova, V.M.; Aliev, O.I.; Maslov, M.; Vasil’ev, A.S.; Tiukavkina, N.A. Clinical efficacy of a novel hemorheological drug ascovertin in patients with vascular encephalopathy. Zh. Nevrol. Psikhiatr. Im. S S Korsakova 2004, 104, 33–37. [Google Scholar]
- Sinegre, T.; Teissandier, D.; Milenkovic, D.; Morand, C.; Lebreton, A. Epicatechin influences primary hemostasis, coagulation and fibrinolysis. Food Funct. 2019, 10, 7291–7298. [Google Scholar] [CrossRef]
- Sinegre, T.; Milenkovic, D.; Bourgne, C.; Teissandier, D.; Nasri, Y.; Dannus, L.T.; Morand, C.; Lebreton, A. Impact of epicatechin on the procoagulant activities of microparticles. Nutrients 2020, 12, 2935. [Google Scholar] [CrossRef]
- Kubatiev, A.A.; Yadigarova, Z.T.; Rud’ko, I.A.; Tyukavkina, N.A.; Bykov, V.A. Diquertin suppresses ADP- and thrombin-induced accumulation of cytoplasmic calcium in human thrombocytes. Pharm. Chem. J. 1999, 33, 629–630. [Google Scholar] [CrossRef]
- Ivanov, I.S.; Sidehmenova, A.V.; Smol’yakova, V.I.; Chernysheva, G.A.; Plotnikov, M.B. Inhibition of adenosine diphosphate-induced platelet aggregation by alpha-lipoic acid and dihydroquercetin in vitro. Indian J. Pharmacol. 2014, 46, 430–432. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Deuster, P. Comparison of quercetin and dihydroquercetin: Antioxidant-independent actions on erythrocyte and platelet membrane. Chem. Biol. Interact. 2009, 182, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, B.L.; Xu, C.G.; Zhou, X.J. Dihydroquercetin protects against renal fibrosis by activating the Nrf2 pathway. Phytomedicine 2020, 69, 153185. [Google Scholar] [CrossRef]
- Majzunova, M.; Dovinova, I.; Barancik, M.; Chan, J.Y. Redox signaling in pathophysiology of hypertension. J. Biomed. Sci. 2013, 20, 69. [Google Scholar] [CrossRef] [Green Version]
- Peterhans, E. Reactive oxygen species and nitric oxide in viral diseases. Biol. Trace Elem. Res. 1997, 56, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I.; et al. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Lingappan, K. NF-κB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, M.A.; Hayes, J.D. The Keap1/Nrf2 pathway in health and disease: From the bench to the clinic. Biochem. Soc. Trans. 2015, 43, 687–689. [Google Scholar] [CrossRef] [Green Version]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Ruijters, E.J.; Weseler, A.R.; Kicken, C.; Haenen, G.R.; Bast, A. The flavanol (−)-epicatechin and its metabolites protect against oxidative stress in primary endothelial cells via a direct antioxidant effect. Eur. J. Pharmacol. 2013, 715, 147–153. [Google Scholar] [CrossRef]
- Kostyuk, V.A.; Potapovich, A.I.; Strigunova, E.N.; Kostyuk, T.V.; Afanas’ev, I.B. Experimental evidence that flavonoid metal complexes may act as mimics of superoxide dismutase. Arch. Biochem. Biophys. 2004, 428, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Říha, M.; Karlíčková, J.; Filipský, T.; Macáková, K.; Rocha, L.; Bovicelli, P.; Silvestri, I.P.; Saso, L.; Jahodář, L.; Hrdina, R.; et al. In vitro evaluation of copper-chelating properties of flavonoids. RSC Adv. 2014, 4, 32628–32638. [Google Scholar] [CrossRef] [Green Version]
- Shubina, V.S.; Shatalin, Y.V. Antioxidant and iron-chelating properties of taxifolin and its condensation product with glyoxylic acid. J. Food Sci. Technol. 2017, 54, 1467–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steffen, Y.; Gruber, C.; Schewe, T.; Sies, H. Mono-O-methylated flavanols and other flavonoids as inhibitors of endothelial NADPH oxidase. Arch. Biochem. Biophys. 2008, 469, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.-I.; Nihei, K.-I.; Ishihara, Y.; Yoshizawa, F.; Yagasaki, K. Anti-hyperuricemic effect of taxifolin in cultured hepatocytes and model mice. Cytotechnology 2017, 69, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Kuwabara, M.; Hisatome, I.; Niwa, K.; Hara, S.; Roncal-Jimenez, C.A.; Bjornstad, P.; Nakagawa, T.; Andres-Hernando, A.; Sato, Y.; Jensen, T.; et al. Uric acid is a strong risk marker for developing hypertension from prehypertension: A 5-year Japanese cohort study. Hypertension 2018, 71, 78–86. [Google Scholar] [CrossRef]
- Balis, P.; Berenyiova, A.; Radosinska, J.; Kvandova, M.; Bernatova, I.; Puzserova, A. High concentration of uric acid failed to affect endothelial function of small mesenteric arteries, femoral arteries and aortas from aged Wistar-Kyoto rats. J. Physiol. Pharmacol. 2020, 71, 399–408. [Google Scholar] [CrossRef]
- Okushio, K.; Suzuki, M.; Matsumoto, N.; Nanjo, F.; Hara, Y. Identification of (−)-epicatechin metabolites and their metabolic fate in the rat. Drug Metab. Dispos. 1999, 27, 309–316. [Google Scholar]
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In Vitro Antioxidant versus Metal Ion Chelating Properties of Flavonoids: A Structure-Activity Investigation. PLoS ONE 2016, 11, e0165575. [Google Scholar] [CrossRef]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; Leon, R.; Lopez, M.G.; Oliva, B.; et al. Transcription factor NRF2 as a therapeutic target for chronic diseases: A systems medicine approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.; Sun, W.; Zhang, Z.; Zheng, Y. The role of Nrf2-mediated pathway in cardiac remodeling and heart failure. Oxid. Med. Cell. Longev. 2014, 2014, 260429. [Google Scholar] [CrossRef] [Green Version]
- Kerins, M.J.; Ooi, A. The roles of NRF2 in modulating cellular iron homeostasis. Antioxid. Redox Signal. 2018, 29, 1756–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravingerová, T.; Kindernay, L.; Barteková, M.; Ferko, M.; Adameová, A.; Zohdi, V.; Bernátová, I.; Ferenczyová, K.; Lazou, A. The molecular mechanisms of iron metabolism and its role in cardiac dysfunction and cardioprotection. Int. J. Mol. Sci. 2020, 21, 7889. [Google Scholar] [CrossRef]
- Barančík, M.; Grešová, L.; Barteková, M.; Dovinová, I. Nrf2 as a key player of redox regulation in cardiovascular diseases. Physiol. Res. 2016, 65, S1–S10. [Google Scholar] [CrossRef] [PubMed]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef] [Green Version]
- Leonardo, C.C.; Doré, S. Dietary flavonoids are neuroprotective through Nrf2-coordinated induction of endogenous cytoprotective proteins. Nutr. Neurosci. 2011, 14, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhan, J.; Zhao, M.; Dai, H.; Deng, Y.; Zhou, W.; Zhao, L. Protective mechanism of taxifolin for chlorpyrifos neurotoxicity in BV2 cells. Neurotoxicology 2019, 74, 74–80. [Google Scholar] [CrossRef]
- Lee, S.B.; Cha, K.H.; Selenge, D.; Solongo, A.; Nho, C.W. The Chemopreventive effect of taxifolin is exerted through ARE-dependent gene regulation. Biol. Pharm. Bull. 2007, 30, 1074–1079. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Sanchez, I.; Maya, L.; Ceballos, G.; Villarreal, F. (−)-epicatechin activation of endothelial cell endothelial nitric oxide synthase, nitric oxide, and related signaling pathways. Hypertension 2010, 55, 1398–1405. [Google Scholar] [CrossRef]
- Moreno-Ulloa, A.; Romero-Perez, D.; Villarreal, F.; Ceballos, G.; Ramirez-Sanchez, I. Cell membrane mediated (−)-epicatechin effects on upstream endothelial cell signaling: Evidence for a surface receptor. Bioorg. Med. Chem. Lett 2014, 24, 2749–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Sanchez, I.; Aguilar, H.; Ceballos, G.; Villarreal, F. (−)-Epicatechin-induced calcium independent eNOS activation: Roles of HSP90 and AKT. Mol. Cell. Biochem. 2012, 370, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Sánchez, I.; Rodríguez, A.; Moreno-Ulloa, A.; Ceballos, G.; Villarreal, F. (−)-Epicatechin-induced recovery of mitochondria from simulated diabetes: Potential role of endothelial nitric oxide synthase. Diab. Vasc. Dis. Res. 2016, 13, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Romanska, H.M.; Polak, J.M.; Coleman, R.A.; James, R.S.; Harmer, D.W.; Allen, J.C.; Bishop, A.E. iNOS gene upregulation is associated with the early proliferative response of human lung fibroblasts to cytokine stimulation. J. Pathol. 2002, 197, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, D.; Talero, E.; Siracusa, R.; Alcaide, A.; Cordaro, M.; Maria Zubelia, J.; Bruschetta, G.; Crupi, R.; Esposito, E.; Cuzzocrea, S.; et al. Protective effect of polyphenols in an inflammatory process associated with experimental pulmonary fibrosis in mice. Br. J. Nutr. 2015, 114, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Sun, X.; Xia, T.; Mao, Q.; Zhong, L. Pretreatment with dihydroquercetin, a dietary flavonoid, protected against concanavalin A-induced immunological hepatic injury in mice and TNF-α/ActD-induced apoptosis in HepG2 cells. Food Funct. 2018, 9, 2341–2352. [Google Scholar] [CrossRef]
- Choi, S.E.; Park, K.H.; Han, B.H.; Jeong, M.S.; Seo, S.J.; Lee, D.I.; Joo, S.S.; Lee, M.W. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by phenolic compounds from roots of Rhododendron mucronulatum. Phytother. Res. 2011, 25, 1301–1305. [Google Scholar] [CrossRef]
- Ortiz-Vilchis, P.; Ortiz-Flores, M.; Pacheco, M.; Ramirez-Sanchez, I.; Moreno-Ulloa, A.; Vega, L.; Ortiz, A.; Villarreal, F.; Rubio-Gayosso, I.; Najera, N.; et al. The cardioprotective effects of (−)-Epicatechin are mediated through arginase activity inhibition in a murine model of ischemia/reperfusion. Eur. J. Pharmacol. 2018, 818, 335–342. [Google Scholar] [CrossRef]
- Bordage, S.; Pham, T.N.; Zedet, A.; Gugglielmetti, A.S.; Nappey, M.; Demougeot, C.; Girard-Thernier, C. Investigation of mammal arginase inhibitory properties of natural ubiquitous polyphenols by using an optimized colorimetric microplate assay. Planta Med. 2017, 83, 647–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Wang, H.; Wang, X.; Zhu, L.; Mao, L. The absence of Nrf2 enhances NF-κB-dependent inflammation following scratch injury in mouse primary cultured astrocytes. Mediat. Inflamm. 2012, 2012, 217580. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef]
- Solak, Y.; Afsar, B.; Vaziri, N.D.; Aslan, G.; Yalcin, C.E.; Covic, A.; Kanbay, M. Hypertension as an autoimmune and inflammatory disease. Hypertens. Res. 2016, 39, 567–573. [Google Scholar] [CrossRef]
- Morrison, M.; Van der Heijden, R.; Heeringa, P.; Kaijzel, E.; Verschuren, L.; Blomhoff, R.; Kooistra, T.; Kleemann, R. Epicatechin attenuates atherosclerosis and exerts anti-inflammatory effects on diet-induced human-CRP and NFkappaB in vivo. Atherosclerosis 2014, 233, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Cremonini, E.; Kang, H.; Kang, J.; Haj, F.G.; Oteiza, P.I. Anti-inflammatory actions of (−)-epicatechin in the adipose tissue of obese mice. Int. J. Biochem. Cell Biol. 2016, 81, 383–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, M.B.; Bhalla, T.N.; Gupta, G.P.; Mitra, C.R.; Bhargava, K.P. Anti-inflammatory activity of taxifolin. Jpn. J. Pharmacol. 1971, 21, 377–382. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Wang, W.-Y.; Chang, C.-C.; Liou, K.-T.; Sung, Y.-J.; Liao, J.-F.; Chen, C.-F.; Chang, S.; Hou, Y.-C.; Chou, Y.-C.; et al. Taxifolin ameliorates cerebral ischemia-reperfusion injury in rats throughits anti-oxidative effect and modulation of NF-kappa B activation. J. Biomed. Sci. 2006, 13, 127–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, T.; Wang, S.; Zhang, X.; Zai, W.; Fan, J.; Chen, W.; Bian, Q.; Luan, J.; Shen, Y.; Zhang, Y.; et al. Kidney protection effects of dihydroquercetin on diabetic nephropathy through suppressing ROS and NLRP3 inflammasome. Phytomedicine 2018, 41, 45–53. [Google Scholar] [CrossRef]
- Ye, Y.; Wang, X.; Cai, Q.; Zhuang, J.; Tan, X.; He, W.; Zhao, M. Protective effect of taxifolin on H2O2-induced H9C2 cell pyroptosis. Zhong Nan Da Xue Xue Bao. Yi Xue Ban J. Cent. South Univ. Med. Sci. 2017, 42, 1367–1374. [Google Scholar] [CrossRef]
- Rodrigues, T.S.; De Sá, K.S.G.; Ishimoto, A.Y.; Becerra, A.; Oliveira, S.; Almeida, L.; Gonçalves, A.V.; Perucello, D.B.; Andrade, W.A.; Castro, R.; et al. Inflammasomes are activated in response to SARS-CoV-2 infection and are associated with COVID-19 severity in patients. J. Exp. Med. 2021, 218. [Google Scholar] [CrossRef]
- Nakagawa, P.; Gomez, J.; Grobe, J.L.; Sigmund, C.D. The renin-angiotensin system in the central nervous system and its role in blood pressure regulation. Curr. Hypertens. Rep. 2020, 22, 7. [Google Scholar] [CrossRef]
- Zicha, J.; Hojná, S.; Kopkan, L.; Červenka, L.; Vaněčková, I. The absence of sympathoexcitation during the development of hypertension in Cyp1a1 Ren-2 transgenic rats. Physiol. Res. 2019, 68, 329–334. [Google Scholar] [CrossRef]
- Drábková, N.; Hojná, S.; Zicha, J.; Vaněčková, I. Contribution of selected vasoactive systems to blood pressure regulation in two models of chronic kidney disease. Physiol. Res. 2020, 69, 405–414. [Google Scholar] [CrossRef]
- Dillenburg, D.R.; Mostarda, C.; Moraes-Silva, I.C.; Ferreira, D.; Bos, D.D.; Duarte, A.A.; Irigoyen, M.C.; Rigatto, K. Resveratrol and grape juice differentially ameliorate cardiovascular autonomic modulation in L-NAME-treated rats. Auton. Neurosci. 2013, 179, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Nakagawa, Y.; Mikome, K.; Yamamoto, H.; Osakabe, N. Enhancement of energy expenditure following a single oral dose of flavan-3-ols associated with an increase in catecholamine secretion. PLoS ONE 2014, 9, e0112180. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Ishimura, K.; Oya, S.; Kamino, M.; Fujii, Y.; Nanba, F.; Toda, T.; Ishii, T.; Adachi, T.; Suhara, Y.; et al. Comparison of the sympathetic stimulatory abilities of B-type procyanidins based on induction of uncoupling protein-1 in brown adipose tissue (BAT) and increased plasma catecholamine (CA) in mice. PLoS ONE 2018, 13, e0201203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Känel, R.; Meister, R.E.; Stutz, M.; Kummer, P.; Arpagaus, A.; Huber, S.; Ehlert, U.; Wirtz, P.H. Effects of dark chocolate consumption on the prothrombotic response to acute psychosocial stress in healthy men. Thromb. Haemost. 2014, 112, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Biancardi, V.C.; Sharma, N.M. Connecting sympathetic and renin–angiotensin system overdrive in neurogenic hypertension through miRNA-181a. Hypertens. Res. 2020, 43, 1309–1310. [Google Scholar] [CrossRef] [PubMed]
- Rupp, H.; Jäger, B. The renin-angiotensin system and the sympathetic nervous system in hypertension and congestive heart failure: Implications for therapeutic interventions. J. Clin. Basic Cardiol. 2001, 4, 47–51. [Google Scholar]
- Reid, I.A. Interactions between ANG II, sympathetic nervous system, and baroreceptor reflexes in regulation of blood pressure. Am. J. Physiol. 1992, 262, E763–E778. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I. Signaling by the angiotensin-converting enzyme. Circ. Res. 2006, 98, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Alzamora, A.C.; Santos, R.A.S.; Campagnole-Santos, M.J. Hypotensive effect of ANG II and ANG-(1–7) at the caudal ventrolateral medulla involves different mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R1187–R1195. [Google Scholar] [CrossRef] [Green Version]
- Hrenak, J.; Simko, F. Renin-Angiotensin System: An important player in the pathogenesis of acute respiratory distress syndrome. Int. J. Mol. Sci. 2020, 21, 8038. [Google Scholar] [CrossRef]
- Zemlin, A.E.; Wiese, O.J. Coronavirus disease 2019 (COVID-19) and the renin-angiotensin system: A closer look at angiotensin-converting enzyme 2 (ACE2). Ann. Clin. Biochem. 2020, 57, 339–350. [Google Scholar] [CrossRef]
- Lovren, F.; Pan, Y.; Quan, A.; Teoh, H.; Wang, G.; Shukla, P.C.; Levitt, K.S.; Oudit, G.Y.; Al-Omran, M.; Stewart, D.J.; et al. Angiotensin converting enzyme-2 confers endothelial protection and attenuates atherosclerosis. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, H1377–H1384. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Takahashi, Y.; Yamaki, K. Inhibitory effect of catechin-related compounds on renin activity. Biomed. Res. 2013, 34, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Nakamura, T.; Munemasa, S.; Murata, Y.; Nakamura, Y. Galloylated catechins as potent inhibitors of angiotensin converting enzyme. Food Sci. Technol. Res. 2016, 22, 847–851. [Google Scholar] [CrossRef] [Green Version]
- Persson, I.A.; Josefsson, M.; Persson, K.; Andersson, R.G. Tea flavanols inhibit angiotensin-converting enzyme activity and increase nitric oxide production in human endothelial cells. J. Pharm. Pharmacol. 2006, 58, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Montes-Rivera, J.; Arellano-Mendoza, M.; Nájera, N.; Del Valle-Mondragón, L.; Villarreal, F.; Rubio-Gayosso, I.; Perez-Duran, J.; Meaney, E.; Ceballos, G. Effect of (−)-epicatechin on the modulation of progression markers of chronic renal damage in a 5/6 nephrectomy experimental model. Heliyon 2019, 5, e01512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchtaridi, M.; Fauzi, M.; Khairul Ikram, N.K.; Mohd Gazzali, A.; Wahab, H.A. Natural flavonoids as potential angiotensin-converting enzyme 2 inhibitors for anti-SARS-CoV-2. Molecules 2020, 25, 3980. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Said, A.; Tundis, R.; Rashed, K.; Statti, G.A.; Hufner, A.; Menichini, F. Inhibition of angiotensin converting enzyme (ACE) by flavonoids isolated from Ailanthus excelsa (Roxb) (Simaroubaceae). Phytother. Res. 2007, 21, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, L.; Castillo, J.; Quiñones, M.; Garcia-Vallvé, S.; Arola, L.; Pujadas, G.; Muguerza, B. Inhibition of angiotensin-converting enzyme activity by flavonoids: Structure-activity relationship studies. PLoS ONE 2012, 7, e49493. [Google Scholar] [CrossRef] [Green Version]
- Cacanyiova, S.; Krskova, K.; Zorad, S.; Frimmel, K.; Drobna, M.; Valaskova, Z.; Misak, A.; Golas, S.; Breza, J.; Breza, J., Jr. Arterial hypertension and plasma glucose modulate the vasoactive effects of nitroso-sulfide coupled signaling in human intrarenal arteries. Molecules 2020, 25, 2886. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Lüscher, T. COVID-19 is, in the end, an endothelial disease. Eur. Heart J. 2020, 41, 3038–3044. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Yao, X.Q.; Chan, F.L.; Lau, C.W.; Huang, Y. (−)-Epicatechin induces and modulates endothelium-dependent relaxation in isolated rat mesenteric artery rings. Acta Pharmacol. Sin. 2002, 23, 1188–1192. [Google Scholar]
- Huang, Y.; Yao, X.Q.; Tsang, S.Y.; Lau, C.W.; Chen, Z.Y. Role of endothelium/nitric oxide in vascular response to flavonoids and epicatechin. Acta Pharmacol. Sin. 2000, 21, 1119–1124. [Google Scholar] [PubMed]
- Ramirez-Sanchez, I.; Nogueira, L.; Moreno, A.; Murphy, A.; Taub, P.; Perkins, G.; Ceballos, G.M.; Hogan, M.; Malek, M.; Villarreal, F. Stimulatory effects of the flavanol (−)-epicatechin on cardiac angiogenesis: Additive effects with exercise. J. Cardiovasc. Pharmacol. 2012, 60, 429–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalopin, M.; Tesse, A.; Martinez, M.C.; Rognan, D.; Arnal, J.F.; Andriantsitohaina, R. Estrogen receptor alpha as a key target of red wine polyphenols action on the endothelium. PLoS ONE 2010, 5, e8554. [Google Scholar] [CrossRef]
- Moreno-Ulloa, A.; Mendez-Luna, D.; Beltran-Partida, E.; Castillo, C.; Guevara, G.; Ramirez-Sanchez, I.; Correa-Basurto, J.; Ceballos, G.; Villarreal, F. The effects of (−)-epicatechin on endothelial cells involve the G protein-coupled estrogen receptor (GPER). Pharmacol. Res. 2015, 100, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Flores, M.; Portilla-Martínez, A.; Cabrera-Pérez, F.; Nájera, N.; Meaney, E.; Villarreal, F.; Pérez-Durán, J.; Ceballos, G. PXR is a target of (−)-epicatechin in skeletal muscle. Heliyon 2020, 6, e05357. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Bi, R.; Hao, J.; Wang, S.; Huo, Y.; Demoz, R.M.; Banda, R.; Tian, S.; Xin, C.; Fu, M.; et al. A study on the protective effects of taxifolin on human umbilical vein endothelial cells and THP-1 cells damaged by hexavalent chromium: A probable mechanism for preventing cardiovascular disease induced by heavy metals. Food Funct. 2020, 11, 3851–3859. [Google Scholar] [CrossRef]
- Radosinska, J.; Vrbjar, N. The role of red blood cell deformability and Na,K-ATPase function in selected risk factors of cardiovascular diseases in humans: Focus on hypertension, diabetes mellitus and hypercholesterolemia. Physiol. Res. 2016, 65, S43–S54. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, K. Red blood cell abnormalities and hypertension. Hypertens. Res. 2020, 43, 72–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hacioglu, G.; Yalcin, O.; Bor-Kucukatay, M.; Ozkaya, G.; Baskurt, O.K. Red blood cell rheological properties in various rat hypertension models. Clin. Hemorheol. Microcirc. 2002, 26, 27–32. [Google Scholar] [PubMed]
- Chabanel, A.; Schachter, D.; Chien, S. Increased rigidity of red blood cell membrane in young spontaneously hypertensive rats. Hypertension 1987, 10, 603–607. [Google Scholar] [CrossRef] [Green Version]
- Lominadze, D.; Schuschke, D.A.; Joshua, I.G.; Dean, W.L. Increased ability of erythrocytes to aggregate in spontaneously hypertensive rats. Clin. Exp. Hypertens. 2002, 24, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Gkaliagkousi, E.; Passacquale, G.; Douma, S.; Zamboulis, C.; Ferro, A. Platelet activation in essential hypertension: Implications for antiplatelet treatment. Am. J. Hypertens. 2010, 23, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Guzmán, M.; Jiménez, R.; Sánchez, M.; Romero, M.; O’Valle, F.; Lopez-Sepulveda, R.; Quintela, A.M.; Galindo, P.; Zarzuelo, M.J.; Bailón, E.; et al. Chronic (−)-epicatechin improves vascular oxidative and inflammatory status but not hypertension in chronic nitric oxide-deficient rats. Br. J. Nutr. 2011, 106, 1337–1348. [Google Scholar] [CrossRef] [Green Version]
- Prince, P.D.; Fraga, C.G.; Galleano, M. (−)-Epicatechin administration protects kidneys against modifications induced by short-term l-NAME treatment in rats. Food Funct. 2020, 11, 318–327. [Google Scholar] [CrossRef]
- Satoru, S.; Yui, K.; Mayu, M.; Hiroyuki, S.; Riko, H.; Kohtaroh, G.; Kohhei, U.; Yohko, F. Taxifolin Potently Diminishes Levels of Reactive Oxygen Species in Living Cells Possibly by Scavenging Peroxyl Radicals. Am. J. Pharmacol. Toxicol. 2018, 13, 1–6. [Google Scholar] [CrossRef]
- Litterio, M.C.; Jaggers, G.; Sagdicoglu, C.G.; Adamo, A.M.; Costa, M.A.; Oteiza, P.I.; Fraga, C.G.; Galleano, M. Blood pressure-lowering effect of dietary (−)-epicatechin administration in L-NAME-treated rats is associated with restored nitric oxide levels. Free Radic. Biol. Med. 2012, 53, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Rhee, M.H.; Endale, M.; Kamruzzaman, S.M.; Lee, W.M.; Park, H.J.; Yoo, M.J.; Cho, J.Y. Taxifolin inhibited the nitric oxide production and expression of pro-inflammatory cytokine mRNA in lipopolysaccharide-stimulated RAW 264.7 cells. J. Exp. Biomed. Sci. 2008, 14, 147–155. [Google Scholar]
- Slashcheva, G.A.; Rykov, V.A.; Lobanov, A.V.; Murashev, A.N.; Kim, Y.A.; Arutyunyan, T.V.; Korystova, A.F.; Kublik, L.N.; Levitman, M.K.; Shaposhnikona, V.V.; et al. Dihydroquercetin does not affect age-dependent increase in blood pressure and angiotensin-converting enzyme activity in the aorta of hypertensive rats. Bull. Exp. Biol. Med. 2016, 161, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Arutyunyan, T.V.; Korystova, A.F.; Kublik, L.N.; Levitman, M.; Shaposhnikova, V.V.; Korystov, Y.N. Effects of taxifolin on the activity of angiotensin-converting enzyme and reactive oxygen and nitrogen species in the aorta of aging rats and rats treated with the nitric oxide synthase inhibitor and dexamethasone. Age 2013, 35, 2089–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, P.D.; Rodríguez Lanzi, C.; Fraga, C.G.; Galleano, M. Dietary (−)-epicatechin affects NF-κB activation and NADPH oxidases in the kidney cortex of high-fructose-fed rats. Food Funct. 2019, 10, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; De Simone, G.; Dominiczak, A.; et al. 2018 ESC/ESH Guidelines for the management of arterial hypertension: The task force for the management of arterial hypertension of the European Society of Cardiology and the European Society of Hypertension. J. Hypertens. 2018, 36, 1953–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernatova, I.; Conde, M.V.; Kopincova, J.; González, M.C.; Puzserova, A.; Arribas, S.M. Endothelial dysfunction in spontaneously hypertensive rats: Focus on methodological aspects. J. Hypertens. Suppl. 2009, 27, S27–S31. [Google Scholar] [CrossRef]
- Fuchs, L.C.; Hoque, A.M.; Clarke, N.L. Vascular and hemodynamic effects of behavioral stress in borderline hypertensive and Wistar-Kyoto rats. Am. J. Physiol. 1998, 274, R375–R382. [Google Scholar] [CrossRef] [PubMed]
- Slezak, P.; Puzserova, A.; Balis, P.; Sestakova, N.; Majzunova, M.; Dovinova, I.; Kluknavsky, M.; Bernatova, I. Genotype-related effect of crowding stress on blood pressure and vascular function in young female rats. BioMed Res. Int. 2014, 2014, 413629. [Google Scholar] [CrossRef] [Green Version]
- Connolly, K.; Jackson, D.; Batacan, R.; Fenning, A. Epicatechin improves lipid profile and oxidative stress status, but does not reduce abdominal fat or blood pressure in an obese SHR model of metabolic syndrome. Heart Lung Circ. 2015, 24, S187. [Google Scholar] [CrossRef] [Green Version]
- Bernatova, I.; Pechanova, O.; Babal, P.; Kysela, S.; Stvrtina, S.; Andriantsitohaina, R. Wine polyphenols improve cardiovascular remodeling and vascular function in NO-deficient hypertension. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H942–H948. [Google Scholar] [CrossRef] [Green Version]
- Manning, R.D., Jr.; Hu, L.; Mizelle, H.L.; Montani, J.P.; Norton, M.W. Cardiovascular responses to long-term blockade of nitric oxide synthesis. Hypertension 1993, 22, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Paulis, L.; Zicha, J.; Kunes, J.; Hojna, S.; Behuliak, M.; Celec, P.; Kojsova, S.; Pechanova, O.; Simko, F. Regression of L-NAME-induced hypertension: The role of nitric oxide and endothelium-derived constricting factor. Hypertens. Res. 2008, 31, 793–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, C.; Leung, S.W.; Vanhoutte, P.M.; Man, R.Y. Chronic inhibition of nitric-oxide synthase potentiates endothelium-dependent contractions in the rat aorta by augmenting the expression of cyclooxygenase-2. J. Pharmacol. Exp. Ther. 2010, 334, 373–380. [Google Scholar] [CrossRef]
- Liskova, S.; Petrova, M.; Karen, P.; Kunes, J.; Zicha, J. Effects of aging and hypertension on the participation of endothelium-derived constricting factor (EDCF) in norepinephrine-induced contraction of rat femoral artery. Eur. J. Pharmacol. 2011, 667, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, M.; Shinozaki, M.; Hirakata, H.; Tamaki, K.; Hirano, T.; Tokumoto, M.; Goto, H.; Okuda, S.; Fujishima, M. Locally activated renin-angiotensin system associated with TGF-beta1 as a major factor for renal injury induced by chronic inhibition of nitric oxide synthase in rats. J. Am. Soc. Nephrol. 2000, 11, 616–624. [Google Scholar] [PubMed]
- Majzunova, M.; Kvandova, M.; Berenyiova, A.; Balis, P.; Dovinova, I.; Cacanyiova, S. Chronic NOS inhibition affects oxidative state and antioxidant response differently in the kidneys of young normotensive and hypertensive rats. Oxid. Med. Cell. Longev. 2019, 2019, 5349398. [Google Scholar] [CrossRef] [PubMed]
- Potue, P.; Wunpathe, C.; Maneesai, P.; Kukongviriyapan, U.; Prachaney, P.; Pakdeechote, P. Nobiletin alleviates vascular alterations through modulation of Nrf-2/HO-1 and MMP pathways in L-NAME induced hypertensive rats. Food Funct. 2019, 10, 1880–1892. [Google Scholar] [CrossRef]
- Basting, T.; Lazartigues, E. DOCA-Salt Hypertension: An update. Curr. Hypertens. Rep. 2017, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.V.; Kiat, H. The mechanisms underlying fructose-induced hypertension: A review. J. Hypertens. 2015, 33, 912–920. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.H.; Chu, C.H.; Wen, S.W.; Lai, C.C.; Cheng, P.W.; Tseng, C.J. Excessive fructose intake impairs baroreflex sensitivity and led to elevated blood pressure in rats. Nutrients 2019, 11, 2581. [Google Scholar] [CrossRef] [Green Version]
- Chou, C.L.; Pang, C.Y.; Lee, T.J.; Fang, T.C. Direct renin inhibitor prevents and ameliorates insulin resistance, aortic endothelial dysfunction and vascular remodeling in fructose-fed hypertensive rats. Hypertens. Res. 2013, 36, 123–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemančíková, A.; Török, J. Effect of perivascular adipose tissue on arterial adrenergic contractions in normotensive and hypertensive rats with high fructose intake. Physiol. Res. 2017, 66, S537–S544. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biological Effects | (−)-Epicatechin | (+)-Taxifolin and/or (−)-Taxifolin |
|---|---|---|
| Antihypertensive | [38,58,59,60,61,62,63,64] | [65,66] |
| Vascular | [38,58,59,67,68,69,70,71] | [66] |
| Cardioprotective | [72,73,74] | [75,76] |
| Antidiabetic, antihyperglycemic, or antidyslipidemic | [77,78,79,80,81] | [82] |
| Hepatoprotective | [79] | [83] |
| Respiratory | [84] | [85] |
| Antiinflammatory | [86,87,88] | [89,90] |
| Improved erythrocyte deformability | [38] | [91] |
| Antiaggregatory, antithrombotic, or anticoagulant | [38,92,93] | [64,94,95,96] |
| Antifibrotic | [84] | [75,97] |
| Antiviral | [13,14] | [17] |
| Cellular or Molecular Mechanisms | (−)-Epicatechin | (+)-Taxifolin and/or (−)-Taxifolin |
|---|---|---|
| Chelation of Fe2+, Cu2+, Zn2+ | [107] | [108] |
| ROS scavenging | [105,106] | [108] |
| Alterations of iron metabolism | [60,72] | [40,83] |
| Activation of antioxidant enzymes (SOD, CAT, or GPx) | [181,182] | [76,183] |
| Reduction of NOX or XO | [61,62,88,109,182,184] | [96,109,110] |
| Elevation of endothelial NO | [38,61,62,68,181] | [66] |
| Reduction of inducible NOS-produced NO | [87,90,131] | [90,129,130,131,185] |
| Arginase inhibition | [58,132,133] | [133] |
| Reduction of ACE | [160,161] | [65,186,187] |
| Nrf2 induction | [61,121] | [76,121,122] |
| NFκB reduction | [87,137,188] | [140] |
| Inflammasome reduction | [141,142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernatova, I.; Liskova, S. Mechanisms Modified by (−)-Epicatechin and Taxifolin Relevant for the Treatment of Hypertension and Viral Infection: Knowledge from Preclinical Studies. Antioxidants 2021, 10, 467. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030467
Bernatova I, Liskova S. Mechanisms Modified by (−)-Epicatechin and Taxifolin Relevant for the Treatment of Hypertension and Viral Infection: Knowledge from Preclinical Studies. Antioxidants. 2021; 10(3):467. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030467
Chicago/Turabian StyleBernatova, Iveta, and Silvia Liskova. 2021. "Mechanisms Modified by (−)-Epicatechin and Taxifolin Relevant for the Treatment of Hypertension and Viral Infection: Knowledge from Preclinical Studies" Antioxidants 10, no. 3: 467. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10030467