
Noise-Induced Vascular Dysfunction, Oxidative Stress, and Inflammation Are Improved by Pharmacological Modulation of the NRF2/HO-1 Axis

 , , , , ,
, , , , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
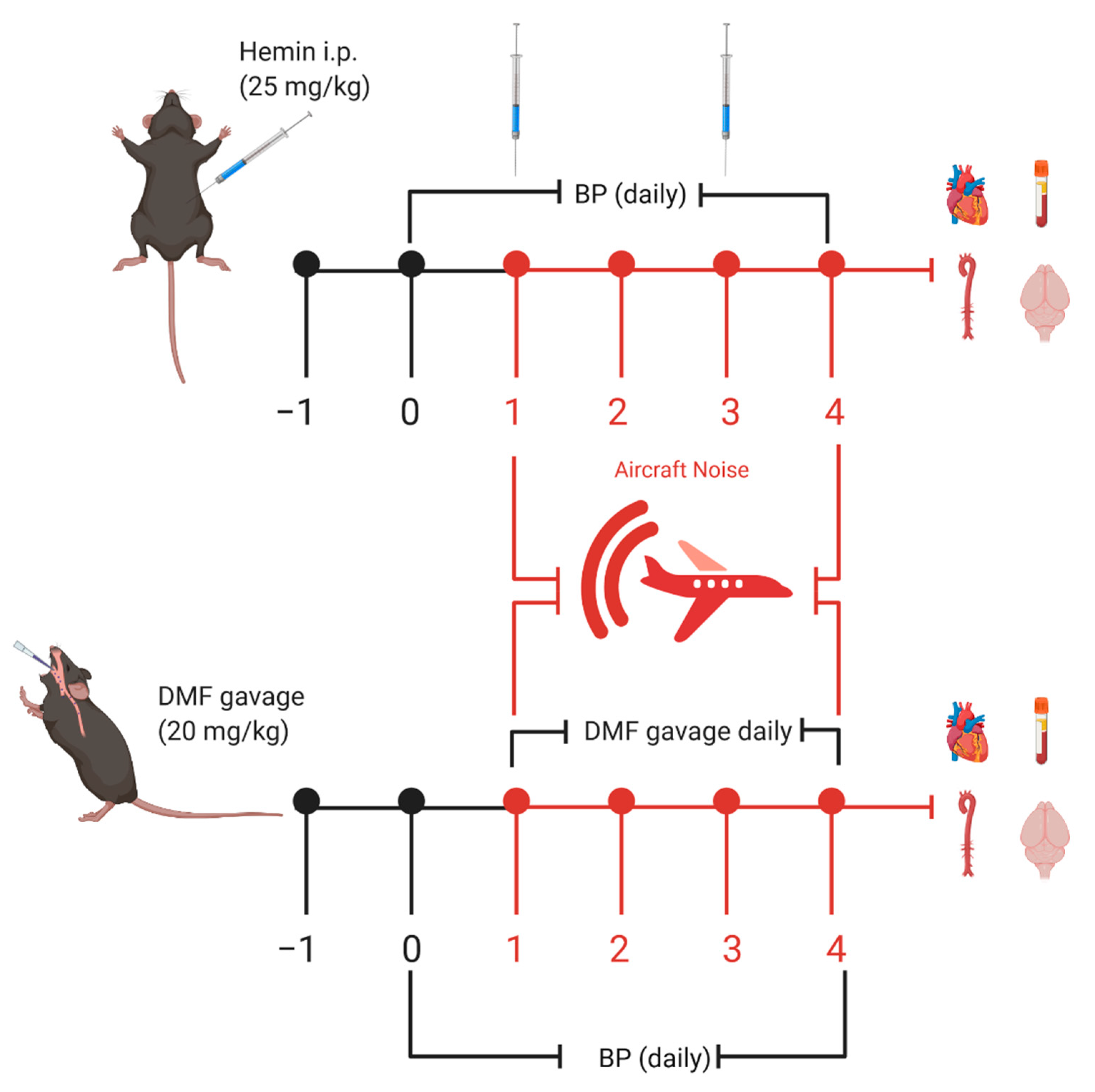
2.1. Animals
2.2. Sacrifice of Animals, Organ Removal, and Sample Preparation
2.3. Hemin Treatment
2.4. Dimethyl Fumarate Treatment
2.5. Noise Exposure
2.6. Noninvasive Blood Pressure (NIBP)
2.7. Isometric Tension Studies
2.8. Detection of Oxidative Stress and Inflammation in Cortical, Cardiac, and Aortic Tissues
2.9. Detection of Bilirubin by HPLC Method Plasma
2.10. Western Blotting and Dot Blot Analysis
2.11. Quantitative Reverse Transcription Real-Time PCR (qRT-PCR)
2.12. Statistical Analysis
3. Results
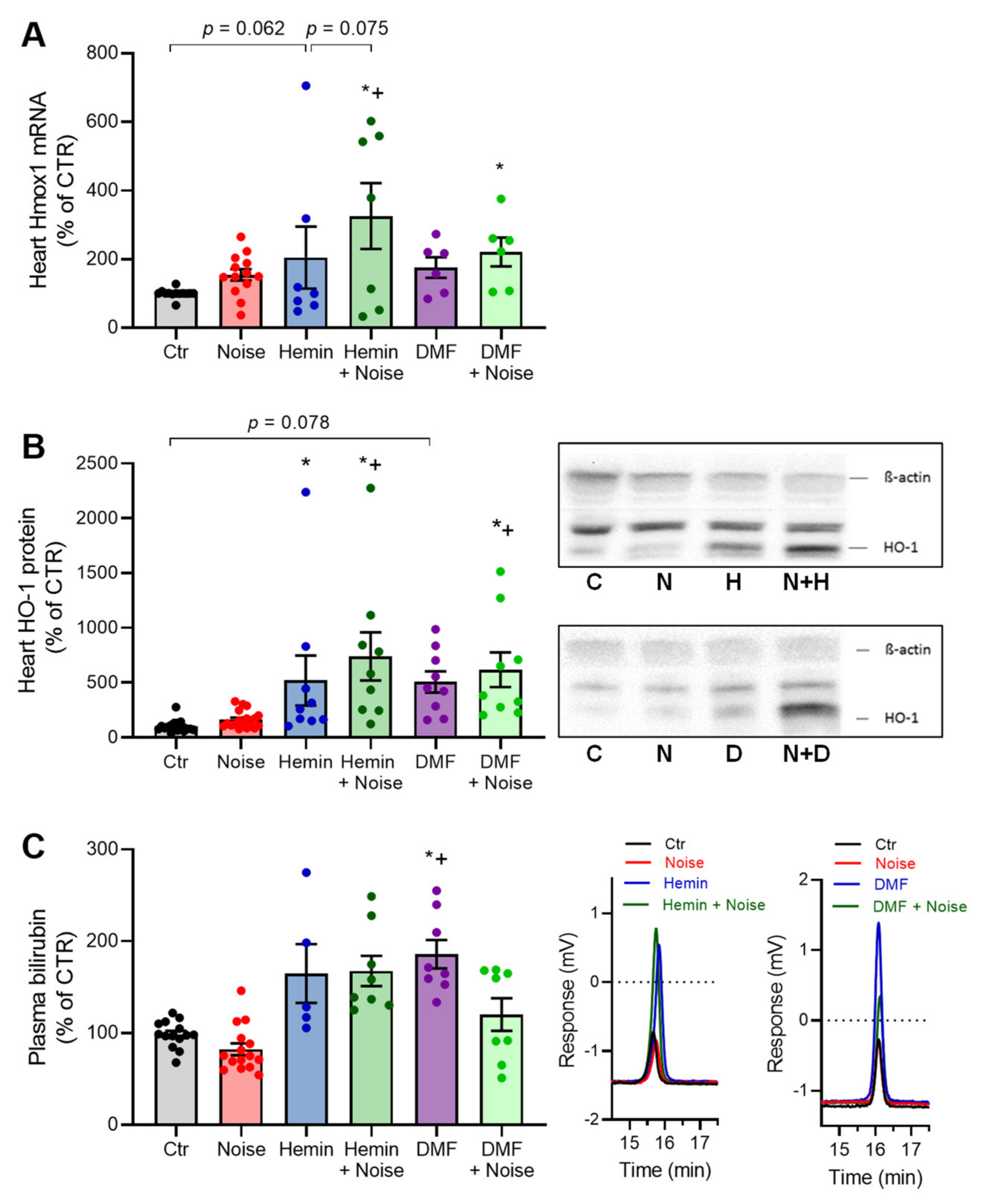
3.1. Effects of Aircraft Noise, Hemin, and DMF on HO-1 Expression and Its Subproduct Bilirubin
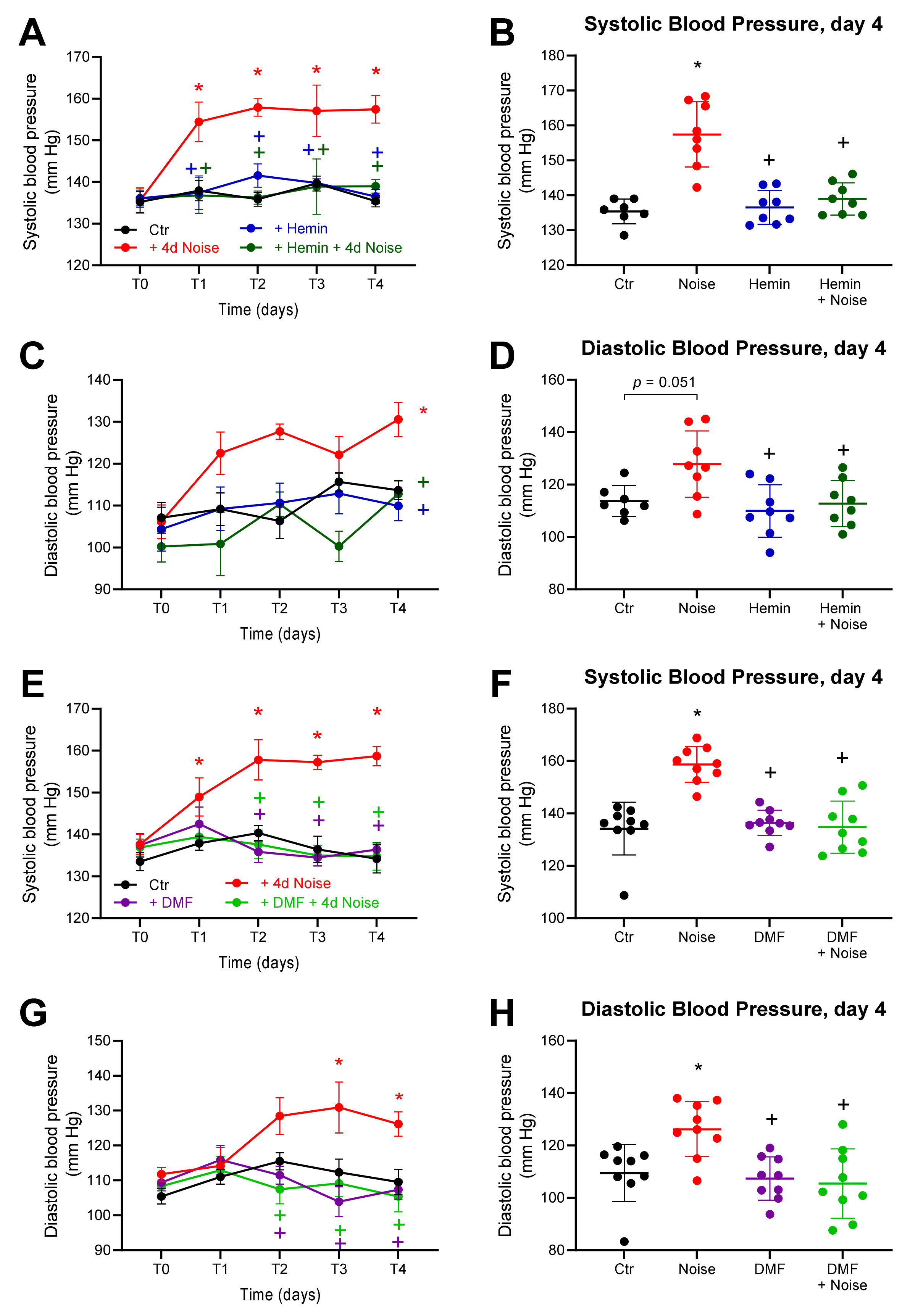
3.2. Effects of Aircraft Noise and HO-1 Inducer and NRF2 Activator on Blood Pressure
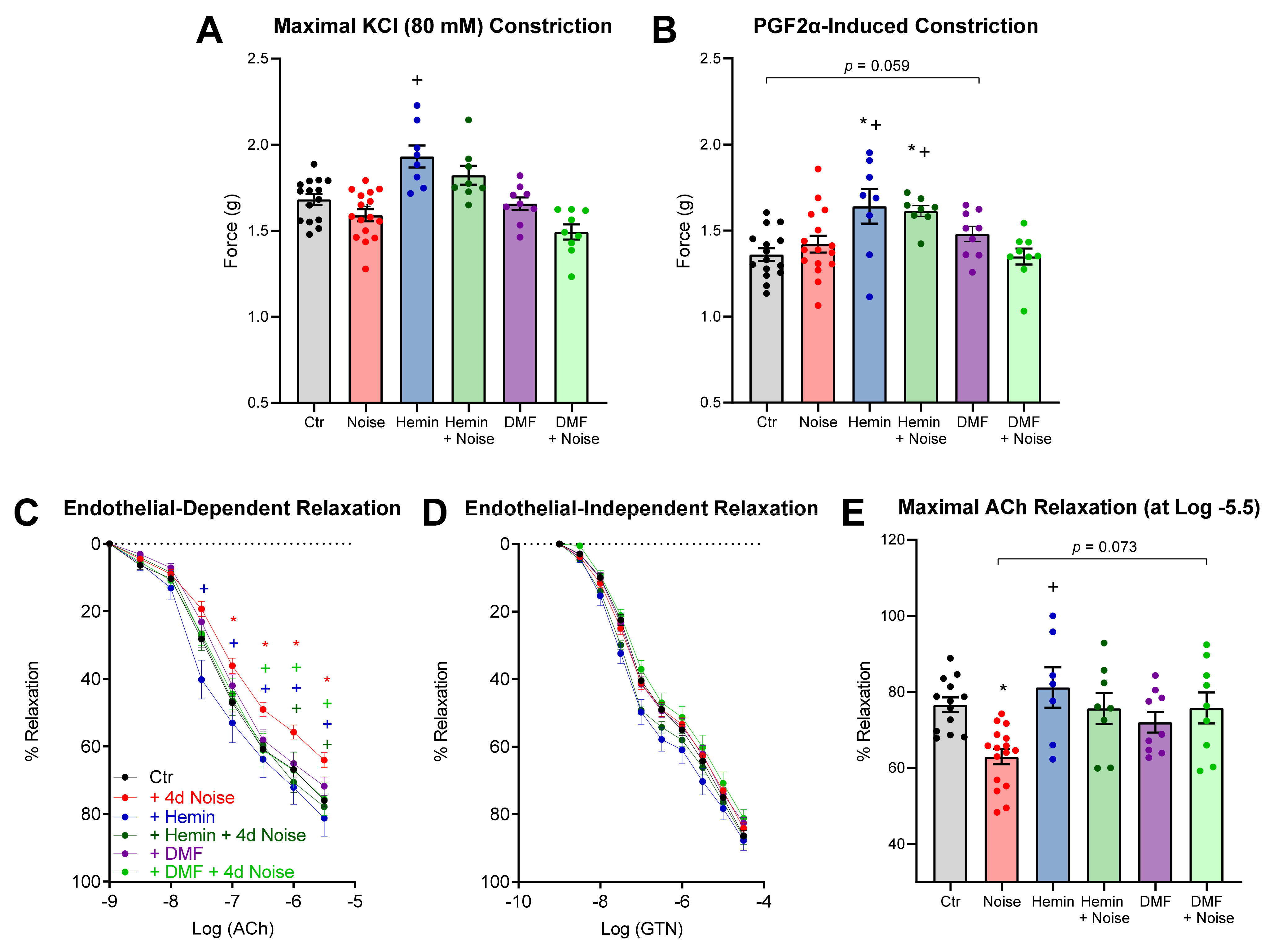
3.3. Vascular Function Is Protected by Induction of HO-1 and Activation of NRF2
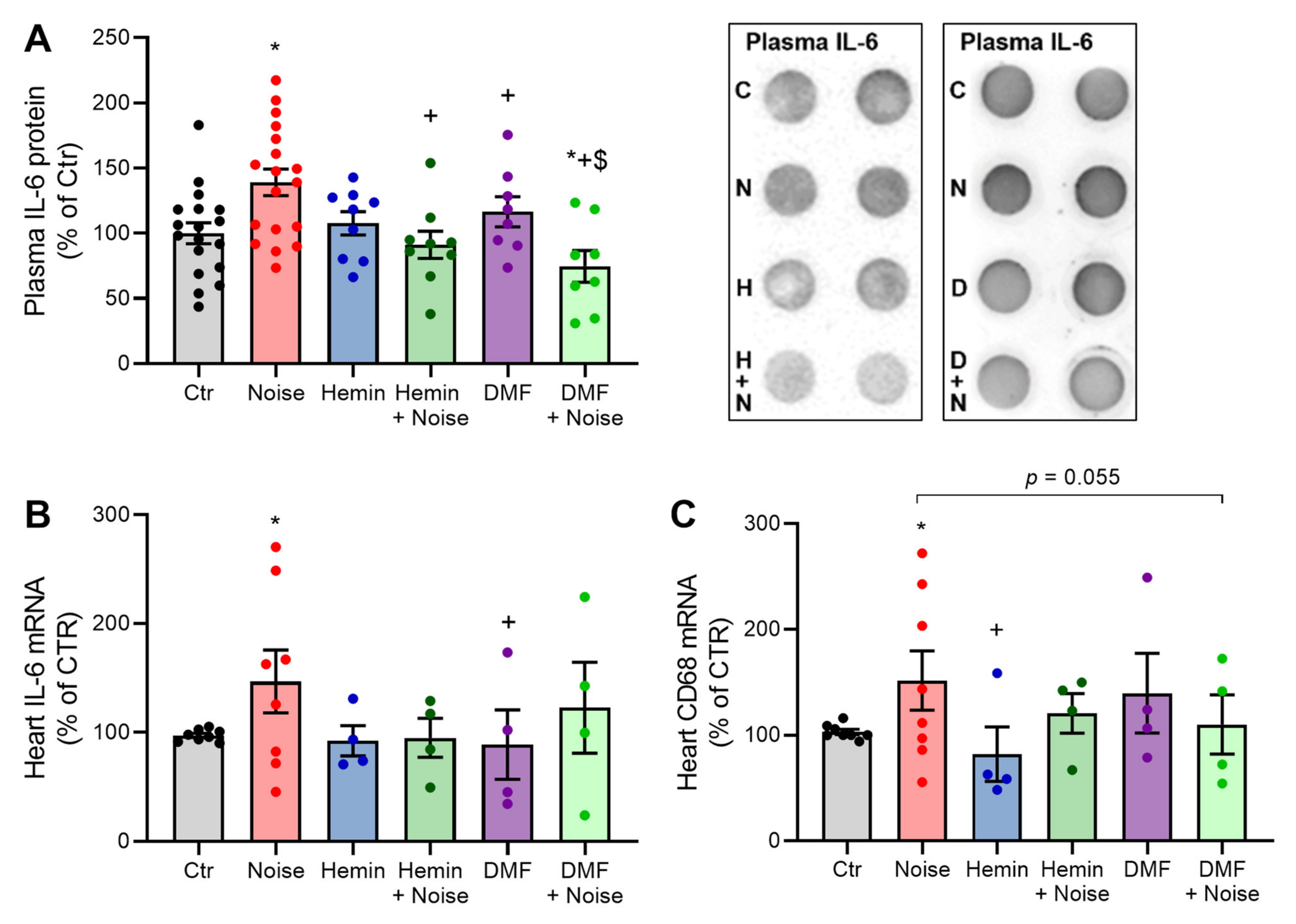
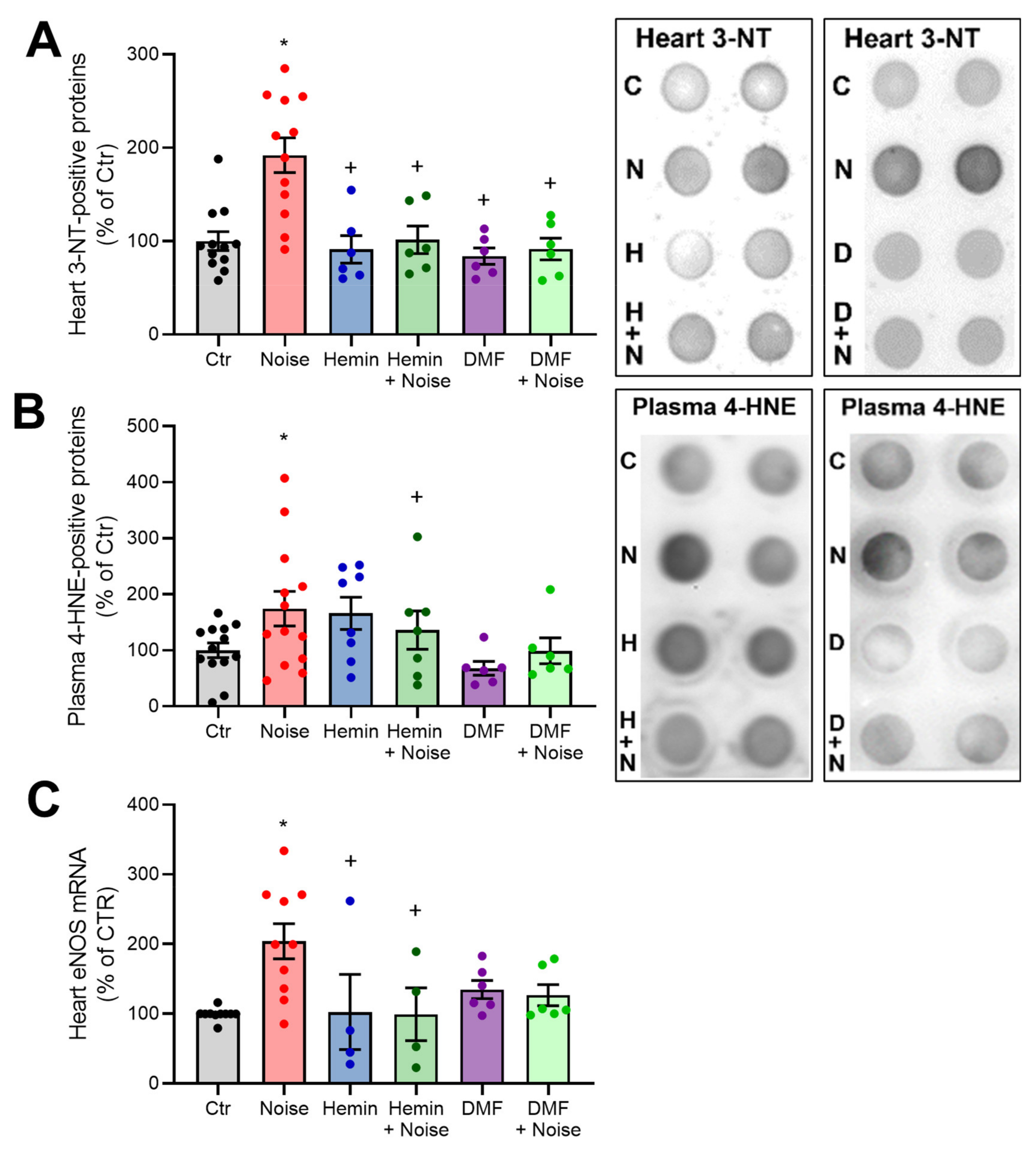
3.4. Induction of HO-1 and Activation of NRF2 Maintains Inflammatory Parameters at Basal Levels
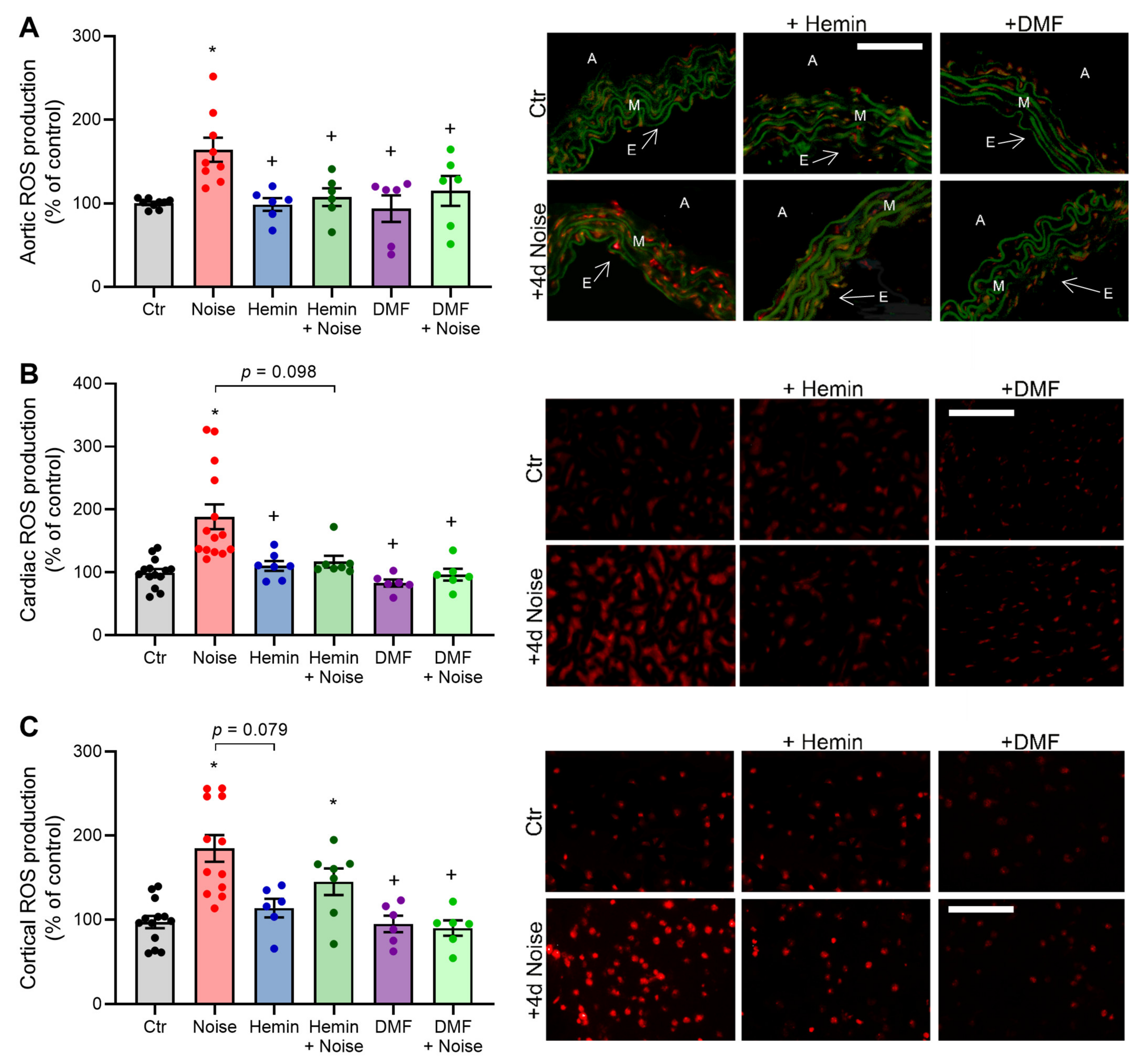
3.5. Effects of Aircraft Noise and Induction of HO-1 and Activation of NRF2 on ROS Production in Aortic, Cardiac, and Cerebral Tissues
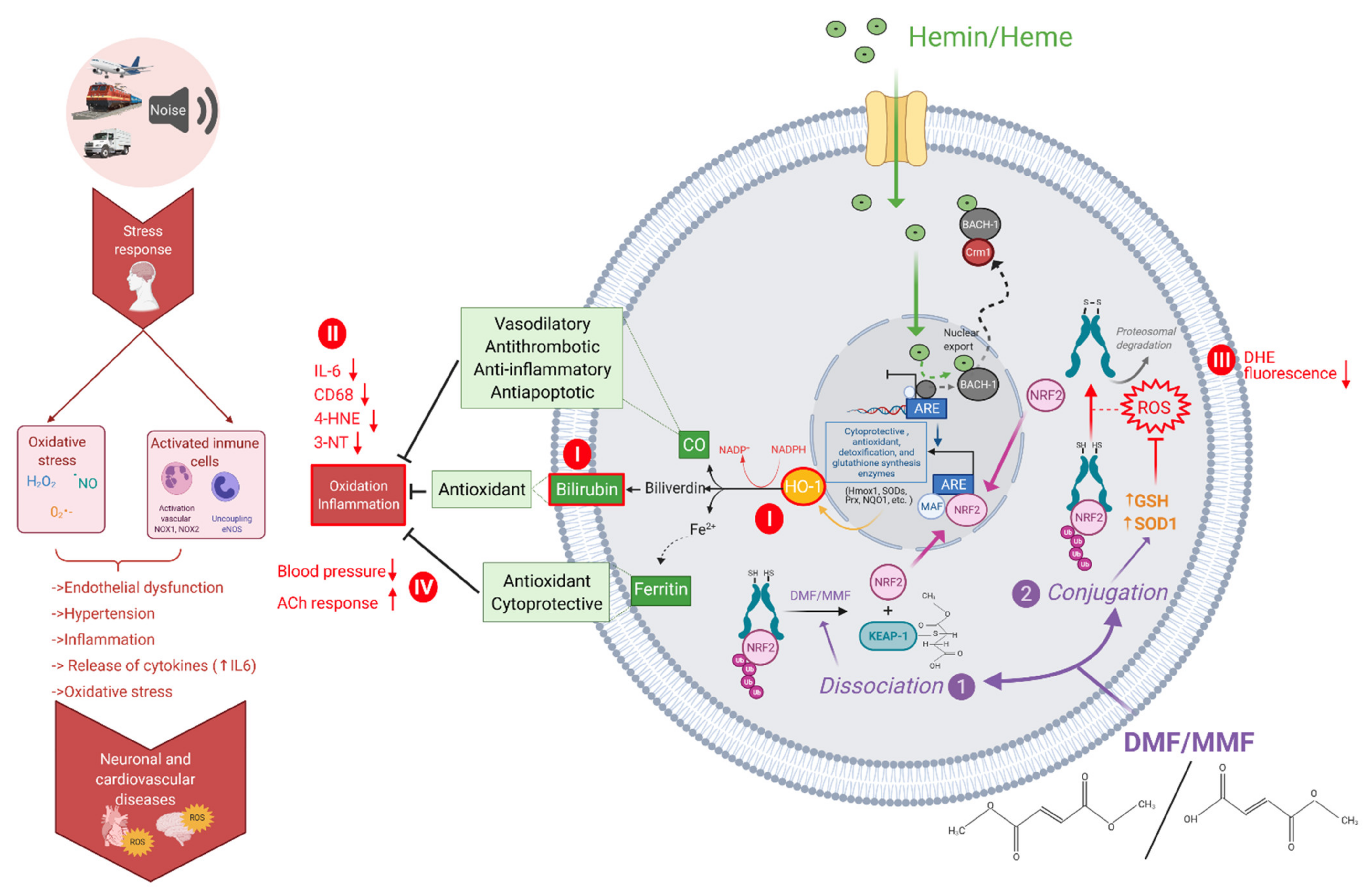
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
List of Abbreviations
| 3-NT | 3-nitrotyrosine |
| 4-HNE | 4-hydroxynonenal |
| ACh | acetylcholine |
| CD68 | cluster of differentiation 68 |
| dB(A) | decibel |
| DHE | dihydroethidium |
| DMF | dimethyl fumarate |
| DMSO | dimethyl sulfoxide |
| eNOS | endothelial •NO synthase (type 3) |
| GTN | nitroglycerine |
| HO-1 | heme oxygenase-1 |
| IL | interleukin |
| IP | intraperitoneal injection |
| KCl | potassium chloride |
| Leq | equivalent continuous sound pressure level |
| NIBP | noninvasive blood pressure |
| NO | nitric oxide |
| NOX | NADPH oxidase |
| NRF2 | nuclear-factor erythroid 2-related factor 2 |
| p.o. | per os, oral administration |
| PBS | phosphate-buffered saline |
| ROS | reactive oxygen species |
| qRT-PCR | quantitative reverse transcription real-time polymerase chain reaction |
| SPL | sound pressure level |
| TBT | TATA box-binding protein |
| VPR | volume–pressure recording cuff |
References
- Daiber, A.; Steven, S.; Weber, A.; Shuvaev, V.V.; Muzykantov, V.R.; Laher, I.; Li, H.; Lamas, S.; Munzel, T. Targeting vascular (endothelial) dysfunction. Br J. Pharmacol. 2017, 174, 1591–1619. [Google Scholar] [CrossRef]
- Kalsch, H.; Hennig, F.; Moebus, S.; Mohlenkamp, S.; Dragano, N.; Jakobs, H.; Memmesheimer, M.; Erbel, R.; Jockel, K.H.; Hoffmann, B.; et al. Are air pollution and traffic noise independently associated with atherosclerosis: The heinz nixdorf recall study. Eur. Heart J. 2014, 35, 853–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, F.; Kolle, K.; Kreuder, K.; Schnorbus, B.; Wild, P.; Hechtner, M.; Binder, H.; Gori, T.; Munzel, T. Nighttime aircraft noise impairs endothelial function and increases blood pressure in patients with or at high risk for coronary artery disease. Clin. Res. Cardiol. 2015, 104, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munzel, T.; Daiber, A.; Steven, S.; Tran, L.P.; Ullmann, E.; Kossmann, S.; Schmidt, F.P.; Oelze, M.; Xia, N.; Li, H.; et al. Effects of noise on vascular function, oxidative stress, and inflammation: Mechanistic insight from studies in mice. Eur. Heart J. 2017, 38, 2838–2849. [Google Scholar] [CrossRef] [Green Version]
- Kroller-Schon, S.; Daiber, A.; Steven, S.; Oelze, M.; Frenis, K.; Kalinovic, S.; Heimann, A.; Schmidt, F.P.; Pinto, A.; Kvandova, M.; et al. Crucial role for nox2 and sleep deprivation in aircraft noise-induced vascular and cerebral oxidative stress, inflammation, and gene regulation. Eur. Heart J. 2018, 39, 3528–3539. [Google Scholar] [CrossRef] [PubMed]
- Ponka, P. Cell biology of heme. Am. J. Med. Sci 1999, 318, 241–256. [Google Scholar] [CrossRef]
- Maxwell, S.R.; Lip, G.Y. Free radicals and antioxidants in cardiovascular disease. Br. J. Clin. Pharmacol. 1997, 44, 307–317. [Google Scholar] [CrossRef]
- Victor, V.M.; Rocha, M. Targeting antioxidants to mitochondria: A potential new therapeutic strategy for cardiovascular diseases. Curr. Pharm. Des. 2007, 13, 845–863. [Google Scholar] [CrossRef]
- Schuhmacher, S.; Wenzel, P.; Schulz, E.; Oelze, M.; Mang, C.; Kamuf, J.; Gori, T.; Jansen, T.; Knorr, M.; Karbach, S.; et al. Pentaerythritol tetranitrate improves angiotensin ii-induced vascular dysfunction via induction of heme oxygenase-1. Hypertension 2010, 55, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Balla, G.; Jacob, H.S.; Eaton, J.W.; Belcher, J.D.; Vercellotti, G.M. Hemin: A possible physiological mediator of low density lipoprotein oxidation and endothelial injury. Arterioscler. Thromb. 1991, 11, 1700–1711. [Google Scholar] [CrossRef] [Green Version]
- Choi, A.M. Heme oxygenase-1 protects the heart. Circ. Res. 2001, 89, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Zabalgoitia, M.; Colston, J.T.; Reddy, S.V.; Holt, J.W.; Regan, R.F.; Stec, D.E.; Rimoldi, J.M.; Valente, A.J.; Chandrasekar, B. Carbon monoxide donors or heme oxygenase-1 (ho-1) overexpression blocks interleukin-18-mediated nf-kappa b-pten-dependent human cardiac endothelial cell death. Free Radic. Biol. Med. 2008, 44, 284–298. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.; Rossmann, H.; Muller, C.; Kossmann, S.; Oelze, M.; Schulz, A.; Arnold, N.; Simsek, C.; Lagrange, J.; Klemz, R.; et al. Heme oxygenase-1 suppresses a pro-inflammatory phenotype in monocytes and determines endothelial function and arterial hypertension in mice and humans. Eur. Heart J. 2015, 36, 3437–3446. [Google Scholar] [CrossRef] [Green Version]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of nrf2/ho-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell Mol. Life Sci 2016, 73, 3221–3247. [Google Scholar] [CrossRef] [Green Version]
- Belcher, J.D.; Chen, C.; Nguyen, J.; Zhang, P.; Abdulla, F.; Nguyen, P.; Killeen, T.; Xu, P.; O’Sullivan, G.; Nath, K.A.; et al. Control of oxidative stress and inflammation in sickle cell disease with the nrf2 activator dimethyl fumarate. Antioxid Redox Signal. 2017, 26, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Havrdova, E.; Giovannoni, G.; Gold, R.; Fox, R.J.; Kappos, L.; Phillips, J.T.; Okwuokenye, M.; Marantz, J.L. Effect of delayed-release dimethyl fumarate on no evidence of disease activity in relapsing-remitting multiple sclerosis: Integrated analysis of the phase iii define and confirm studies. Eur. J. Neurol 2017, 24, 726–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, M.; Thierry, A.; Delbauve, S.; Preyat, N.; Soares, M.P.; Roumeguere, T.; Leo, O.; Flamand, V.; Le Moine, A.; Hougardy, J.M. Specific expression of heme oxygenase-1 by myeloid cells modulates renal ischemia-reperfusion injury. Sci. Rep. 2017, 7, 197. [Google Scholar] [CrossRef] [PubMed]
- Campolo, M.; Casili, G.; Biundo, F.; Crupi, R.; Cordaro, M.; Cuzzocrea, S.; Esposito, E. The neuroprotective effect of dimethyl fumarate in an mptp-mouse model of parkinson’s disease: Involvement of reactive oxygen species/nuclear factor-kappab/nuclear transcription factor related to nf-e2. Antioxid Redox Signal. 2017, 27, 453–471. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.G.; Parrish, J.L.; Hughes, L.F.; Toth, L.A.; Caspary, D.M. Hearing in laboratory animals: Strain differences and nonauditory effects of noise. Comp. Med. 2005, 55, 12–23. [Google Scholar] [PubMed]
- Munzel, T.; Giaid, A.; Kurz, S.; Stewart, D.J.; Harrison, D.G. Evidence for a role of endothelin 1 and protein kinase c in nitroglycerin tolerance. Proc. Natl. Acad. Sci. USA 1995, 92, 5244–5248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelze, M.; Knorr, M.; Kroller-Schon, S.; Kossmann, S.; Gottschlich, A.; Rummler, R.; Schuff, A.; Daub, S.; Doppler, C.; Kleinert, H.; et al. Chronic therapy with isosorbide-5-mononitrate causes endothelial dysfunction, oxidative stress, and a marked increase in vascular endothelin-1 expression. Eur. Heart J. 2013, 34, 3206–3216. [Google Scholar] [CrossRef] [Green Version]
- Oelze, M.; Daiber, A.; Brandes, R.P.; Hortmann, M.; Wenzel, P.; Hink, U.; Schulz, E.; Mollnau, H.; von Sandersleben, A.; Kleschyov, A.L.; et al. Nebivolol inhibits superoxide formation by nadph oxidase and endothelial dysfunction in angiotensin ii-treated rats. Hypertension 2006, 48, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, P.; Schulz, E.; Oelze, M.; Muller, J.; Schuhmacher, S.; Alhamdani, M.S.; Debrezion, J.; Hortmann, M.; Reifenberg, K.; Fleming, I.; et al. At1-receptor blockade by telmisartan upregulates gtp-cyclohydrolase i and protects enos in diabetic rats. Free Radic Biol. Med. 2008, 45, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Oelze, M.; Kroller-Schon, S.; Steven, S.; Lubos, E.; Doppler, C.; Hausding, M.; Tobias, S.; Brochhausen, C.; Li, H.; Torzewski, M.; et al. Glutathione peroxidase-1 deficiency potentiates dysregulatory modifications of endothelial nitric oxide synthase and vascular dysfunction in aging. Hypertension 2014, 63, 390–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, T.; Hortmann, M.; Oelze, M.; Opitz, B.; Steven, S.; Schell, R.; Knorr, M.; Karbach, S.; Schuhmacher, S.; Wenzel, P.; et al. Conversion of biliverdin to bilirubin by biliverdin reductase contributes to endothelial cell protection by heme oxygenase-1-evidence for direct and indirect antioxidant actions of bilirubin. J. Mol. Cell Cardiol 2010, 49, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Hanf, A.; Oelze, M.; Manea, A.; Li, H.; Munzel, T.; Daiber, A. The anti-cancer drug doxorubicin induces substantial epigenetic changes in cultured cardiomyocytes. Chem. Biol. Interact. 2019, 313, 108834. [Google Scholar] [CrossRef]
- Christensen, J.S.; Raaschou-Nielsen, O.; Tjonneland, A.; Overvad, K.; Nordsborg, R.B.; Ketzel, M.; Sorensen, T.; Sorensen, M. Road traffic and railway noise exposures and adiposity in adults: A cross-sectional analysis of the danish diet, cancer, and health cohort. Environ. Health Perspect. 2016, 124, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Steven, S.; Frenis, K.; Kalinovic, S.; Kvandova, M.; Oelze, M.; Helmstadter, J.; Hahad, O.; Filippou, K.; Kus, K.; Trevisan, C.; et al. Exacerbation of adverse cardiovascular effects of aircraft noise in an animal model of arterial hypertension. Redox Biol. 2020, 34, 101515. [Google Scholar] [CrossRef]
- Griendling, K.K.; FitzGerald, G.A. Oxidative stress and cardiovascular injury: Part i: Basic mechanisms and in vivo monitoring of ros. Circulation 2003, 108, 1912–1916. [Google Scholar] [CrossRef] [Green Version]
- Griendling, K.K.; FitzGerald, G.A. Oxidative stress and cardiovascular injury: Part ii: Animal and human studies. Circulation 2003, 108, 2034–2040. [Google Scholar] [CrossRef]
- Harrison, D.; Griendling, K.K.; Landmesser, U.; Hornig, B.; Drexler, H. Role of oxidative stress in atherosclerosis. Am. J. Cardiol. 2003, 91, 7A–11A. [Google Scholar] [CrossRef]
- Jay, D.; Hitomi, H.; Griendling, K.K. Oxidative stress and diabetic cardiovascular complications. Free Radic Biol. Med. 2006, 40, 183–192. [Google Scholar] [CrossRef]
- Herzog, J.; Schmidt, F.P.; Hahad, O.; Mahmoudpour, S.H.; Mangold, A.K.; Garcia Andreo, P.; Prochaska, J.; Koeck, T.; Wild, P.S.; Sorensen, M.; et al. Acute exposure to nocturnal train noise induces endothelial dysfunction and pro-thromboinflammatory changes of the plasma proteome in healthy subjects. Basic Res. Cardiol. 2019, 114, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, F.P.; Herzog, J.; Schnorbus, B.; Ostad, M.A.; Lasetzki, L.; Hahad, O.; Schafers, G.; Gori, T.; Sorensen, M.; Daiber, A.; et al. The impact of aircraft noise on vascular and cardiac function in relation to noise event number - a randomized trial. Cardiovasc. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.P.; Basner, M.; Kroger, G.; Weck, S.; Schnorbus, B.; Muttray, A.; Sariyar, M.; Binder, H.; Gori, T.; Warnholtz, A.; et al. Effect of nighttime aircraft noise exposure on endothelial function and stress hormone release in healthy adults. Eur. Heart J. 2013, 34, 3508a–3514a. [Google Scholar] [CrossRef]
- Munzel, T.; Daiber, A.; Ullrich, V.; Mulsch, A. Vascular consequences of endothelial nitric oxide synthase uncoupling for the activity and expression of the soluble guanylyl cyclase and the cgmp-dependent protein kinase. Arterioscler Thromb. Vasc. Biol. 2005, 25, 1551–1557. [Google Scholar] [CrossRef]
- Daiber, A.; Di Lisa, F.; Oelze, M.; Kroller-Schon, S.; Steven, S.; Schulz, E.; Munzel, T. Crosstalk of mitochondria with nadph oxidase via reactive oxygen and nitrogen species signalling and its role for vascular function. Br. J. Pharmacol. 2015, 174, 1670–1689. [Google Scholar] [CrossRef] [Green Version]
- Babisch, W. The noise/stress concept, risk assessment and research needs. Noise Health 2002, 4, 1–11. [Google Scholar]
- Daiber, A.; Kroller-Schon, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Vujacic-Mirski, K.; Kuntic, M.; Bayo Jimenez, M.T.; Helmstadter, J.; Steven, S.; et al. Environmental noise induces the release of stress hormones and inflammatory signaling molecules leading to oxidative stress and vascular dysfunction-signatures of the internal exposome. Biofactors 2019, 45, 495–506. [Google Scholar] [CrossRef]
- Munzel, T.; Sorensen, M.; Daiber, A. Transportation noise pollution and cardiovascular disease. Nat. Rev. Cardiol 2021. [Google Scholar] [CrossRef] [PubMed]
- Karbach, S.; Wenzel, P.; Waisman, A.; Munzel, T.; Daiber, A. Enos uncoupling in cardiovascular diseases--the role of oxidative stress and inflammation. Curr Pharm Des. 2014, 20, 3579–3594. [Google Scholar] [CrossRef] [PubMed]
- Di Cesare, A.; Di Meglio, P.; Nestle, F.O. The il-23/th17 axis in the immunopathogenesis of psoriasis. J. Investig. Dermatol. 2009, 129, 1339–1350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardi, C.; Matheson, R.; Zachariae, C.; Cameron, G.; Li, L.; Edson-Heredia, E.; Braun, D.; Banerjee, S. Anti-interleukin-17 monoclonal antibody ixekizumab in chronic plaque psoriasis. N. Engl. J. Med. 2012, 366, 1190–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, K.A.; Leonardi, C.; Menter, A.; Ortonne, J.P.; Krueger, J.G.; Kricorian, G.; Aras, G.; Li, J.; Russell, C.B.; Thompson, E.H.; et al. Brodalumab, an anti-interleukin-17-receptor antibody for psoriasis. N. Engl. J. Med. 2012, 366, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Crispin, J.C.; Tsokos, G.C. Il-17 in systemic lupus erythematosus. J. Biomed. Biotechnol. 2010, 2010, 943254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choy, E. Understanding the dynamics: Pathways involved in the pathogenesis of rheumatoid arthritis. Rheumatology 2012, 51 (Suppl. 5), v3–v11. [Google Scholar] [CrossRef] [Green Version]
- Pasceri, V.; Yeh, E.T. A tale of two diseases: Atherosclerosis and rheumatoid arthritis. Circulation 1999, 100, 2124–2126. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.; Kossmann, S.; Munzel, T.; Daiber, A. Redox regulation of cardiovascular inflammation - immunomodulatory function of mitochondrial and nox-derived reactive oxygen and nitrogen species. Free Radic Biol. Med. 2017, 109, 48–60. [Google Scholar] [CrossRef]
- Wenzel, P.; Knorr, M.; Kossmann, S.; Stratmann, J.; Hausding, M.; Schuhmacher, S.; Karbach, S.H.; Schwenk, M.; Yogev, N.; Schulz, E.; et al. Lysozyme m-positive monocytes mediate angiotensin ii-induced arterial hypertension and vascular dysfunction. Circulation 2011, 124, 1370–1381. [Google Scholar] [CrossRef] [Green Version]
- Joles, J.A.; Koomans, H.A. Causes and consequences of increased sympathetic activity in renal disease. Hypertension 2004, 43, 699–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hybertson, B.M.; Gao, B.; Bose, S.K.; McCord, J.M. Oxidative stress in health and disease: The therapeutic potential of nrf2 activation. Mol. Aspects Med. 2011, 32, 234–246. [Google Scholar] [CrossRef]
- Li, J.; Ichikawa, T.; Janicki, J.S.; Cui, T. Targeting the nrf2 pathway against cardiovascular disease. Expert Opin. Ther. Targets 2009, 13, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Liby, K.T.; Sporn, M.B. Synthetic oleanane triterpenoids: Multifunctional drugs with a broad range of applications for prevention and treatment of chronic disease. Pharmacol. Rev. 2012, 64, 972–1003. [Google Scholar] [CrossRef] [Green Version]
- Schaue, D.; McBride, W.H. Opportunities and challenges of radiotherapy for treating cancer. Nat. Rev. Clin. Oncol. 2015, 12, 527–540. [Google Scholar] [CrossRef]
- Brune, B.; Dehne, N.; Grossmann, N.; Jung, M.; Namgaladze, D.; Schmid, T.; von Knethen, A.; Weigert, A. Redox control of inflammation in macrophages. Antioxids Redox Signal. 2013, 19, 595–637. [Google Scholar] [CrossRef] [Green Version]
- Aminzadeh, M.A.; Reisman, S.A.; Vaziri, N.D.; Shelkovnikov, S.; Farzaneh, S.H.; Khazaeli, M.; Meyer, C.J. The synthetic triterpenoid rta dh404 (cddo-dhtfea) restores endothelial function impaired by reduced nrf2 activity in chronic kidney disease. Redox Biol. 2013, 1, 527–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Rizky, L.; Stefanovic, N.; Tate, M.; Ritchie, R.H.; Ward, K.W.; de Haan, J.B. The nuclear factor (erythroid-derived 2)-like 2 (nrf2) activator dh404 protects against diabetes-induced endothelial dysfunction. Cardiovasc. Diabetol. 2017, 16, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.M.; Sharma, A.; Stefanovic, N.; Yuen, D.Y.; Karagiannis, T.C.; Meyer, C.; Ward, K.W.; Cooper, M.E.; de Haan, J.B. Derivative of bardoxolone methyl, dh404, in an inverse dose-dependent manner lessens diabetes-associated atherosclerosis and improves diabetic kidney disease. Diabetes 2014, 63, 3091–3103. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.E.; Timme-Laragy, A.R.; Karchner, S.I.; Stegeman, J.J. Nrf2 and nrf2-related proteins in development and developmental toxicity: Insights from studies in zebrafish (danio rerio). Free Radic Biol. Med. 2015, 88, 275–289. [Google Scholar] [CrossRef] [Green Version]
- Meissner, M.; Doll, M.; Hrgovic, I.; Reichenbach, G.; Konig, V.; Hailemariam-Jahn, T.; Gille, J.; Kaufmann, R. Suppression of vegfr2 expression in human endothelial cells by dimethylfumarate treatment: Evidence for anti-angiogenic action. J. Invest. Dermatol. 2011, 131, 1356–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, N.G.; Kappas, A. Pharmacological and clinical aspects of heme oxygenase. Pharmacol. Rev. 2008, 60, 79–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balla, J.; Vercellotti, G.M.; Jeney, V.; Yachie, A.; Varga, Z.; Eaton, J.W.; Balla, G. Heme, heme oxygenase and ferritin in vascular endothelial cell injury. Mol. Nutr. Food Res. 2005, 49, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.; Yamamoto, Y.; McDonagh, A.F.; Glazer, A.N.; Ames, B.N. Bilirubin is an antioxidant of possible physiological importance. Science 1987, 235, 1043–1046. [Google Scholar] [CrossRef]
- Hopkins, P.N.; Wu, L.L.; Hunt, S.C.; James, B.C.; Vincent, G.M.; Williams, R.R. Higher serum bilirubin is associated with decreased risk for early familial coronary artery disease. Arterioscler. Thromb. Vasc. Biol. 1996, 16, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.Y.; Takeshige, K.; Cheung, B.S.; Minakami, S. Bilirubin inhibits the activation of superoxide-producing nadph oxidase in a neutrophil cell-free system. Biochim. Biophys. Acta 1991, 1076, 369–373. [Google Scholar] [CrossRef]
- Amit, Y.; Boneh, A. Bilirubin inhibits protein kinase c activity and protein kinase c-mediated phosphorylation of endogenous substrates in human skin fibroblasts. Clin. Chim. Acta 1993, 223, 103–111. [Google Scholar] [CrossRef]
- Nitti, M.; Furfaro, A.L.; Mann, G.E. Heme oxygenase dependent bilirubin generation in vascular cells: A role in preventing endothelial dysfunction in local tissue microenvironment? Front. Physiol. 2020, 11, 23. [Google Scholar] [CrossRef]
- Kruger, A.L.; Peterson, S.J.; Schwartzman, M.L.; Fusco, H.; McClung, J.A.; Weiss, M.; Shenouda, S.; Goodman, A.I.; Goligorsky, M.S.; Kappas, A.; et al. Upregulation of heme oxygenase provides vascular protection in an animal model of diabetes through its anti-oxidant and anti-apoptotic effects. J. Pharmacol. Exp. Ther. 2006, 319, 1144–1152. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, P.; Oelze, M.; Coldewey, M.; Hortmann, M.; Seeling, A.; Hink, U.; Mollnau, H.; Stalleicken, D.; Weiner, H.; Lehmann, J.; et al. Heme oxygenase-1: A novel key player in the development of tolerance in response to organic nitrates. Arterioscler Thromb. Vasc. Biol. 2007, 27, 1729–1735. [Google Scholar] [CrossRef]
- Detsika, M.G.; Duann, P.; Atsaves, V.; Papalois, A.; Lianos, E.A. Heme oxygenase 1 up-regulates glomerular decay accelerating factor expression and minimizes complement deposition and injury. Am. J. Pathol. 2016, 186, 2833–2845. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, F.; Curtis, L.M.; Truong, L.; Poss, K.; Visner, G.A.; Madsen, K.; Nick, H.S.; Agarwal, A. Heme oxygenase-1 gene ablation or expression modulates cisplatin-induced renal tubular apoptosis. Am. J. Physiol. Renal. Physiol. 2000, 278, F726–F736. [Google Scholar] [CrossRef] [Green Version]
- Scuderi, S.A.; Ardizzone, A.; Paterniti, I.; Esposito, E.; Campolo, M. Antioxidant and anti-inflammatory effect of nrf2 inducer dimethyl fumarate in neurodegenerative diseases. Antioxidants 2020, 9, 630. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado, A.; Manda, G.; Hassan, A.; Alcaraz, M.J.; Barbas, C.; Daiber, A.; Ghezzi, P.; Leon, R.; Lopez, M.G.; Oliva, B.; et al. Transcription factor nrf2 as a therapeutic target for chronic diseases: A systems medicine approach. Pharmacol. Rev. 2018, 70, 348–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempen, E.V.; Casas, M.; Pershagen, G.; Foraster, M. Who environmental noise guidelines for the european region: A systematic review on environmental noise and cardiovascular and metabolic effects: A summary. Int. J. Environ. Res. Public Health 2018, 15, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munzel, T.; Schmidt, F.P.; Steven, S.; Herzog, J.; Daiber, A.; Sorensen, M. Environmental noise and the cardiovascular system. J. Am. Coll. Cardiol. 2018, 71, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Munzel, T.; Kroller-Schon, S.; Oelze, M.; Gori, T.; Schmidt, F.P.; Steven, S.; Hahad, O.; Roosli, M.; Wunderli, J.M.; Daiber, A.; et al. Adverse cardiovascular effects of traffic noise with a focus on nighttime noise and the new who noise guidelines. Annu. Rev. Public Health 2020, 41, 309–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bayo Jimenez, M.T.; Frenis, K.; Kröller-Schön, S.; Kuntic, M.; Stamm, P.; Kvandová, M.; Oelze, M.; Li, H.; Steven, S.; Münzel, T.; et al. Noise-Induced Vascular Dysfunction, Oxidative Stress, and Inflammation Are Improved by Pharmacological Modulation of the NRF2/HO-1 Axis. Antioxidants 2021, 10, 625. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040625
Bayo Jimenez MT, Frenis K, Kröller-Schön S, Kuntic M, Stamm P, Kvandová M, Oelze M, Li H, Steven S, Münzel T, et al. Noise-Induced Vascular Dysfunction, Oxidative Stress, and Inflammation Are Improved by Pharmacological Modulation of the NRF2/HO-1 Axis. Antioxidants. 2021; 10(4):625. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040625
Chicago/Turabian StyleBayo Jimenez, Maria Teresa, Katie Frenis, Swenja Kröller-Schön, Marin Kuntic, Paul Stamm, Miroslava Kvandová, Matthias Oelze, Huige Li, Sebastian Steven, Thomas Münzel, and et al. 2021. "Noise-Induced Vascular Dysfunction, Oxidative Stress, and Inflammation Are Improved by Pharmacological Modulation of the NRF2/HO-1 Axis" Antioxidants 10, no. 4: 625. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040625