
New Insights on Hemp Oil Enriched in Cannabidiol: Decarboxylation, Antioxidant Properties and In Vitro Anticancer Effect

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. C. sativa L. Plant Growth
2.2. Hemp Oil Extraction
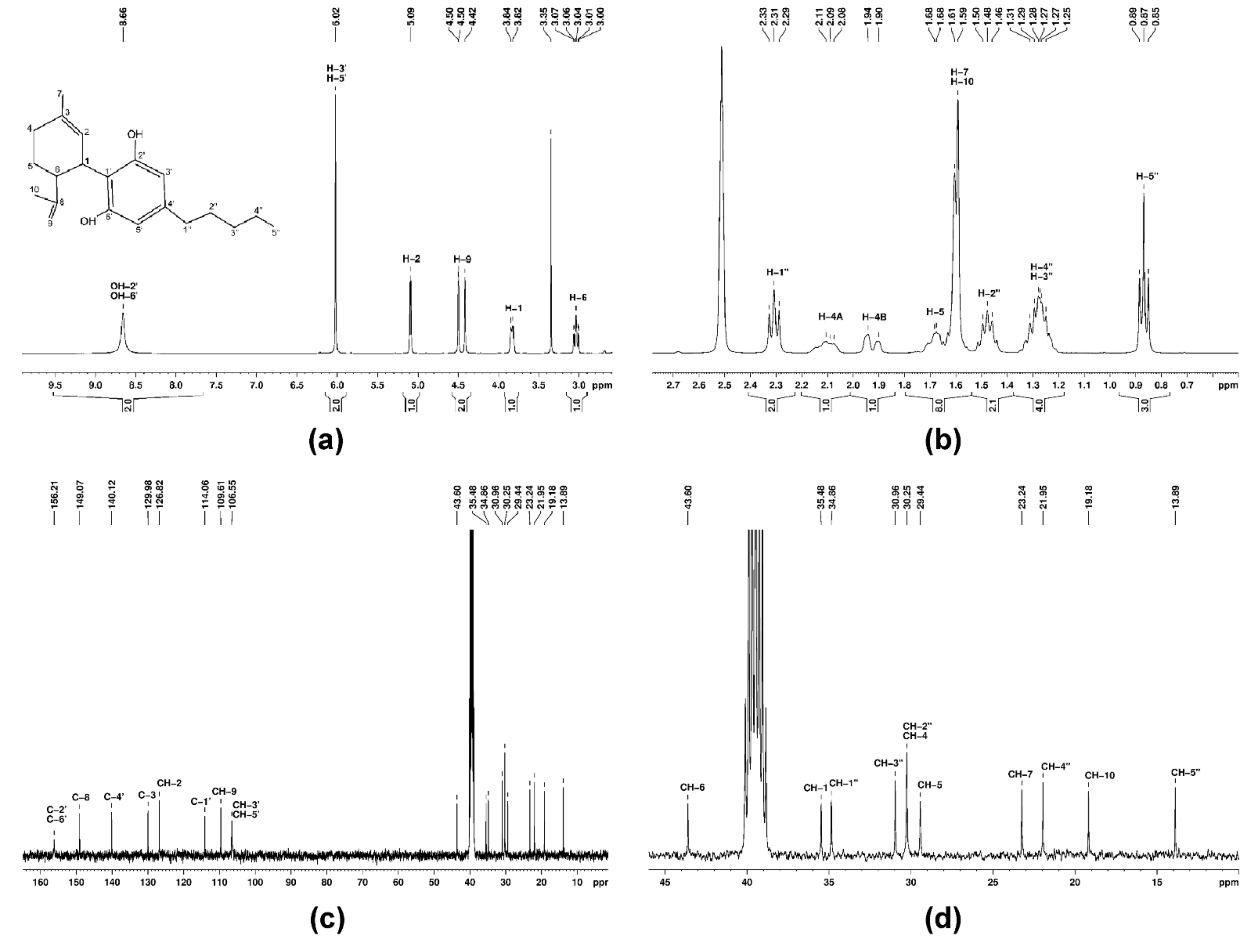
2.3. CBD Purification and Characterization
2.4. Decarboxylation of CBDA from Hemp Oil
2.5. In Vitro Antioxidant Activity Evaluation
2.5.1. Ferrous Ions’ (Fe2+) Chelating Activity
2.5.2. Ferric Ions (Fe3+) Reducing Antioxidant Power (FRAP) Assay
2.5.3. Superoxide Anion Radical (O2●−) Scavenging Activity
2.5.4. Hydroxyl Radical (HO●) Scavenging Ability
2.5.5. Lipid Peroxidation Inhibitory Assay
2.6. Cell Culture
2.7. Cytotoxicity Assay (MTS)
2.8. Morphological Analysis
2.9. Acridine Orange/Ethidium Bromide (AO/EB) Staining for Apoptosis
2.10. Evaluation of Intracellular H2O2 Production
2.11. RNA Isolation and Gene Expression Analysis
2.12. Statistical Analysis
3. Results and Discussion
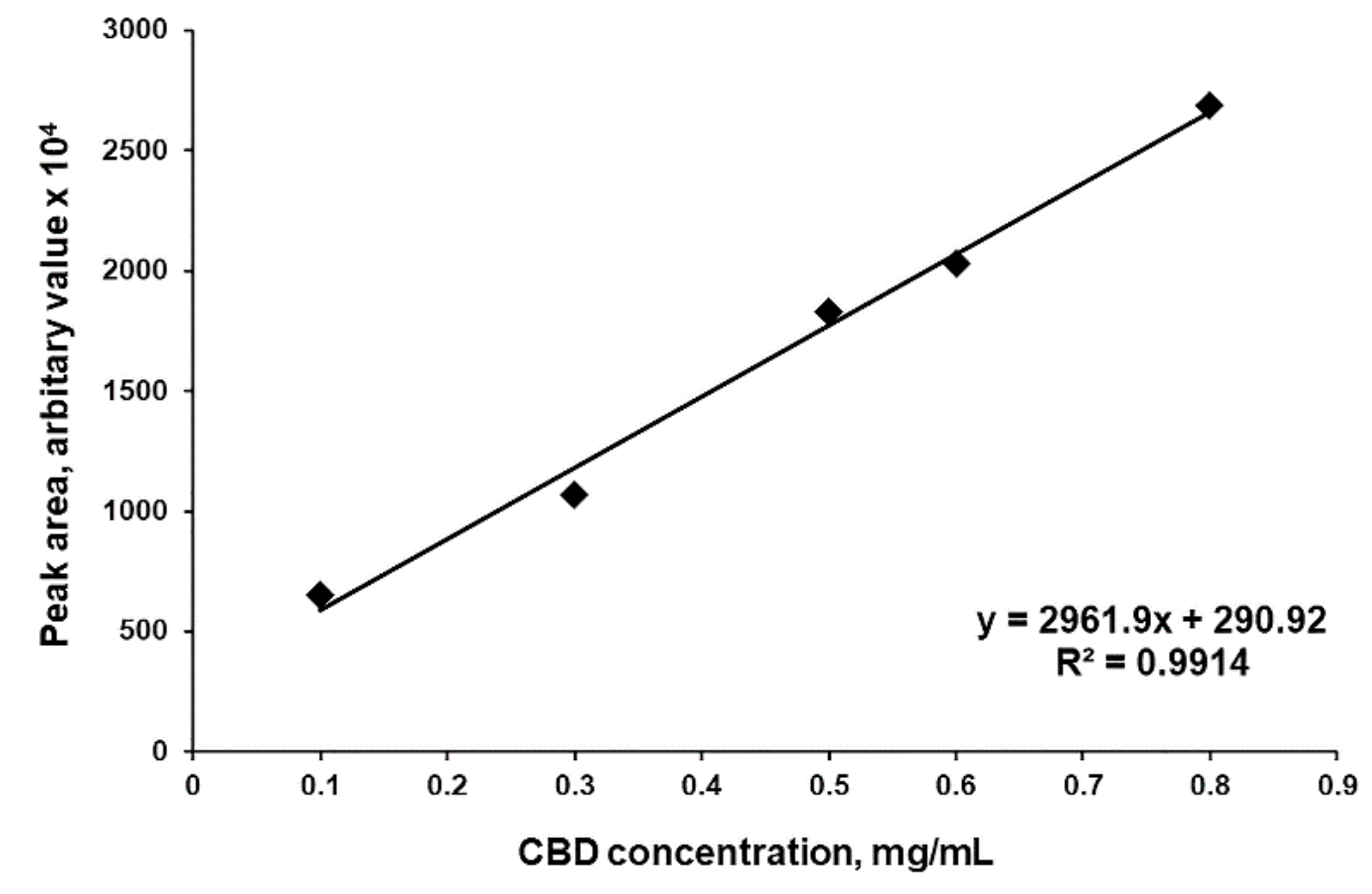
3.1. Hemp Oil Extraction, Purification and Characterization
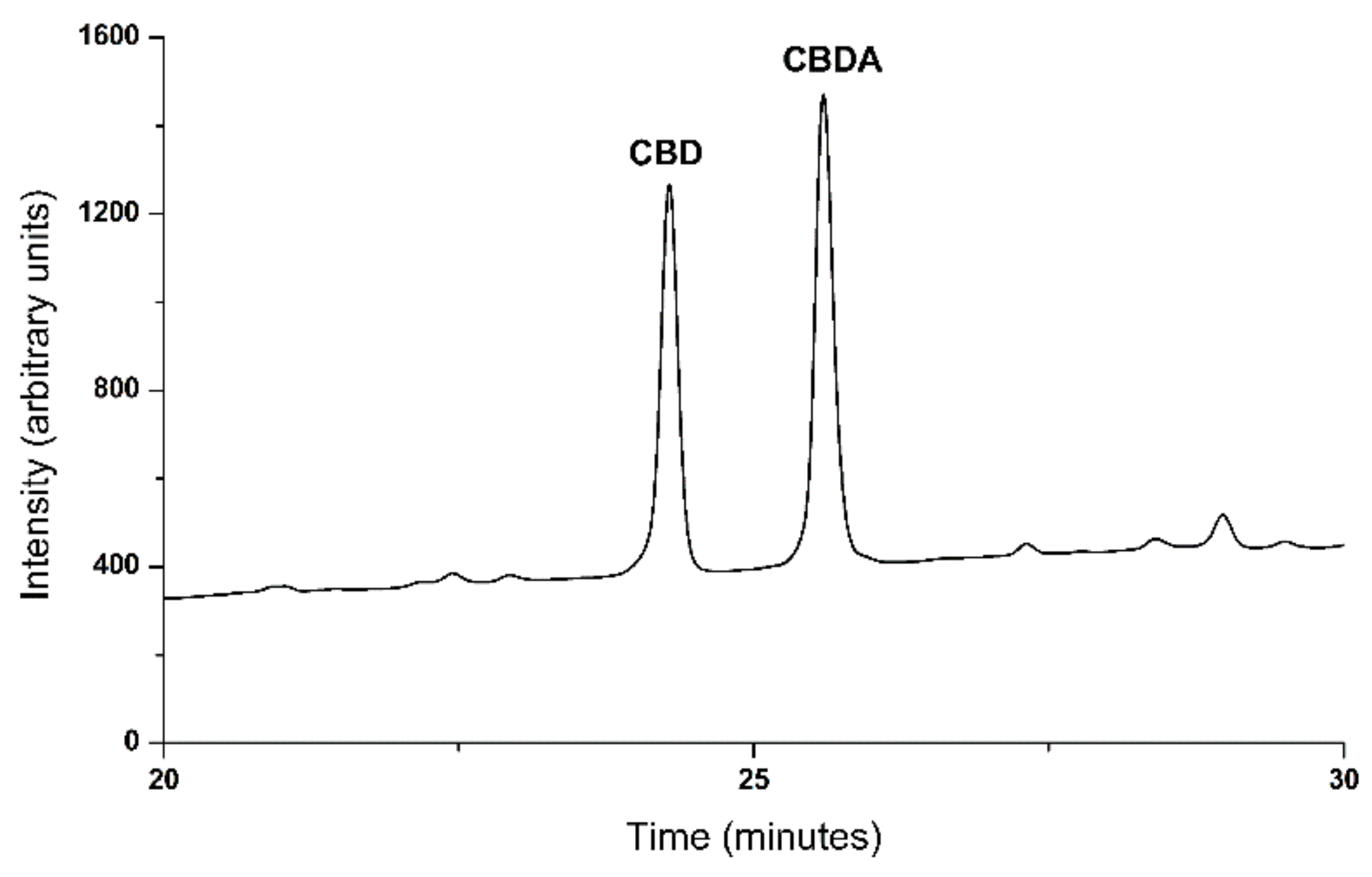
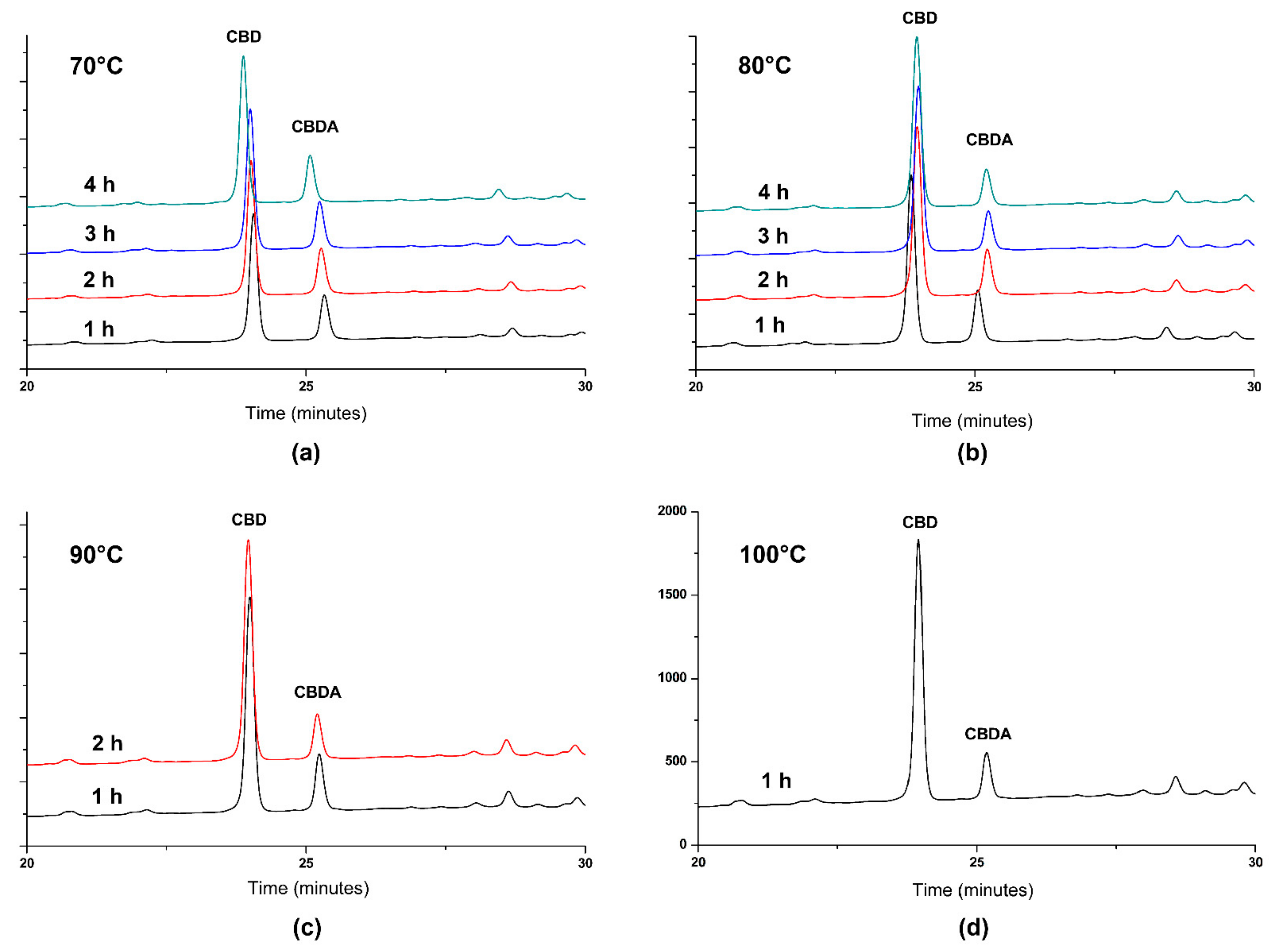
3.2. Decarboxylation of CBDA from Hemp Oil
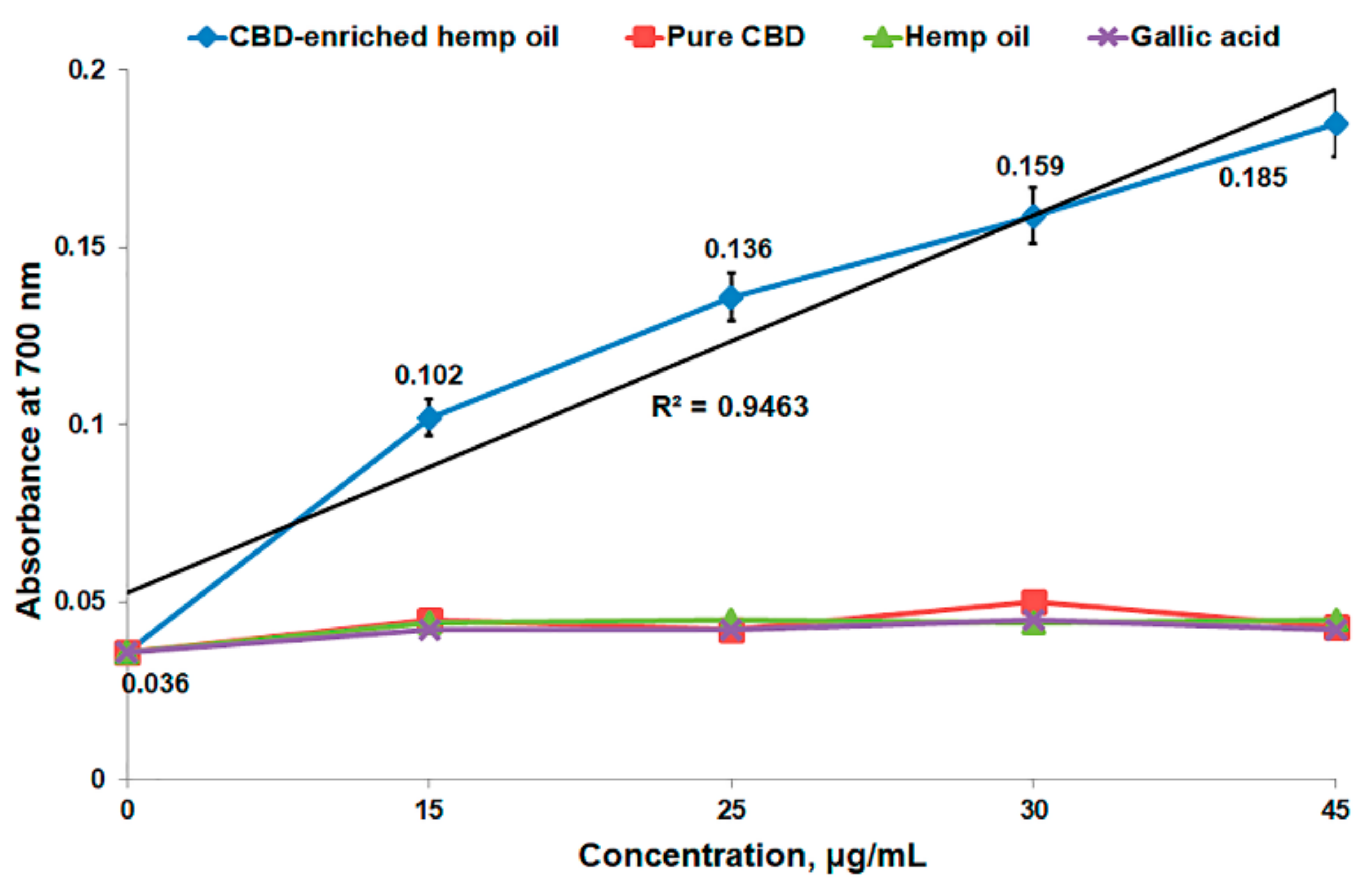
3.3. In Vitro Antioxidant Activity of CBD-Enriched Hemp Oil
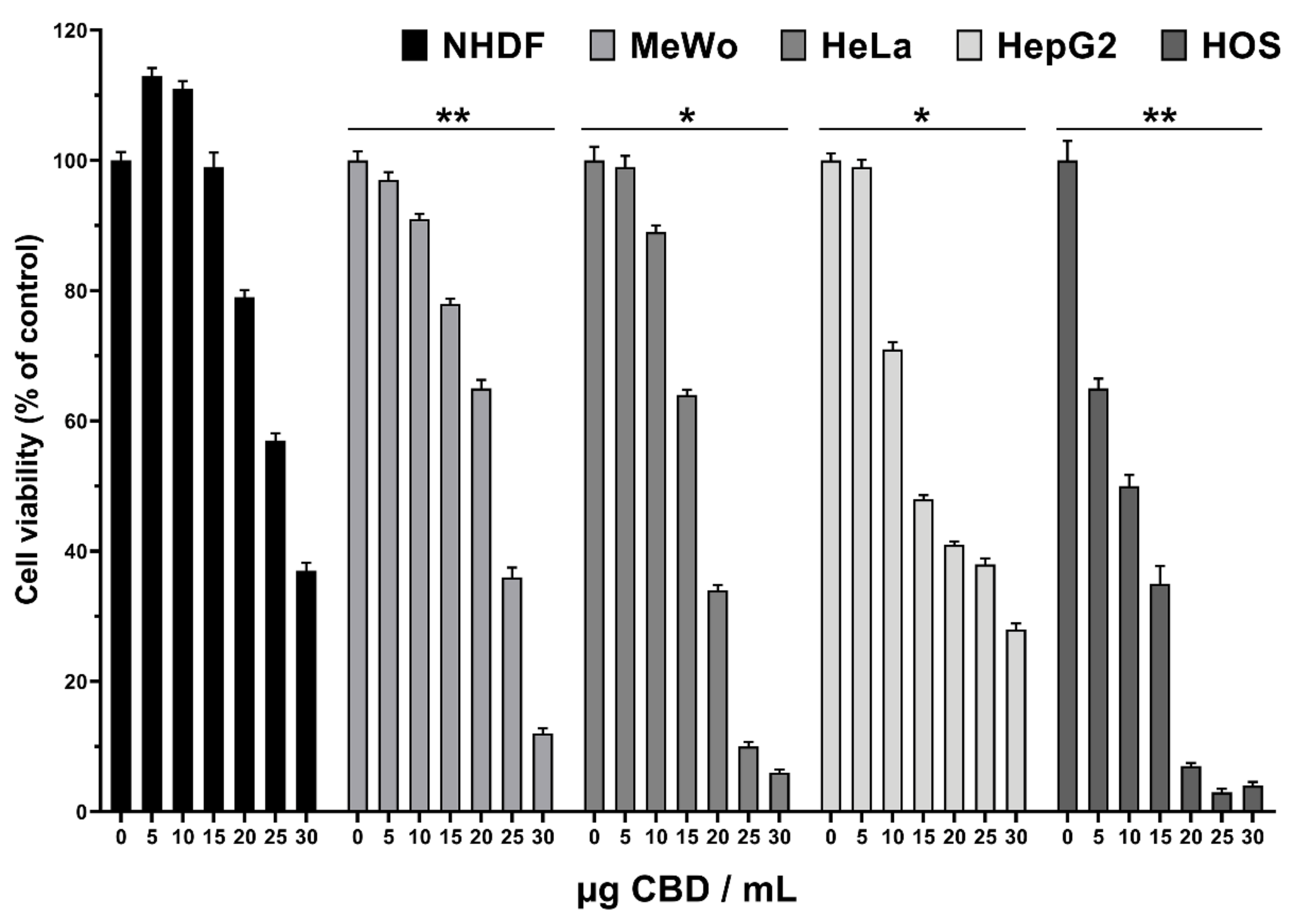
3.4. Cytotoxicity of CBD-Enriched Hemp Oil
3.5. Evaluation of CBD-Enriched Hemp Oil-Induced Apoptosis
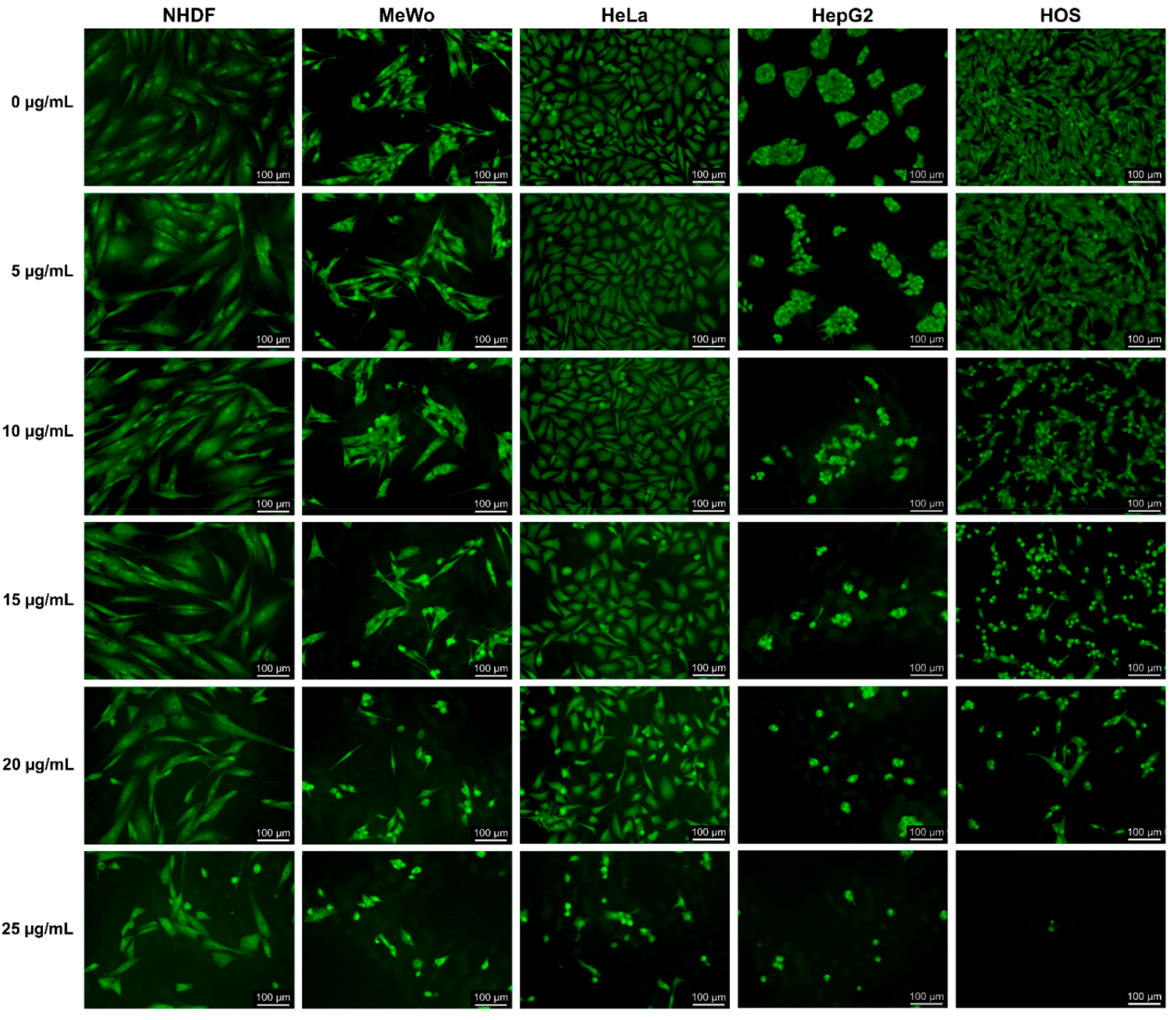
3.5.1. Morphological Analysis
3.5.2. Acridine Orange/Ethidium Bromide (AO/EB) Staining for Apoptosis
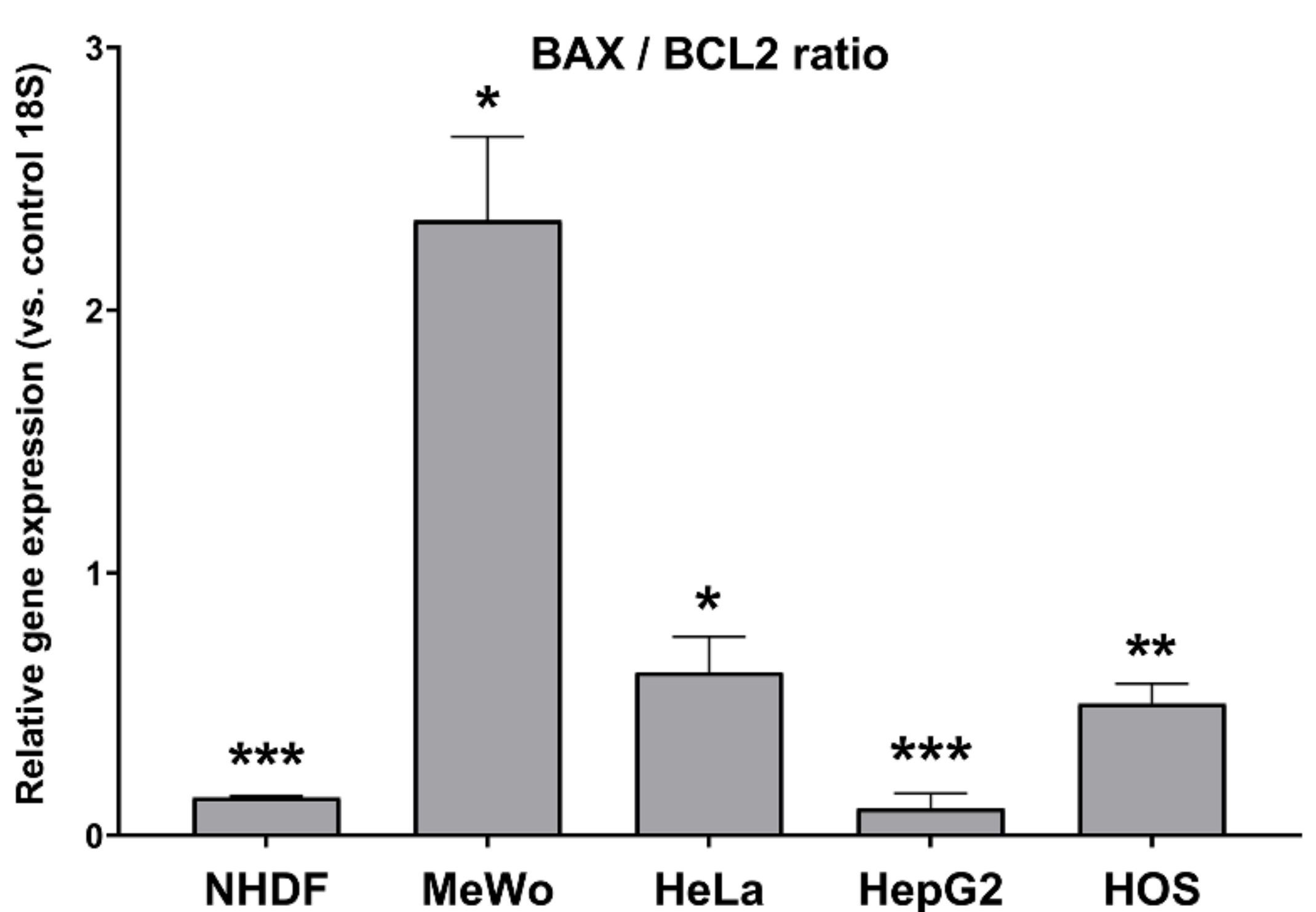
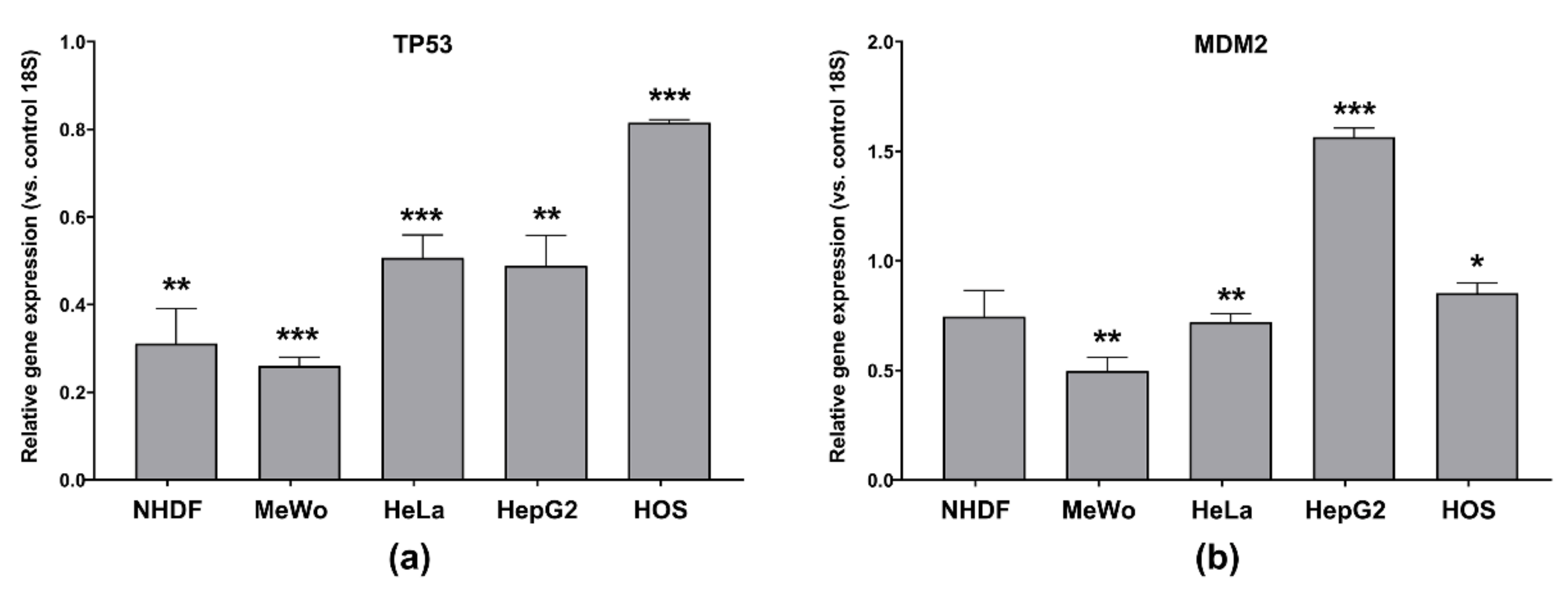
3.5.3. Apoptosis-Related Genes’ Expression
3.6. Assessment of Oxidative Stress Induced by CBD-Enriched Hemp Oil
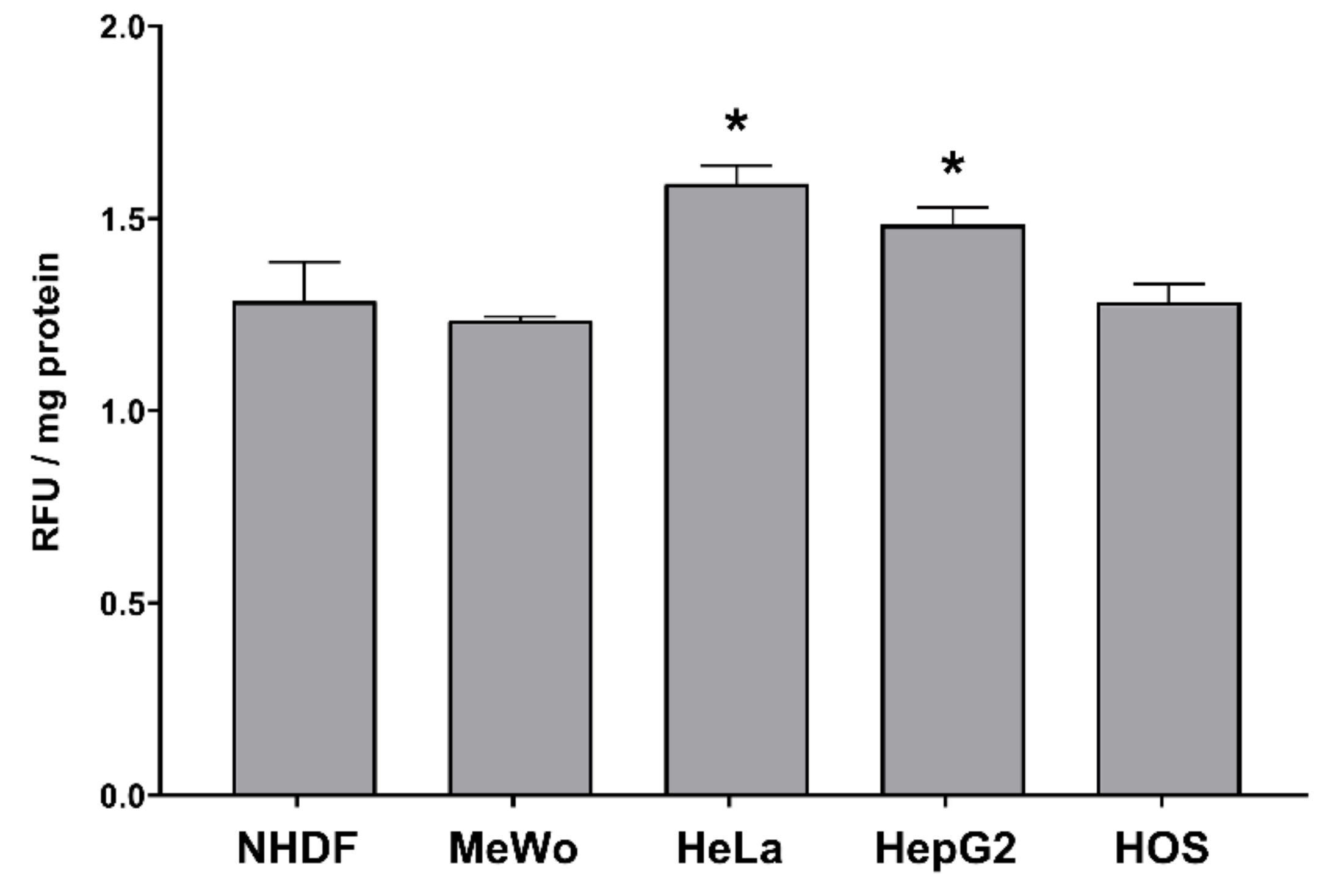
3.6.1. Intracellular H2O2 Production
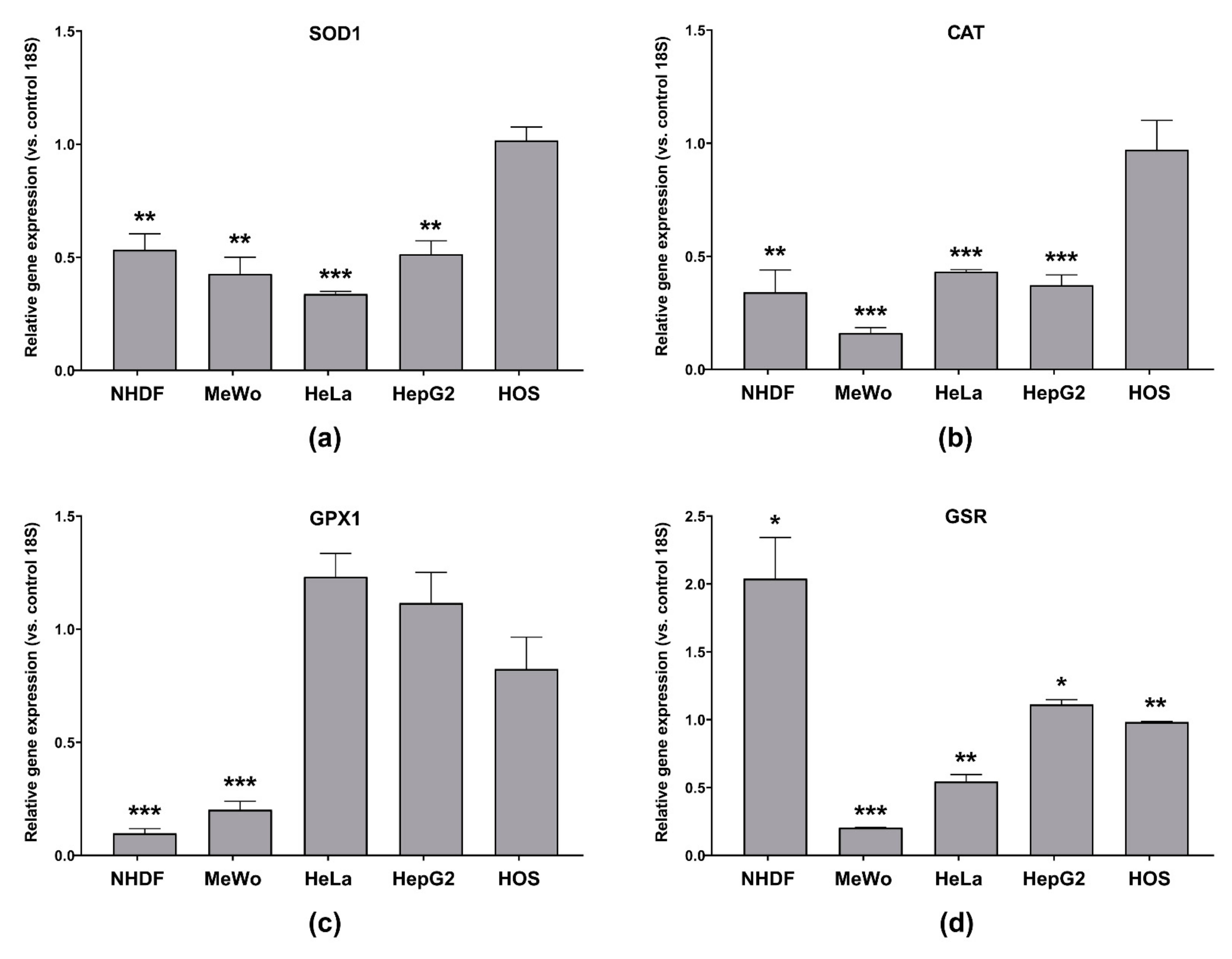
3.6.2. Antioxidant Enzymes’ Gene Expression
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fiorini, D.; Molle, A.; Nabissi, M.; Santini, G.; Benelli, G.; Maggi, F. Valorizing industrial hemp (Cannabis sativa L.) by-products: Cannabidiol enrichment in the inflorescence essential oil optimizing sample pre-treatment prior to distillation. Ind. Crops Prod. 2019, 128, 581–589. [Google Scholar] [CrossRef]
- Di Giacomo, V.; Recinella, L.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Politi, M.; Antolini, M.D.; Acquaviva, A.; et al. Metabolomic profile and antioxidant/anti-inflammatory effects of industrial hemp water extract in fibroblasts, keratinocytes and isolated mouse skin specimens. Antioxidants 2021, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Adorisio, S.; Delfino, D.; Chiavaroli, A.; Brunetti, L.; Recinella, L.; Leone, S.; D’antonio, M.; Zengin, G.; Acquaviva, A.; et al. Comparative investigation of composition, antifungal, and anti-inflammatory effects of the essential oil from three industrial hemp varieties from Italian cultivation. Antibiotics 2021, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- Nagy, D.U.; Cianfaglione, K.; Maggi, F.; Sut, S.; Dall’Acqua, S. Chemical Characterization of Leaves, Male and Female Flowers from Spontaneous Cannabis (Cannabis sativa L.) Growing in Hungary. Chem. Biodivers. 2019, 16. [Google Scholar] [CrossRef]
- Tomko, A.M.; Whynot, E.G.; Ellis, L.D.; Dupré, D.J. Anti-cancer potential of cannabinoids, terpenes, and flavonoids present in cannabis. Cancers 2020, 12, 1985. [Google Scholar] [CrossRef]
- Śledziński, P.; Zeyland, J.; Słomski, R.; Nowak, A. The current state and future perspectives of cannabinoids in cancer biology. Cancer Med. 2018, 7, 765–775. [Google Scholar] [CrossRef]
- Gülçin, I. Fe3+–Fe2+ transformation method: An important antioxidant assay. Methods Mol. Biol. 2015, 1208, 233–246. [Google Scholar] [CrossRef]
- Lennicke, C.; Rahn, J.; Lichtenfels, R.; Wessjohann, L.A.; Seliger, B. Hydrogen peroxide—Production, fate and role in redox signaling of tumor cells. Cell Commun. Signal. 2015, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Atalay, S.; Jarocka-karpowicz, I.; Skrzydlewska, E.; Skrzydlewskas, E. Antioxidative and anti-inflammatory properties of cannabidiol. Antioxidants 2020, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Citti, C.; Pacchetti, B.; Vandelli, M.A.; Forni, F.; Cannazza, G. Analysis of cannabinoids in commercial hemp seed oil and decarboxylation kinetics studies of cannabidiolic acid (CBDA). J. Pharm. Biomed. Anal. 2018, 149, 532–540. [Google Scholar] [CrossRef]
- Ocque, A.J.; Hagler, C.E.; DiFrancesco, R.; Lombardo, J.; Morse, G.D. Development and validation of an assay to measure cannabidiol and Δ9-tetrahydrocannabinol in human EDTA plasma by UHPLC-MS/MS. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1112, 56–60. [Google Scholar] [CrossRef]
- Escrivá, Ú.; Andrés-Costa, M.J.; Andreu, V.; Picó, Y. Analysis of cannabinoids by liquid chromatography–mass spectrometry in milk, liver and hemp seed to ensure food safety. Food Chem. 2017, 228, 177–185. [Google Scholar] [CrossRef]
- Gülçin, I.; Elmastaş, M.; Aboul-Enein, H.Y. Antioxidant activity of clove oil—A powerful antioxidant source. Arab. J. Chem. 2012, 5, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, X.; Chen, D.; Chen, S. Antioxidant activity and mechanism of protocatechuic acid in Vitro. Funct. Foods Heal. Dis. 2011, 1, 232–244. [Google Scholar] [CrossRef]
- Xiong, L.; Ni, X.; Niu, L.; Zhou, Y.; Wang, Q.; Khalique, A.; Liu, Q.; Zeng, Y.; Shu, G.; Pan, K.; et al. Isolation and Preliminary Screening of a Weissella confusa Strain from Giant Panda (Ailuropoda melanoleuca). Probiotics Antimicrob. Proteins 2019, 11, 535–544. [Google Scholar] [CrossRef]
- Ahmed, D.; Khan, M.M.; Saeed, R. Comparative analysis of phenolics, flavonoids, and antioxidant and antibacterial potential of methanolic, hexanic and aqueous extracts from Adiantum caudatum leaves. Antioxidants 2015, 4, 394–409. [Google Scholar] [CrossRef]
- Badisa, R.B.; Darling-Reed, S.F.; Joseph, P.; Cooperwood, J.S.; Latinwo, L.M.; Goodman, C.B. Selective cytotoxic activities of two novel synthetic drugs on human breast carcinoma MCF-7 cells. Anticancer Res. 2009, 29, 2993–2996. [Google Scholar]
- Ribble, D.; Goldstein, N.B.; Norris, D.A.; Shellman, Y.G. A simple technique for quantifying apoptosis in 96-well plates. BMC Biotechnol. 2005. [Google Scholar] [CrossRef] [Green Version]
- Toma, L.; Sanda, G.M.; Deleanu, M.; Stancu, C.S.; Sima, A.V. Glycated LDL increase VCAM-1 expression and secretion in endothelial cells and promote monocyte adhesion through mechanisms involving endoplasmic reticulum stress. Mol. Cell. Biochem. 2016, 417, 169–179. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008. [Google Scholar] [CrossRef]
- Fathordoobady, F.; Singh, A.; Kitts, D.D.; Pratap Singh, A. Hemp (Cannabis Sativa L.) Extract: Anti-Microbial Properties, Methods of Extraction, and Potential Oral Delivery. Food Rev. Int. 2019, 35, 664–684. [Google Scholar] [CrossRef]
- Choi, Y.H.; Hazekamp, A.; Peltenburg-Looman, A.M.G.; Frédérich, M.; Erkelens, C.; Lefeber, A.W.M.; Verpoorte, R. NMR assignments of the major cannabinoids and cannabiflavonoids isolated from flowers of Cannabis sativa. Phytochem. Anal. 2004, 15, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Petrović, M.; Debeljak, Ž.; Kezić, N.; Džidara, P. Relationship between cannabinoids content and composition of fatty acids in hempseed oils. Food Chem. 2015, 170, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Borges, R.S.; Da Silva, A.B.F. Cannabidiol as an Antioxidant. In Handbook of Cannabis and Related Pathologies: Biology, Pharmacology, Diagnosis, and Treatment; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. e122–e130. ISBN 9780128008270. [Google Scholar]
- Suktham, T.; Jones, A.; Soliven, A.; Dennis, G.R.; Shalliker, R.A. A comparison of the performance of the cupric reducing antioxidant potential assay and the ferric reducing antioxidant power assay for the analysis of antioxidants using reaction flow chromatography. Microchem. J. 2019, 149, 104046. [Google Scholar] [CrossRef]
- Hacke, A.C.M.; Lima, D.; De Costa, F.; Deshmukh, K.; Li, N.; Chow, A.M.; Marques, J.A.; Pereira, R.P.; Kerman, K. Probing the antioxidant activity of Δ9-tetrahydrocannabinol and cannabidiol in Cannabis sativa extracts. Analyst 2019, 144, 4952–4961. [Google Scholar] [CrossRef]
- Moccia, S.; Siano, F.; Russo, G.L.; Volpe, M.G.; La Cara, F.; Pacifico, S.; Piccolella, S.; Picariello, G. Antiproliferative and antioxidant effect of polar hemp extracts (Cannabis sativa L., Fedora cv.) in human colorectal cell lines. Int. J. Food Sci. Nutr. 2020, 71, 410–423. [Google Scholar] [CrossRef]
- Moldzio, R.; Pacher, T.; Krewenka, C.; Kranner, B.; Novak, J.; Duvigneau, J.C.; Rausch, W.D. Effects of cannabinoids Δ(9)-tetrahydrocannabinol, Δ(9)-tetrahydrocannabinolic acid and cannabidiol in MPP+ affected murine mesencephalic cultures. Phytomedicine 2012, 19, 819–824. [Google Scholar] [CrossRef]
- Choi, W.H.; Park, H.D.; Baek, S.H.; Chu, J.P.; Kang, M.H.; Mi, Y.J. Cannabidiol induces cytotoxicity and cell death via apoptotic pathway in cancer cell lines. Biomol. Ther. 2008, 16, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Lukhele, S.T.; Motadi, L.R. Cannabidiol rather than Cannabis sativa extracts inhibit cell growth and induce apoptosis in cervical cancer cells. BMC Complement. Altern. Med. 2016, 16, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Manosroi, A.; Chankhampan, C.; Kietthanakorn, B.O.; Ruksiriwanich, W.; Chaikul, P.; Boonpisuttinant, K.; Sainakham, M.; Manosroi, W.; Tangjai, T.; Manosroi, J. Pharmaceutical and cosmeceutical biological activities of hemp (cannabis sativa L var. sativa) leaf and seed extracts. Chiang Mai J. Sci. 2019, 46, 180–195. [Google Scholar]
- Blázquez, C.; Carracedo, A.; Barrado, L.; José Real, P.; Luis Fernández-Luna, J.; Velasco, G.; Malumbres, M.; Guzmán, M.; Blázquez, C.; Carracedo, A.; et al. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006, 20, 2633–2635. [Google Scholar] [CrossRef] [Green Version]
- Notaro, A.; Sabella, S.; Pellerito, O.; Di Fiore, R.; De Blasio, A.; Vento, R.; Calvaruso, G.; Giuliano, M. Involvement of PAR-4 in cannabinoid-dependent sensitization of osteosarcoma cells to TRAIL-induced Apoptosis. Int. J. Biol. Sci. 2014, 10, 466–478. [Google Scholar] [CrossRef] [Green Version]
- Niu, F.; Zhao, S.; Xu, C.Y.; Sha, H.; Bi, G.B.; Chen, L.; Ye, L.; Gong, P.; Nie, T.H. Potentiation of the antitumor activity of adriamycin against osteosarcoma by cannabinoid WIN-55,212-2. Oncol. Lett. 2015, 10, 2415–2421. [Google Scholar] [CrossRef] [Green Version]
- Kosgodage, U.S.; Mould, R.; Henley, A.B.; Nunn, A.V.; Guy, G.W.; Thomas, E.L.; Inal, J.M.; Bell, J.D.; Lange, S. Cannabidiol (CBD) is a novel inhibitor for exosome and microvesicle (EMV) release in cancer. Front. Pharmacol. 2018, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Simmerman, E.; Qin, X.; Yu, J.C.; Baban, B. Cannabinoids as a Potential New and Novel Treatment for Melanoma: A Pilot Study in a Murine Model. J. Surg. Res. 2019, 235, 210–215. [Google Scholar] [CrossRef]
- Kasibhatla, S. Acridine Orange/Ethidium Bromide (AO/EB) Staining to Detect Apoptosis. Cold Spring Harb. Protoc. 2006, 2006, pdb-prot4493. [Google Scholar] [CrossRef]
- Kulsoom, B.; Shamsi, T.S.; Afsar, N.A.; Memon, Z.; Ahmed, N.; Hasnain, S.N. Bax, Bcl-2, and Bax/Bcl-2 as prognostic markers in acute myeloid leukemia: Are we ready for bcl-2-directed therapy? Cancer Manag. Res. 2018, 10, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Raisova, M.; Hossini, A.M.; Eberle, J.; Riebeling, C.; Wieder, T.; Sturm, I.; Daniel, P.T.; Orfanos, C.E.; Geilen, C.C. The Bax/Bcl-2 ratio determines the susceptibility of human melanoma cells to CD95/Fas-mediated apoptosis. J. Investig. Dermatol. 2001, 117, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Khodapasand, E.; Jafarzadeh, N.; Farrokhi, F.; Kamalidehghan, B.; Houshmand, M. Is Bax/Bcl-2 ratio considered as a prognostic marker with age and tumor location in colorectal cancer? Iran. Biomed. J. 2015, 19, 69–75. [Google Scholar] [CrossRef]
- Nag, S.; Qin, J.; Srivenugopal, K.S.; Wang, M.; Zhang, R. The MDM2-p53 pathway revisited. J. Biomed. Res. 2013, 27, 254–271. [Google Scholar] [CrossRef]
- Alharris, E.; Singh, N.P.; Nagarkatti, P.S.; Nagarkatti, M. Role of miRNA in the regulation of cannabidiol-mediated apoptosis in neuroblastoma cells. Oncotarget 2019, 10, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massi, P.; Vaccani, A.; Bianchessi, S.; Costa, B.; Macchi, P.; Parolaro, D. The non-psychoactive cannabidiol triggers caspase activation and oxidative stress in human glioma cells. Cell. Mol. Life Sci. 2006, 63, 2057–2066. [Google Scholar] [CrossRef] [PubMed]
- Matés, J.M.; Pérez-Gómez, C.; De Castro, I.N. Antioxidant enzymes and human diseases. Clin. Biochem. 1999, 32, 595–603. [Google Scholar] [CrossRef]
- Usami, N.; Yamamoto, I.; Watanabe, K. Generation of reactive oxygen species during mouse hepatic microsomal metabolism of cannabidiol and cannabidiol hydroxy-quinone. Life Sci. 2008, 83, 717–724. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | Sequence 5′-3′ | Amplicon Size (bp) |
|---|---|---|---|
| 18S | NR_145820.1 | FW 5′ GGAGCCTGCGGCTTAATTTG RV 5′ CCACCCACGGAATCGAGAAA | 100 |
| BAX | NM_138761.3 | FW 5′ TCATGGGCTGGACATTGGAC RV 5′ GCGTCCCAAAGTAGGAGAGG | 96 |
| BCL2 | NM_000633.2 | FW 5′ GCGGCCTCTGTTTGATTTCTC RV 5′ CTTGTGGCCCAGATAGGCA | 100 |
| TP53 | NM_001276696.2 | FW 5′ CAGCACATGACGGAGGTTGT RV 5′ TCATCCAAATACTCCACACGC | 125 |
| MDM2 | NM_002392.5 | FW 5′ CAGTAGCAGTGAATCTACAGGGA RV 5′ CTGATCCAACCAATCACCTGAAT | 85 |
| SOD1 | NM_000454.4 | FW 5′ GGAAGTCGTTTGGCTTGTGG RV 5′ GGGCCTCAGACTACATCCAAG | 70 |
| CAT | NM_001752.4 | FW 5′ CTGACTACGGGAGCCACATC RV 5′ AGATCCGGACTGCACAAAGG | 92 |
| GPX1 | NM_000581.4 | FW 5′ CAGTCGGTGTATGCCTTCTCG RV 5′ GAGGGACGCCACATTCTCG | 105 |
| GSR | NM_000637.5 | FW 5′ GCACTTGCGTGAATGTTGGA RV 5′ TCACATAGGCATCCCGCTTT | 156 |
| Nr. Crt. | Decarboxylation Conditions 1 | CBD in 50 mg Hemp Oil (mg) | CBDA in 50 mg Hemp Oil (mg) | CBD + CBDA in 50 mg Hemp Oil (mg) | CBDin Hemp Oil (%) 2 | CBDA in Hemp Oil (%) 2 | CBD Transformation from CBDA (%) 2 | CBDA + CBD Yield after Decarboxylation Process (%) 2 | CBDA + CBD Degradation Yield (%) 2 |
|---|---|---|---|---|---|---|---|---|---|
| Crude oil | 4.57 | 6.77 | 11.345 | 9.14 | 13.54 | - | - | - | |
| 1 | 70 °C, 1 h | 5.47 | 1.95 | 7.43 | 10.94 | 3.9 | 71.19 | 65.4 | 34.6 |
| 2 | 70 °C, 2 h | 5.7 | 1.89 | 7.59 | 11.4 | 3.78 | 72.08 | 66.9 | 33.1 |
| 3 | 70 °C, 3 h | 6.03 | 2.1 | 8.13 | 12.06 | 4.2 | 68.98 | 71.66 | 28.34 |
| 4 | 70 °C, 4 h | 6.17 | 2.05 | 8.22 | 12.34 | 4.1 | 69.72 | 72.45 | 27.55 |
| 5 | 80 °C, 1 h | 7.47 | 2.38 | 9.85 | 14.94 | 4.76 | 64.84 | 86.82 | 13.18 |
| 6 | 80 °C, 2 h | 7.65 | 2.16 | 9.81 | 15.3 | 4.32 | 68.09 | 86.47 | 13.57 |
| 7 | 80 °C, 3 h | 7.69 | 1.81 | 9.5 | 15.38 | 3.62 | 73.26 | 83.73 | 16.27 |
| 8 | 80 °C, 4 h | 7.83 | 1.7 | 9.53 | 15.66 | 3.4 | 74.89 | 84.0 | 16.0 |
| 9 | 90 °C, 1 h | 8.6 | 2.26 | 10.86 | 17.2 | 4.52 | 66.62 | 95.72 | 4.28 |
| 10 | 90 °C, 2 h | 8.91 | 1.8 | 10.81 | 17.82 | 3.8 | 73.41 | 95.28 | 4.72 |
| 11 | 100 °C, 1 h | 7.9 | 1.35 | 9.25 | 15.8 | 2.7 | 80.06 | 81.53 | 18.47 |
| Samples/Method | Fe2+ Chelating Activity (%) | FRAP Assay (%) 1 | O2●− Scavenging Activity (%) | HO● Scavenging Ability (%) | Lipid Peroxidation Inhibitory Assay (%) 2 |
|---|---|---|---|---|---|
| CBD-enriched hemp oil (15 μg CBD/mL) | 27.26 ± 0.2 | 55.0 ± 1.3 | 69.1 ± 3 | 221.5 ± 2.56 | 59.77 ± 2.0 |
| Pure CBD (15 μg/mL) | 3.7 ± 0.1 | 104.6 ± 2.1 | 0.41 ± 0.01 | 167 ± 2.15 | 1.66 ± 0.04 |
| Crude hemp oil (15 μg CBD/mL) | 8.0 ± 0.03 | 97.7 ± 1.7 | 1.24 ± 0.03 | 20.69 ± 0.63 | 22.36 ± 1.18 |
| Gallic acid (15 μg/mL) | 3.8 ± 0.07 | 95.45 ± 1.3 | 1.23 ± 0.01 | 391.38 ± 5.67 | 21.33 ± 1.03 |
| NHDF | MeWo | HeLa | HepG2 | HOS | |
|---|---|---|---|---|---|
| IC50 (µg CBD/mL) | 26.65 | 21.59 | 16.89 | 16.85 | 8.42 |
| SI | - | 1.23 | 1.58 | 1.58 | 3.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrovici, A.R.; Simionescu, N.; Sandu, A.I.; Paraschiv, V.; Silion, M.; Pinteala, M. New Insights on Hemp Oil Enriched in Cannabidiol: Decarboxylation, Antioxidant Properties and In Vitro Anticancer Effect. Antioxidants 2021, 10, 738. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050738
Petrovici AR, Simionescu N, Sandu AI, Paraschiv V, Silion M, Pinteala M. New Insights on Hemp Oil Enriched in Cannabidiol: Decarboxylation, Antioxidant Properties and In Vitro Anticancer Effect. Antioxidants. 2021; 10(5):738. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050738
Chicago/Turabian StylePetrovici, Anca Roxana, Natalia Simionescu, Andreea Isabela Sandu, Vasile Paraschiv, Mihaela Silion, and Mariana Pinteala. 2021. "New Insights on Hemp Oil Enriched in Cannabidiol: Decarboxylation, Antioxidant Properties and In Vitro Anticancer Effect" Antioxidants 10, no. 5: 738. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10050738